










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSuma Psicológica
Print version ISSN 0121-4381
Suma Psicol. vol.17 no.2 Bogotá July/Dec. 2010
PATRONES DE ALIMENTACIÓN, SUEÑO Y ACTIVIDAD REPRODUCTIVA EN JERBOS DE MONGOLIA (Meriones unguiculatus)
PATTERNS OF FEEDING, SLEEP AND REPRODUCTIVE BEHAVIOR IN MONGOLIAN GERBILS (Meriones unguiculatus)
Stefani Flórez-Acevedo1, Nora Sarmiento1, Gladys S. Martínez1*
1 Fundación Universitaria Konrad Lorenz, Colombia
* Correspondencia relacionada con este articulo debe ser enviada a:
Gladys S. Martínez
Fundación Universitaria Konrad Lorenz
Carrera 9 bis N. 63-42, Bogotá, Colombia.
Tel: (571) 3472311 Ext. 216, Fax: (571) 3472311 Ext. 131.
e-mail:gladys.martinez@konradlorenz.edu.co.
Recibido: Noviembre 10 2010 Aceptado: Diciembre 15 2010
RESUMEN
Los jerbos de Mongolia son roedores utilizados como excelente modelo biológico. A pesar de esto, su clasificación como especie diurna, nocturna o crepuscular no ha sido clara. Los experimentos que se presentan en este artículo evaluaron patrones de alimentación, sueño-actividad y actividad reproductiva y copulativa en condiciones de luz/oscuridad 12:12 en Jerbos de Mongolia. Los resultados de los experimentos sugieren un patrón nocturno de comportamiento en estos roedores.
Palabras clave: ritmos biológicos; sueño; actividad reproductiva; copula; Jerbos de Mongolia.
ABSTRACT
Mongolian Gerbils are often used as a biological model, but it remains unclear whether these rodents display nocturnal, diurnal, or crepuscular patterns of behavior. The experiments presented below studied patterns of sleep-activity, feeding, and reproductive behavior under 12:12 light dark cycles. All data from these experiments suggest a nocturnal pattern of behavior in these rodents.
Keywords: biological rhythms; sleep; activity; copulatory behavior; Mongolian Gerbils.
RITMOS DE ALIMENTACIÓN, SUEÑO Y ACTIVIDAD REPRODUCTIVA EN JERBOS DE MONGOLIA (Meriones unguiculatus)
Los ritmos biológicos constituyen una relación entre el medio ambiente y el funcionamiento hormonal, comportamental y fisiológico de los seres vivos, permitiendo la adaptación de los individuos a su entorno (Yerushalmi & Green, 2009). Estos ritmos tienen ventajas en la supervivencia de los organismos, en la medida en que aumenta la capacidad para reducir el riesgo de depredación, señalan el momento indicado para que el apareamiento sea exitoso y sincronizan la hora propicia para el consumo de alimentos dependiendo de la disponibilidad de los recursos alimenticios en el ambiente, entre otros (Kronfeld-Schor & Dayan, 2008).
Los ritmos biológicos se presentan en algunos organismos procarióticos y en todos los eucarióticos - desde el hongo hasta los seres humanos- (Chandrashekaran, 1998) y pueden clasificarse como: circadianos, ultradianos o infradianos. Los ritmos circadianos tienen una frecuencia de entre 20 y 28 horas y están presentes en actividades psicofisiológicas como el sueño y la actividad locomotora (Laposky, Bass, Kohsaka & Turek, 2008; Lavie, 2001). Los ritmos ultradianos duran menos de 19 horas, como ocurre con los ritmos respiratorios, alimenticios y cardiacos (Hernández-P. et al., 2004). Finalmente, los ritmos infradianos, se presentan en un período mayor a 29 horas y son característicos de actividades como el ciclo reproductivo; a diferencia de los demás ritmos, éste ha sido poco investigado (Barber, 2010; Diambra & Menna-Barreto, 2004).
