











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkLos camélidos sudamericanos fueron el recurso animal más explotado por las poblaciones humanas a lo largo de toda el área andina, principalmente, durante el Holoceno tardío, y han tenido un papel central en su subsistencia (Bonavia 2009; Le Neün et al. 2023; Mengoni Goñalons 2007; Olivera y Grant 2008; Wheeler, Pires-Ferreira y Kaulicke 1976; Yacobaccio 2003). Por esto, su identificación taxonómica, a partir de los restos óseos recuperados de los contextos arqueológicos, es central para los estudios zooarqueológicos del área que buscan comprender y explicar las estrategias económicas empleadas por los grupos humanos.
En el NOA se encuentran representadas únicamente tres de las cuatro especies que conforman la familia Camelidae: dos especies silvestres, la vicuña (Vicugna vicugna) y el guanaco (Lama guanicoe), y una doméstica, la llama (Lama glama). Esto se debe a que en esta porción del área andina no están dadas las condiciones ecológicas necesarias para que puedan desarrollarse prácticas de cría de alpaca (Vicugna pacos), especie que requiere ambientes húmedos de altura (Yacobaccio 2001).
En términos esqueletales, las especies de camélidos sudamericanos tienen gran semejanza morfológica (Izeta 2006; Menegaz, Salemme y Ortiz 1988). Los denominados camélidos pequeños (vicuñas y alpacas) cuentan con tamaños corporales similares. Lo mismo sucede con los camélidos grandes (guanacos y llamas), cuyas dimensiones corporales son parecidas, siendo la llama de mayor tamaño que el guanaco norteño. Por lo tanto, el empleo de la morfometría tradicional (MT) (sensuMarcus 1990) u osteometría para su identificación taxonómica, a partir de los restos óseos recuperados del registro arqueológico, presenta limitaciones debido a los solapamientos de tamaño existentes entre los taxa silvestres y domésticos que constituyen cada grupo de tamaño (Cartajena 2009; L’Heureux 2010; Mengoni Goñalons y Yacobaccio 2006).
Los estudios zooarqueológicos en el NOA no enfrentan demasiadas dificultades para identificar a la vicuña por medio de sus restos esqueletales debido a la ausencia, en la región, de la especie doméstica de porte corporal similar (i. e. la alpaca). Por el contrario, los guanacos y las llamas coexisten en el registro de distintos sitios del área a partir de los ca. 4100 AP (Yacobaccio 2003, 2001 ). Por ende, estimar sus frecuencias de representación es fundamental para caracterizar las estrategias de caza y pastoreo desarrolladas por las poblaciones humanas prehispánicas.
Los estudios de morfometría geométrica (MG) brindan la posibilidad de estudiar de manera cuantitativa la forma de una estructura, independientemente del tamaño, conservando la información sobre su geometría mediante los análisis (Adams, Rohlf y Slice 2004; Zeldich et al. 2004). Han sido empleados en diversas investigaciones zooarqueológicas, entre otros temas, para identificar especies encontradas en el registro, diferenciar taxa silvestres y domésticos y estudiar procesos de domesticación (Balasse et al. 2016; Evin et al. 2013; Haruda 2017; Owen et al. 2014; Seetah et al. 2014, entre otros). Su aplicación al estudio de los camélidos sudamericanos es reciente, aunque ha generado información biológica y morfológica valiosa y variada acerca de estos ungulados (Balcarcel et al. 2021; Conte e Izeta 2019; Hernández 2019, 2016 ; Hernández y L’Heureux 2019; Hernández, L’Heureux y Leoni 2021; L’Heureux y Hernández 2016, 2015 ; L’Heureux, Hernández y Kaufmann 2023; L’Heureux y Kaufmann 2013, 2012 ; López y Gómez 2016).