Los ritmos biológicos son sincronizados por señales externas o Zeitgebers (ZT) y son controlados por marcapasos o relojes biológicos endógenos que pueden funcionar en ausencia de claves ambientales (Herrero, 2007). El principal ZT es el ciclo de luz/oscuridad, (Challet, Caldelas, Graff & Pevet, 2003; Paul, Saafir & Tosini, 2009): los pulsos de luz ajustan diariamente la fase de inicio y la de finalización de los diferentes ritmos (Corona & Fernández, 2008), generando así patrones de actividad diurnos, nocturnos o crepusculares (Gattermann et al., 2008). Además del ciclo luz/oscuridad, se han identificado otros ZTs, como la temperatura, la humedad, la disponibilidad del alimento y los intercambios sociales (Schulz & Steimer, 2009; Vieira & Morato, 2005).
El principal reloj biológico de los mamíferos es el Núcleo Supraquiasmático (NSQ) localizado en el hipotálamo, en ambos lados de la línea media y por encima del quiasma óptico (Hastings, 1998; Piggins, 2002; Schibler, 2005; Weinert, 2005). En general una lesión o destrucción del NSQ produce la pérdida de casi todos los ritmos, aunque pueden persistir algunos (Barnard & Nolan, 2008; Hastings, 1998; Karatsoreos & Silver, 2007; Rusak & Zucker, 1979; Satinoff & Prosser, 1988; Wachulec, Li, Tanaka, Peloso & Satinoff, 1997). Estas lesiones también pueden suprimir algunos ritmos ultradianos (Szafarczyk, Ixart, Malaval, Nouguier-Soule & Assenmacher, 1979) o alterar ritmos infradianos de reproducción en la mayoría de los mamíferos (Hofman, 2004).
El NSQ recibe información fotópica través de vías aferentes desde la retina (Challet, 2007; Corona, Juárez, Zabaleta & Caba, 2008; Schulz & Steimer, 2009). Esta información es luego transmitida al resto del organismo para organizar los procesos fisiológicos rítmicamente. La principal vía eferentes se localiza en la porción ventral del núcleo paraventricular (Corona et al., 2008). Si esta zona es lesionada se presenta una pérdida selectiva del ritmo circadiano de actividad-descanso (Abrahamson & Moore, 2006). El NSQ influye también en los ritmos de liberación de hormonas al enviar señales a glándulas del sistema endocrino como la glándula pineal y a órganos periféricos como el hígado, la tiroides y las glándulas suprarrenales (Corona et al., 2008).
Además del NSQ, existen genes sincronizadores en otras estructuras del organismo que participan en la regulación de ritmos como la alimentación (Martínez, 2009; Stephan, 2002; Takeda & Maemura, 2010). Estos genes sincronizadores hacen parte del sistema circadiano periférico (Laposky et al., 2008; Schibler, 2005) que en contraste con el NSQ, parece ser sincronizado por factores neuronales derivados del Sistema Nervioso Central (SNC; (Takeda & Maemura, 2010) y estos, a diferencia del NQS, intervienen de manera limitada dentro de algunos ritmos. La mayoría de estudios sobre ritmos biológicos se han centrado en ritmos circadianos y en un número limitado de modelos animales, generalmente mamíferos nocturnos. Si bien resulta ventajoso trabajar con estas especies, ello implica que aún se desconocen mecanismos neurales de patrones comportamentales en especies diurnas, igualmente relevantes para el progreso de ésta área (McElhinny, Smale & Holekamp, 1997).
Dentro de las especies nocturnas más estudiadas se encuentran la rata Wistar (Rattus norvegicus; (Vosko, Hagenauer, Hummer & Lee, 2009) y el hámster dorado (Refinetti, 1999). Entre las especies diurnas estudiadas se encuentran la ardilla de pradera manto-dorado (Spermophilus lateralis), el degus (Octodon degus) y la rata de hierba africana (Arvicanthis niloticus; (Smale, Lee & Nunez, 2003). Sin embargo, existen otras especies utilizadas como modelos de investigación cuya clasificación según sus patrones fisiológicos y comportamentales no ha sido posible o ha tenido pocos avances. Este es el caso de los jerbos de Mongolia (Meriones Unguiculatus), roedores nativos del noroeste de China y Mongolia que pertenecen a la familia erbiliana (Karakas & Gunduz, 2002).