Teniendo en consideración lo mencionado, en este trabajo nos proponemos diferenciar las primeras falanges pertenecientes a las especies de camélidos de mayor porte mediante el estudio de la forma de dichos elementos. Para ello se expone la revisión de una muestra de especímenes óseos correspondientes a Lama sp. procedentes del sitio Quebrada del Cementerio (Jujuy, Argentina), datado en torno a los 1065-1404 años DC (Nielsen 2001; Nielsen et al. 2003-2005 ), con técnicas de MG en dos dimensiones (2D). Los resultados de este estudio, que fue realizado entre 2019 y 2024, son integrados y comparados con los obtenidos en investigaciones zooarqueológicas previas, a partir del análisis del tamaño de las falanges con técnicas de MT (Mercolli 2005, 2004 ). Se espera que estos nuevos estudios permitan precisar la composición taxonómica del grupo de camélidos de mayor tamaño, compuesto por dos especies de dimensiones corporales similares y con baja variabilidad morfológica interespecífica (L’Heureux 2010; Yacobaccio et al. 1997-1998 ), para discutir más adelante las características y el aporte de la caza de guanacos y del pastoreo de llamas a la subsistencia de las poblaciones humanas que habitaron el tramo medio de la Quebrada de Humahuaca entre los 900-600 años AP.
Quebrada del Cementerio: ubicación y antecedentes
Quebrada del Cementerio se localiza en el tramo medio de la Quebrada de Humahuaca (Jujuy, Argentina) a una altitud de 2560 m s. n. m. (Nielsen et al. 2003-2005 ). El sitio se halla próximo a la localidad actual de Maimará (departamento de Tilcara) y a 1,5 km de distancia hacia el oeste del río Grande, principal curso de agua que recorre la quebrada troncal (Nielsen et al. 2003-2005 ) (figura 1).
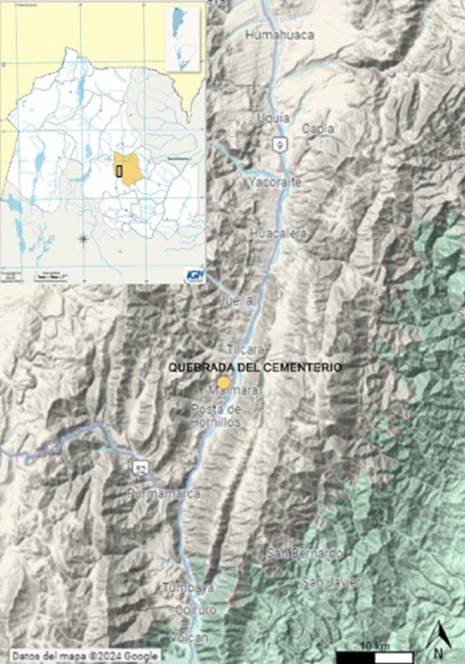
Fuente: zona de Quebrada de Humahuaca, Jujuy (Argentina) con localización del sitio arqueológico Quebrada del Cementerio. 2024. Anahí Hernández y Pablo Mercolli, Google Maps, https://www.google.com/maps/@-23.5392078,-65.1628891,10z/data=!5m1!1e4?hl=es&entry=ttu&g_ep=EgoyMDI0MTExMS4wIKXMDSoASAFQAw%3D%3D (22 de octubre de 2024).
Figura 1. Localización geográfica del sitio Quebrada del Cementerio.
Los trabajos arqueológicos se llevaron a cabo en 1997, en el marco de una escuela de campo organizada por el Instituto Interdisciplinario Tilcara. Estos se concentraron en dos sectores: el sector 1 que se corresponde con una estructura situada en el límite oriental del sitio (recinto 1) y, el sector 2, que comprende una terraza localizada en el espacio central (conjunto conformado por varios recintos y área de descarte previa) (Nielsen et al. 2003-2005 ). A partir de estas investigaciones, el sitio fue caracterizado como un asentamiento de ocupación breve (sensuNielsen y Rivolta 1997), con áreas de actividad doméstica y ausencia de estructuras destinadas a las prácticas agrícolas (Mercolli 2010, 2004 ). Los fechados radiocarbónicos obtenidos arrojaron antigüedades de 835 ± 40 AP y 630 ± 35 AP, y ubican la ocupación del sitio entre los 1065-1404 años DC (Nielsen 2001; Nielsen et al. 2003-2005 ).