Los jerbos son socialmente monógamos, viven en grupo con una organización social basada en una pareja estable reproductiva (Razzoli & Valsecchi, 2006) y exhiben conducta biparental (Wynne-Edwards, 2007). Esta especie ha sido utilizada en investigaciones relacionadas con reproducción y alimentación (Karakas, Camsari, Serin & Gunduz, 2005), enfermedades inflamatorias intestinales (Bleich, Martin, Bleich & Klos 2010) y en lesiones cerebrales (Radenovic, Selanovic & Andjus, 2008), entre otras. Si bien se han realizado algunos estudios, poco se conoce acerca de sus ritmos biológicos (Weinert, Weinandy & Gattermann, 2007), debido principalmente a resultados contradictorios reportados en la literatura: bajo distintas condiciones de laboratorio muestran patrones de actividad diurnos, nocturnos o crepusculares (Demas, El-Bakry, Mintz, Zarhan & Bartness, 2001). Por ejemplo, Susic & Masirevic, (1986) analizaron tanto registros comportamentales como electroencefalográficas las 24 horas con ciclos de luz/ oscuridad (12 L:12 D) durante 48 horas y encontraron que los jerbos en cautiverio son animales crepusculares, mostrando picos de actividad en las transiciones entre la luz y la oscuridad. Weinert et al. (2007), por su parte, evaluaron patrones de actividad de los jerbos bajo dos parámetros diferentes de foto-periodo (L: D= 14:10 y 10:14 h) e incluyeron dos condiciones ambientales: uno con acceso a la rueda y otro sin acceso a la rueda y encontraron que los patrones de actividad en Jerbos eran bimodales si estos no tenían acceso a la rueda, con picos de actividad en la primera mitad del tiempo de exposición a la luz y en la transición entre la luz y la oscuridad; por el contrario, cuando tenían acceso a la rueda, sus picos de actividad ocurrían principalmente durante el periodo de oscuridad. Otra investigación en la misma línea, evaluó en períodos de 24 h la frecuencia locomoción, excavación, acicalamiento, alimentación e ingesta de líquidos. Los jerbos exhibieron patrones crepusculares con picos entre las 6 y 8 am y las 6 y 8 pm, en condiciones de luz natural (Pietrewicz, Hoff & Higgins, 1982).
Al comparar los resultados de los estudios mencionados, se encuentran divergencias que pueden corresponder a la metodología y procedimientos en la medición de la actividad locomotora (Demas et al., 2001). En un intento por definir más claramente los ritmos en Jerbos de Mongolia, en el presente estudio se evalúan patrones de sueño - actividad, reproducción e ingesta en esta especie.
MÉTODO GENERAL
Sujetos
En el estudio se utilizaron en total 27 jerbos de Mongolia adultos, criados en el laboratorio de Psicobiología de la Fundación Universitaria Konrad Lorenz, cuyos padres fueron originalmente adquiridos en el Instituto Nacional de Salud. 11 fueron usados para evaluar patrones de alimentación, 4 para evaluar patrones de sueño-actividad y 6 parejas (6 hembras y 6 machos) para el estudio de patrones de reproducción. Los sujetos se alojaron en jaulas de confinamiento marca Tecniplast, fabricadas en policarbonato (41.91 x 20.32 x20.32 cm), en cuartos experimentales con temperaturas de 23 ± 2 °C, humedad relativa entre 40 y 59 % y un ciclo de luz/oscuridad de 12 h (12:12 LD), con las luces encendidas a las 6:00 am (ZT 0) y apagadas a las 6:00 pm (ZT 12). Los sujetos recibieron alimento y agua ad libitum durante todo el estudio. Para realizar los experimentos se siguieron las especificaciones establecidas en la Guía para el cuidado y Uso de los animales de Laboratorio (National Research Council, 2010).
EXPERIMENTO 1 RITMOS DE INGESTA
Método
11 jerbos (6 hembras y 5 machos) fueron alojados individualmente en jaulas. Diariamente se suministró alimento (previamente pesado) a cada sujeto y se registró cada 3 horas el peso del alimento no consumido, durante las 24 horas del día en un período de 15 días. Se utilizó la prueba t de Student para determinar diferencias en el consumo de alimento entre el día y la noche. Para determinar si existían diferencias significativas entre el consumo de alimento en los diferentes ZTs se utilizó la prueba F.RESULTADOS
El registro del consumo de alimento de los 11 sujetos durante los 15 días, mostró una diferencia significativa entre la cantidad de comida ingerida entre el día y la noche (t 1, 10 = 8.14, p < 0.05), con un mayor consumo cuando las luces estaban encendidas (Figura 1).

Al comparar el consumo de alimento por cada ZT, se encontraron diferencias significativas entre ZT0 y los demás ZTs (3, 6, 9, 12, 15). Así mismo se encontraron diferencias en el consumo en el ZT (18), frente a los demás ZTs (Figura 2).

EXPERIMENTO 2 RITMOS DE SUEÑO Y ACTIVIDAD
Método
4 jerbos (2 hembras y 2 machos) fueron ubicados individualmente. Su comportamiento fue video grabado continuamente durante 15 días para lo cual se utilizaron cámaras Sony (VZ-205D) con lente de 3.6 milímetros (mm), potencia DC12V, 30 leds y 420TVL de resolución horizontal.
Para evaluar los videos se tuvo en cuenta el procedimiento realizado por (Novak, Smale & Nunez, 1999), en el que cada hora fue dividida en 12 intervalos de 5 minutos y el comportamiento de los animales fue registrado el primer minuto de cada intervalo las 24 h. Los animales se catalogaron como dormidos si se encontraban quietos, enrollados con la cabeza baja o recostados sobre alguno de sus lados. Se consideró que estaban despiertos si permanecían en la misma posición presentando algún tipo de movimiento, caminaban, corrían, saltaban, bebían, comían, se colgaban de las tapas de las jaulas, se acicalaban o emitían alguna conducta estereotipada. Para el análisis de datos, el número de intervalos de 5 min en el que se registraba sueño fue totalizado para cada ZT por cada animal. Estos datos fueron utilizados para determinar el promedio de presentación de conducta de sueño para el período de luz y el período de oscuridad por cada animal, en cada día de grabación. Se utilizó la prueba t de Student para muestras relacionadas para comparar la frecuencia de sueño que se presenta durante el día y durante la noche y para comparar las medias entre cada ZT.
Dos observadores llevaron a cabo el registro de información y el índice de acuerdo entre ellos fue de 94.4%.
RESULTADOS
Se evidenció una disminución en el promedio de la frecuencia registrada en las conductas de sueño en el ZT 15 indicando posible aumento en la actividad locomotora. La cantidad promedio de sueño registrada durante el día fue constante.
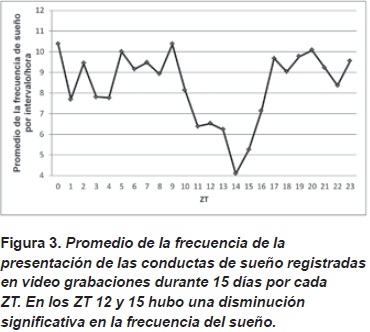
Al compara lo los datos para los ZTs en los que se hicieron los registros de los patrones de alimentación (Figura 4) se encontraron diferencias significativas en los ZT 12 y ZT15 con respecto a los demás ZTs.

EXPERIMENTO 3 RITMOS DE ACTIVIDAD REPRODUCTIVA Y COPULATIVA POST-PARTO
Método
6 parejas (macho y hembra) se situaron en sus respectivas jaulas y se filmaron las 24 horas del día, desde el momento en que se ubicaron en parejas hasta el nacimiento del segundo parto. Para determinar la hora del parto, los animales eran observados diariamente y cuando se encontraba una nueva camada, se revisaba la video grabación de ese día y se registraba la hora en la que se observaba la primer cría como la hora del parto ± 1 minuto (McElhinny et al., 1997). Las crías fueron removidas a los 25. Para registrar la actividad copulativa post-parto se utilizó el programa Etholog 2.2 (Ottoni, 2000), incluyendo las conductas de olfateo a genitales, escape de monta, intento de monta y monta.