El conjunto arqueofaunístico recuperado fue abundante (NISP 4737) y su revisión estuvo a cargo de uno de los autores de esta investigación (Mercolli 2010, 2004). Entre los taxa representados, los camélidos resultaron ser preponderantes en la muestra (50 %) (Mercolli 2010, 2004 ). Los estudios osteométricos llevados a cabo sobre las falanges y los metapodios de estos artiodáctilos indicaron un amplio predominio de especímenes de tamaño similar al de la llama actual (63,2 %), seguidos de aquellos de tamaño pequeño (30,5 %) que pertenecerían a vicuñas y, por último, los de tamaño intermedio (6,3 %) que se asimilan a guanacos andinos o a llamas pequeñas (Mercolli 2010, 2004 ). En relación con el perfil etario, los estados de fusión ósea sugieren una baja representación de crías, un grupo intermedio de animales juveniles y subadultos y un porcentaje menor de adultos (Mercolli 2010, 2004 ). En cambio, los análisis de mandíbulas y maxilares muestran una mayor proporción de camélidos adultos de 4, 9 y más de 10 años (Mercolli 2011).
Materiales y métodos
Este artículo se basó en el estudio, con técnicas de MG 2D, de 41 primeras falanges completas de camélidos modernos de gran porte (n = 19 guanacos y n = 22 llamas) procedentes de distintas localidades geográficas, que hacen parte de colecciones personales y de distintas instituciones nacionales1 (Hernández, L’Heureux y Leoni 2021), y de 47 especímenes arqueológicos recuperados del sitio Quebrada del Cementerio. Dicho estudio se desarrolló entre 2019 (relevamiento fotográfico de especímenes arqueológicos y de algunos especímenes actuales) y 2024 (realización de análisis estadísticos en el periodo 2022-2024). Las primeras falanges arqueológicas fueron discriminadas en delanteras (n = 23) y posteriores (n = 24), siguiendo los criterios definidos por Kent (1982). La elección de dicho elemento poscraneal se debe a que suelen ser abundantes y presentar buenos niveles de preservación en contextos arqueológicos y, en función de la información morfológica proporcionada, han sido ampliamente utilizadas en los estudios de MT (Cartajena 2009; Izeta, Otaola y Gasco 2009, Yacobaccio 2010), incluyendo el caso particular de Quebrada del Cementerio (Mercolli 2010, 2004 ).
Entre los guanacos modernos se hallan representados individuos pertenecientes a las dos subespecies reconocidas: el guanaco norteño (Lama guanicoe cacsilensis) y el guanaco patagónico (Lama guanicoe guanicoe). Entre ambas subespecies existen diferencias significativas en su tamaño corporal, siendo el guanaco patagónico más grande que el norteño (Franklin 1982; Lönnberg 1913; Mengoni Goñalons y Yacobaccio 2006; Raedecke 1979). Por lo tanto, no suelen utilizarse muestras provenientes de zonas meridionales a los 35° de latitud sur en los análisis de MT u osteometría de muestras procedentes del NOA. Su inclusión en este trabajo obedece a que no se revisa el tamaño de las falanges, sino exclusivamente las variables de forma de dicho elemento. Asimismo, análisis previos de elementos poscraneales de guanacos con técnicas de MG 2D no han mostrado, hasta el momento, variaciones de forma ligadas a un patrón geográfico clinal (Hernández 2019; L’Heureux y Hernández 2016) como sí sucede con los estudios de tamaño (Gasco y Cardillo 2014; Kent 1982; L’Heureux 2008; L’Heureux y Cornaglia 2015; Mengoni Goñalons y Yacobaccio 2006).
Las imágenes bidimensionales de las primeras falanges delanteras y traseras derechas completas (vista dorsal) de guanacos y llamas de más de 36 meses de edad fueron analizadas mediante configuraciones de landmarks y semilandmarks. Se definieron un total de 10 landmarks y 4 semilandmarks ( L’Heureux y Hernández 2016) (figura 2). Para reducir el efecto de la localización inicial arbitraria de los semilandmarks, estos fueron deslizados siguiendo el criterio de Bending Energy (Adams, Rohlf y Slice 2013). Además, se llevó a cabo una Análisis Generalizado de Procrustes para ajustar las coordenadas de puntos y eliminar toda la información no relacionada con la forma (Zeldich et al. 2004).
Se evaluó la existencia de alometría en la muestra mediante una regresión multivariante entre las variables dependientes de forma (coordenadas de Procrustes) y las variables independientes de tamaño (centroid size) (Klingenberg 2011). De esta manera, se estimó si este último incidía en las variaciones de forma para remover el efecto alométrico en caso de existir. En este sentido, si la regresión indicara alometría, las variables de forma empleadas ulteriormente para los análisis serían los residuos de la regresión, ya que solo proporcionan datos de forma (Klingenberg 2010). En cambio, si la regresión no mostrara efecto alométrico, se emplearían las coordenadas de Procrustes.
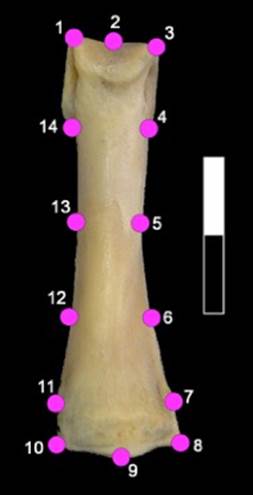
Nota: landmarks: 1. Extremo latero-distal (dirección craneal) de la superficie articular distal; 2. Punto más profundo del surco de la epífisis distal; 3. Extremo disto-medial de la superficie articular distal; 4. Unión medial de la epífisis distal y la diáfisis; 7. Ancho máximo de la epífisis proximal (medial); 8. Extremo medial de la superficie articular proximal; 9. Punto más proximal (dirección caudal) de la superficie dorsal de la epífisis proximal; 10. Extremo lateral de la superficie articular proximal; 11. Ancho máximo de la epífisis proximal (lateral); 14. Unión lateral de la epífisis distal y la diáfisis.
Fuente: fotografía de los autores, Buenos Aires, Argentina, 2024.
Figura 2. Configuración de landmarks y semilandmarks definidos sobre las primeras falanges de camélidos.
Más adelante, las variables de forma fueron sometidas a dos pruebas estadísticas multivariadas: análisis de componentes principales (PCA) y análisis discriminante (DA). El PCA se utilizó con el objetivo de conocer la máxima variación morfológica existente en las primeras falanges de guanacos y llamas modernas, y estudiar la distribución de los especímenes arqueológicos dentro de dicho espacio de variación. Junto a los PCA se presentan las variaciones morfológicas extremas versus la forma consenso (Bookstein 1996). Por su parte, el DA permitió analizar llamas y guanacos modernos para estimar el porcentaje de especímenes correctamente clasificados con validación cruzada, y clasificar a los especímenes arqueológicos con el grupo de llamas o de guanacos modernos mediante la función predictiva del test. Para ello se seleccionó la cantidad de variables de Procrustes o componentes principales (PC) que mostraron el mejor ajuste de la función predictiva (Kovarovic et al. 2011).
Los análisis fueron llevados a cabo por un único observador. Previamente, se evaluó la incidencia del error intraobservador en la digitalización de landmarks y semilandmarks con una prueba de Anova de medidas repetidas (Zar 1999) y el coeficiente de correlación intraclase (CCI; Yezerinac, Lougheed y Handford 1992). Los resultados indicaron buenos niveles de acuerdo entre las series de datos revisadas y una elevada consistencia en la obtención y replicabilidad de los datos (Hernández 2019).
Los estudios se ejecutaron mediante el uso de los softwares tpsUtil 1.82, tpsDig 2.31, tpsRelw 1.70, MorphoJ 1.07a y Past 3.24 (Hammer, Harper y Ryan 2001; Klingenberg 2011; Rohlf 2022, 2019, 2017 ).
Resultados
Las regresiones de forma-tamaño para las primeras falanges delanteras y posteriores arrojaron resultados dispares. Así, para las primeras falanges delanteras se identificó la existencia de alometría (p = 0,00), de modo que el 7,52 % de las variaciones de forma registradas son explicadas por cambios en su tamaño (figura 3A). Por el contrario, no se registró efecto alométrico para las primeras falanges posteriores (p = 0,5) (figura 3B). En este caso, solo el 1,5 % de la variación de la forma de las falanges se explica por cambios en su tamaño.