RESULTADOS
Como lo muestra la Figura 5, la mayoría de los partos ocurrieron a lo largo de la fase de luz, con sólo un parto al inicio de la fase de oscuridad.

La actividad copulativa post-parto se inició en promedio 6±20 h después del parto en todas las parejas.
Discusión
El objetivo de esta investigación fue determinar los ritmos biológicos de los Jerbos de Mongolia bajo condiciones específicas de laboratorio. Aunque los datos obtenidos no son concluyentes, sí sugieren que los patrones de actividad de esta especie presentan una tendencia nocturna. Si bien existen estudios anteriores que apoyan estos hallazgos, otros han arrojado conclusiones diferentes (Pietrewicz et al., 1982; Susic & Masirevic, 1986) lo cual puede obedecer tanto a variables metodológicas, en cuanto a las conductas que se eligen para ser observadas y la forma en que se realizan sus registros, como a factores ambientales, teniendo en cuenta que estos animales son estacionarios (Karakas et al., 2005).
Al analizar los resultados se observa mayor consumo de alimento durante el día (experimento 1). Sin embargo, datos del experimento 2 en el que se evalúan patrones de sueño muestran una frecuencia mayor del sueño durante la fase nocturna. La contradicción aparente en los datos puede estar relacionada con la estrategia utilizada para la recolección de información sobre el consumo, que involucraba el ingreso de los investigadores cada tres horas al cuarto de alojamiento. Un registro con un sistema que no implique perturbaciones regulares puede arrojar datos diferentes.
El patrón nocturno sugerido en el experimento 2, parece confirmarse cuando se evalúan los patrones de reproducción: mientras que los partos ocurrieron en períodos de poca actividad, lo cual concuerda lo que suele ocurrir en todas las especies mamíferas (Rowland, Helgeson & Cox, 1984), el apareamiento post-parto se produjo en periodos de actividad en los cuales los episodios de sueño son poco frecuentes.
Los resultados anteriores concuerdan con datos de otras investigaciones que hacen referencia a la presencia de patrones nocturnos en jerbos de Mongolia (Karakas et al., 2005). Es importante sin embargo tener en cuenta que los resultados obtenidos en el presente estudio están basados en condiciones de laboratorio específicas (temperaturas de 23 ± 2 °C, humedad relativa entre 40 y 59 % y un ciclo de luz/oscuridad de 12 h (12:12 LD), y que los patrones de comportamiento aquí descritos puedan estar asociados a factores ambientales, como lo sugiere (Weinert et al., 2007). Registros automáticos de la actividad y del consumo de alimento, se hacen necesarios para confirmar los datos aquí obtenidos.
REFERENCIAS
Abrahamson, E. E. & Moore, R. Y. (2006). Lesions of suprachiasmatic nucleus efferents selectively affect rest-activity rhythm. Molecular and Cellular Endocrinology, 252, 46-56. [ Links ]
Barber, C. F. (2010). A review and applications of monthly circadian rhythms. Journal of Psychiatric and Mental Health Nursing, 17, 274-279. [ Links ]
Barnard, A. R., & Nolan, P. M. (2008). When clocks go bad: neurobehavioural consequences of disrupted circadian timing. PLoS Genetics, 4, e1000040. [ Links ]
Bleich, E. M., Martin, M., Bleich, A. & Klos, A. (2010). The Mongolian gerbil as a model for inflammatory bowel disease. International Journal of Experimental Pathology, 91, 281-287. [ Links ]
Challet, E. (2007). Minireview: Entrainment of the suprachiasmatic clockwork in diurnal and nocturnal mammals. Endocrinology, 148, 5648-5655. [ Links ]