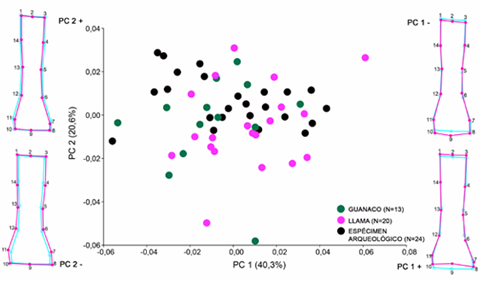
El PCA para las primeras falanges delanteras muestra una amplia superposición en la forma de las falanges pertenecientes a guanacos y llamas modernas, siguiendo los dos ejes principales de variación (figura 4). Los especímenes arqueológicos de Quebrada del Cementerio se agrupan tanto con llamas como con guanacos modernos, aunque en general se distribuyen en mayor medida hacia los valores positivos del PC 1. Los tres primeros PC explican el 70,3 % de la variación morfológica registrada para este elemento.
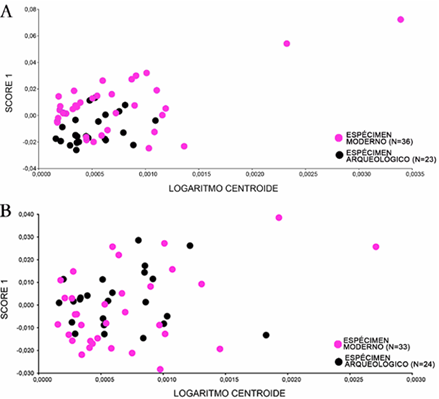
Fuente: los autores, Buenos Aires, 2024.
Figura 3A. Regresión forma-tamaño para las primeras falanges delanteras; 3B. Regresión forma-tamaño para las primeras falanges posteriores.

Nota: variaciones extremas (rosa) versus forma consenso (celeste) para cada extremo de los dos ejes principales.
Fuente: los autores, Buenos Aires, 2024.
Figura 4. PCA para las primeras falanges delanteras de guanacos y llamas modernas y especímenes arqueológicos.
Con respecto a la morfología de las primeras falanges delanteras, se observa que, hacia el extremo positivo del eje principal de variación, las epífisis proximales son ligeramente menos anchas y tienen el extremo dorsal-caudal más pronunciado que la forma consenso, además del punto más profundo del surco de la epífisis distal ligeramente más marcado (figura 4). Hacia el extremo negativo se registra la tendencia opuesta. Al seguir el segundo eje de variación, los cambios se concentran en la epífisis proximal. Hacia el extremo positivo, las falanges son ligeramente menos anchas y con el extremo dorsal-caudal menos pronunciado que la forma consenso, lo que registra la tendencia contraria hacia el extremo negativo del eje.
El PCA correspondiente a las primeras falanges posteriores indica que la forma de las falanges de los guanacos y las llamas modernas ocupan el mismo espacio de variación morfológica a lo largo de los dos ejes principales, y no se registra ningún ordenamiento de los casos (figura 5). Los especímenes arqueológicos de Quebrada del Cementerio se agrupan tanto con llamas como con guanacos modernos, aunque siguiendo el eje de las Y se distribuyen en mayor medida hacia los valores positivos. Los tres primeros PC explican el 72,7 % de la variación morfológica registrada para este elemento.
Nota: variaciones extremas (rosa) versus forma consenso (celeste) para cada extremo de los dos ejes principales.
Fuente: los autores, Buenos Aires, 2024.
Figura 5. PCA para las primeras falanges posteriores de guanacos y llamas modernas y especímenes arqueológicos.
Si tenemos en cuenta las variaciones extremas versus la forma consenso se advierte que, hacia el extremo positivo del principal eje de variación, las falanges muestran el extremo dorsal-caudal menos pronunciado que la forma consenso y el punto más profundo del surco de la epífisis distal menos marcado (figura 5). Hacia el extremo negativo se registra la tendencia opuesta. Al seguir el segundo eje de variación, los cambios se observan en las diáfisis, de modo que hacia el extremo positivo las falanges son menos anchas que la forma consenso y, hacia el extremo negativo del eje, más anchas.
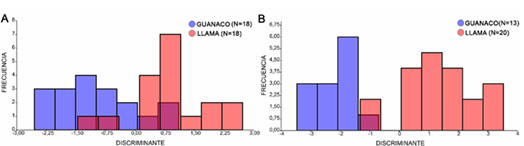
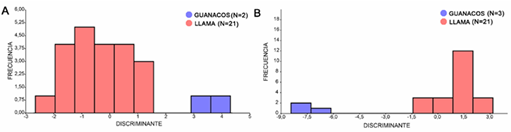
Por otra parte, el DA para las primeras falanges delanteras se realizó tomando a los PC como variables de forma (figura 6A). El mejor ajuste de la función discriminante se logró a partir de la utilización de los primeros 11 PC. En este sentido, el 72,2 % de los especímenes modernos fue clasificado correctamente (tabla 1). Del total de especímenes arqueológicos analizados, solo 2 mostraron características morfológicas similares a los guanacos modernos, mientras que los 21 restantes fueron clasificados junto a las llamas modernas (figura 7A).
Fuente: los autores, Buenos Aires, 2024.
Figura 6A. DA para las primeras falanges delanteras de guanacos y llamas actuales. 6B. DA para las primeras falanges posteriores de guanacos y llamas actuales.
Tabla 1. Tabla con la clasificación de las primeras falanges delanteras y posteriores de guanacos y llamas modernas según función discriminante (validación cruzada)
| Primeras falanges delanteras | Primeras falanges posteriores | ||||||
|---|---|---|---|---|---|---|---|
| Guanaco | Llama | Total | Llama | Guanaco | Total | ||
| Guanaco | 13 | 5 | 18 | Llama | 16 | 4 | 20 |
| Llama | 5 | 13 | 18 | Guanaco | 0 | 13 | 13 |
| Total | 18 | 18 | 36 | Total | 16 | 17 | 33 |
Fuente: los autores, Buenos Aires, 2024.
El DA para las primeras falanges posteriores se llevó a cabo a partir del uso de 12 coordenadas de Procrustes, ya que mostraron el mejor ajuste de la función discriminante (figura 6B). Así, el 87,9 % de los especímenes modernos fue clasificado correctamente (tabla 1). En relación con los especímenes arqueológicos, se observó la misma tendencia que la registrada para las falanges delanteras de modo que, del total de los especímenes arqueológicos analizados, solo 3 fueron clasificados dentro del grupo de guanacos actuales y 21 con las llamas modernas (figura 7B).
Discusión
Los estudios de MG presentados en este artículo permitieron clasificar las falanges arqueológicas procedentes del sitio Quebrada del Cementerio con guanacos y llamas modernas con un elevado grado de precisión, superior al 71 % en el caso de las falanges delanteras, y mayor al 87 % en el caso de las falanges posteriores. En función de ello, los resultados del análisis de la forma de las primeras falanges delanteras y posteriores completas indicaron un amplio predominio de llamas en el sitio (89,4 %), tendencia similar a aquella identificada por las observaciones de MT centradas en el tamaño de la epífisis proximal de estos elementos (94,2 %), aunque con un porcentaje de representación menor (Mercolli 2010, 2004 ).
Los análisis de MT, que incluyen el examen de la muestra de falanges estudiada aquí, se basaron en dos variables métricas y en escasos especímenes modernos de referencia (una vicuña y un guanaco). A partir de ellos se identificó un grupo de tamaño pequeño similar al de las vicuñas actuales, otro de tamaño grande que se aleja del estándar de referencia del guanaco norteño, y que correspondería a las llamas, y un tercer grupo que incluiría especímenes cuyo tamaño es similar tanto al de los guanacos como al de las llamas (Mercolli 2004). En este sentido, se ha señalado la superposición de tamaños existentes entre guanacos andinos grandes y llamas pequeñas (Yacobaccio et al. 1997-1998 ; Yacobaccio 2010) y, por ende, la dificultad de discriminar entre la especie silvestre y doméstica de gran porte. Cabe destacar que diversos autores denominan a este grupo de tamaño como llama-guanaco (López 2003).
Los estudios de MG 2D incluyeron el análisis de una mayor cantidad de camélidos modernos como referencia y de variables de forma. Estos permitieron clasificar a todos los especímenes del grupo de camélidos de mayor porte, incluso aquellos que en las revisiones osteométricas encajan dentro de la categoría llama-guanaco. De esta manera, los estudios de MG 2D facilitan la estimación no solo de la proporción de llamas que se encuentran en el registro del sitio, sino también evaluar la contribución de los guanacos a la subsistencia de las poblaciones humanas con porcentajes de error relativamente bajos. Asimismo, posibilitan la visualización de las variaciones morfológicas registradas en las falanges de estos camélidos.