Challet, E., Caldelas, I., Graff, C. & Pevet, P. (2003). Synchronization of the molecular clockwork by light- and food-related cues in mammals. Biological Chemistry, 384, 711-719. [ Links ]
Chandrashekaran, M. (1998). Biological rhythms research: A personal account. Journal of Biosciences, 23, 545-555. [ Links ]
Corona, A. & Fernández, M. (2008). Bases de los ritmos Biológicos. In M. Caba (Ed.), Bases celulares y moleculares de los ritmos biológicos (pp. 15-24). México: Sociedad Mexicana de Ciencias Fisiológicas. [ Links ]
Corona, A., Juárez, C., Zabaleta, Y. & Caba, M. (2008). Vías de sincronización Fótica y Fisiológica del Núcleo Supraquiasmático. In M. Caba (Ed.), Bases celulares y moleculares de los ritmos biológicos (pp. 39-61). México: Sociedad Mexicana de Ciencias Fisiológicas. [ Links ]
Demas, G. E., El-Bakry, H. A., Mintz, E. M., Zarhan, W. M. & Bartness, T. J. (2001). Wheel-running activity patterns of five species of desert rodents. Biological Rhythm Research, 32, 1-16. [ Links ]
Diambra, L., & Menna-Barreto, L. (2004). Infradian rhythmicity in sleep/wake ratio in developing infants. Chronobiology International, 21, 217-227. [ Links ]
Gattermann, R., Johnston, R. E., Yigit, N., Fritzsche, P., Larimer, S., Ozkurt, S. et al. (2008). Golden hamsters are nocturnal in captivity but diurnal in nature. Biology Letters, 4, 253-255. [ Links ]
Hastings, M. (1998). The brain, circadian rhythms, and clock genes. British Medical Journal, 317, 1704-1707. [ Links ]
Hernández-P. , R., Cadena-Orea, D., Talavera Recoder, L. J., Samano-Aviña, M. G., Aparicio llamas, M., Ochoa-Romero, G. F. et al. (2004). Ritmos biológicos. Variabilidad Horaria en Nacimientos Humanos (1a parte). Revista de Especialidades Médico-Quirúrgicas, 9, 37-42. [ Links ]
Herrero, M. (2007). Ritmos de Actividad Motora, Comportamiento Alimentario e Influencia de la Melatonina exógena en peces Teleósteos. Tesis Doctoral, Universidad de Murcia, Murcia, España. [ Links ]
Hofman, M. A. (2004). The brain's calendar: neural mechanisms of seasonal timing. Biological reviews of the Cambridge Philosophical Society, 79, 61-77. [ Links ]
Karakas, A., Camsari, C., Serin, E. & Gunduz, B. (2005). Effects of photoperiod and food availability on growth, leptin, sexual maturation and maintenance in the Mongolian gerbils (Meriones unguiculatus). Zoological Science, 22, 665-670. [ Links ]
Karakas, A., & Gunduz, B. (2002). Effect of different photoperiods on gonadal maintenance and development in Mongolian gerbils (Meriones unguiculatus). Zoological Science, 19, 233-239. [ Links ]
Karatsoreos, I. N., & Silver, R. (2007). Minireview: The neuroendocrinology of the suprachiasmatic nucleus as a conductor of body time in mammals. Endocrinology, 148, 5640-5647. [ Links ]
Kronfeld-Schor, N. & Dayan, T. (2008). Activity patterns of rodents: the physiological ecology of biological rhythms. Biological Rhythm Research, 39, 193-211. [ Links ]
Laposky, A. D., Bass, J., Kohsaka, A. & Turek, F. W. (2008). Sleep and circadian rhythms: key components in the regulation of energy metabolism. FEBS Letters, 582(1), 142-151. [ Links ]
Lavie, P. (2001). Sleep-wake as a biological rhythm. Annual Review of Psychology, 52, 277-303. [ Links ]
Martinez, G. S. (2009). Regulación circadiana del comportamiento: diferencias entre especies diurnas y nocturnas. Universitas Psychologica, 8, 487-496. [ Links ]
McElhinny, T. L. Smale, L., & Holekamp, K. E. (1997). Patterns of body temperature, activity, and reproductive behavior in a tropical murid rodent, Arvicanthis niloticus. Physiology & behavior, 62, 91-96. [ Links ]
National Research Council. (2010). Guide for the Care and Use of Laboratory Animals. Washington, DC.: National Academy Press. [ Links ]