Por lo tanto, el examen de las variables de forma de elementos poscraneales de guanacos y llamas con técnicas de MG 2D complementa aquellas observaciones de tamaño basadas en la toma de medidas lineales y permite evaluar con mayor detalle y precisión la variabilidad morfológica existente en el grupo de camélidos de mayor porte.
Por otra parte, como se mencionó, el registro arqueológico del sitio estaría dando cuenta de una tendencia en la que claramente el morfotipo similar a las llamas actuales es el dominante. Los resultados muestran una baja representación de especímenes morfológicamente similares a los guanacos actuales en la muestra (10,6 %), aunque dicho porcentaje es superior al estimado con MT (5,8 %). Además, los estudios osteométricos previos sugieren una baja proporción de falanges de tamaño similar a las vicuñas modernas (Mercolli 2004). En suma, los camélidos domésticos son preponderantes en Quebrada del Cementerio, aunque las prácticas de caza de vicuñas y guanacos no fueron abandonadas, sino que continuarían aportando a la subsistencia de los habitantes del sitio. El pastoreo de rebaños de llamas habría sido la estrategia de explotación faunística principal empleada por las poblaciones humanas que habitaron el sitio entre los ca. 900-600 AP. El perfil etario marca un equilibrio entre animales juveniles y adultos, por lo cual las prácticas pastoriles en Quebrada del Cementerio habrían estado centradas en el aprovechamiento de diversos recursos como carne, fibra y animales para carga (Mercolli 2004). La tendencia registrada coincide con lo reportado para otros sitios con ocupaciones humanas contemporáneas o más tardías como el Pucará de Tilcara ( Belotti, López y Otero 2016), Los Amarillos (Mercolli 2016) y La Huerta (Madero 1993, 1991 ).
En este sentido, se ha planteado que para dicho momento cronológico las poblaciones humanas de la región se concentrarían en el valle del río Grande y la porción inferior de las quebradas tributarias, y que se produciría de modo concomitante una intensificación productiva en la cual el pastoreo ocuparía un rol central (Nielsen 1996). A su vez, esto generaría una fuerte presión sobre la estructura de recursos que afectaría el papel de la caza y de los recursos silvestres en el valle principal. La tendencia registrada a partir del estudio de las falanges de Quebrada del Cementerio es acorde con este planteamiento en tanto muestra la centralidad del pastoreo y sugiere que las poblaciones humanas habrían aprovechado temporaria y ocasionalmente áreas muy productivas para la caza de camélidos silvestres (Nielsen 1996).
Consideraciones finales
Los estudios de MG 2D tienen un elevado potencial para diferenciar la forma de las primeras falanges de guanacos y llamas, lo que facilita clasificar a los especímenes arqueológicos de mayor tamaño (Lama sp.) con el grupo de camélidos silvestres o con el grupo de camélidos domésticos. Si bien los datos proporcionados por la MT y la MG no siempre son coherentes, por ejemplo, lo que se ha visto en conjuntos arqueofaunísticos del norte de la Quebrada de Humahuaca, donde las proporciones de las llamas y los guanacos varían según se evalúe su tamaño o su forma (Hernández, L’Heureux y Leoni 2021), en el caso particular de Quebrada del Cementerio se observan resultados consistentes entre ambos tipos de análisis. De esta manera, los estudios de MG 2D confirman la tendencia previamente identificada por los estudios osteométricos, pero permiten alcanzar asignaciones taxonómicas a nivel específico de gran precisión para el grupo de tamaño intermedio, que incluiría guanacos andinos grandes y llamas pequeñas, que a los análisis de MT basados en escasas medidas lineales se les dificulta diferenciar. Por lo tanto, la MG constituye una herramienta de gran utilidad para aquellas investigaciones que buscan describir y explicar el pastoreo de camélidos en la región andina y evaluar la incidencia de la caza en la subsistencia de las poblaciones humanas prehispánicas.
Con este tipo de estudios queda por evaluar un mayor número de arqueofaunas que abarquen segmentos temporales distintos para comparar, más adelante, conjuntos procedentes de sitios localizados en diferentes sectores de la Quebrada de Humahuaca y obtener así tendencias regionales en relación con la proporción de camélidos silvestres y domésticos que aporte al estudio del pastoreo de las poblaciones quebradeñas.