Novak, C. M., Smale, L., & Nunez, A. A. (1999). Fos expression in the sleep-active cell group of the ventrolateral preoptic area in the diurnal murid rodent, Arvicanthis niloticus. Brain Research, 818, 375-382. [ Links ]
Ottoni, E. B. (2000). EthoLog 2.2: a tool for the transcription and timing of behavior observation sessions. Behavior Research Methods, Instruments & Computers, 32, 446-449. [ Links ]
Paul, K. N., Saafir, T. B. & Tosini, G. (2009). The role of retinal photoreceptors in the regulation of circadian rhythms. Reviews In Endocrine & Metabolic Disorders, 10, 271-278. [ Links ]
Pietrewicz, A. T., Hoff, M. P. & Higgins, S. A. (1982). Activity rhythms in the Mongolian gerbil under natural light conditions. Physiology & Behavior, 29, 377-380. [ Links ]
Piggins, H. D. (2002). Human clock genes. Annals of Medicine, 34, 394-400. [ Links ]
Radenovic, L., Selanovic, V. & Andjus, P. (2008). Neuroprotection by MK-801 following cerebral ischemia in Mongolian gerbils. Archives of Biological Sciences, 60, 341-346. [ Links ]
Razzoli, M. & Valsecchi, P. (2006). Different social bonds produce differential effects on behaviour and physiology in Mongolian gerbils. Ethology Ecology & Evolution, 18, 289-306. [ Links ]
Refinetti, R. (1999). Relationship between the daily rhythms of locomotor activity and body temperature in eight mammalian species. American Journal of Physiology, 277, R1493-1500. [ Links ]
Rowland, D. L., Helgeson, V. S. & Cox, C. C., 3rd. (1984). Temporal patterns of parturition in mammals in captivity. Chronobiologia, 11, 31-39. [ Links ]
Rusak, B. & Zucker, I. (1979). Neural regulation of circadian rhythms. Physiological Reviews, 59, 449-526. [ Links ]
Satinoff, E. & Prosser, R. A. (1988). Suprachiasmatic nuclear lesions eliminate circadian rhythms of drinking and activity, but not of body temperature, in male rats. Journal of Biological Rhythms, 3, 1-22. [ Links ]
Schibler, U. (2005). The daily rhythms of genes, cells and organs. Biological clocks and circadian timing in cells. EMBO Report, 6 Spec No, S9-13. [ Links ]
Schulz, P. & Steimer, T. (2009). Neurobiology of circadian systems. CNS Drugs, 23 Suppl 2, 3-13. [ Links ]
Smale, L., Lee, T. & Nunez, A. A. (2003). Mammalian diurnality: some facts and gaps. Journal of Biological Rhythms, 18, 356-366. [ Links ]
Stephan, F. K. (2002). The "other" circadian system: food as a Zeitgeber. Journal of Biological Rhythms, 17, 284-292. [ Links ]
Susic, V. & Masirevic, G. (1986). Sleep patterns in the Mongolian gerbil, Meriones unguiculatus. Physiology & Behavior, 37, 257-261. [ Links ]
Szafarczyk, A., Ixart, G., Malaval, F., Nouguier-Soule, J. & Assenmacher, I. (1979). Effects of lesions of the suprachiasmatic nuclei and of p-chlorophenylalanine on the circadian rhythms of adrenocorticotrophic hormone and corticosterone in the plasma, and on locomotor activity of rats. Journal of Endocrinology, 83, 1-16. [ Links ]
Takeda, N. & Maemura, K. (2010). Circadian clock and vascular disease. Hypertension Research, 33, 645-651. [ Links ]
Vieira, E. & Morato, M. (2005). Longitudinal study of the spectral composition of behavioral rhythms in the rat. Biological Rhythm Research, 36, 131-140. [ Links ]
Vosko, A. M., Hagenauer, M. H., Hummer, D. L. & Lee, T. M. (2009). Period gene expression in the diurnal degu (Octodon degus) differs from the nocturnal laboratory rat (Rattus norvegicus). American Journal of Physiology -Regulatory, Integrative and Comparative Physiology, 296, R353-361. [ Links ]
Wachulec, M., Li, H., Tanaka, H., Peloso, E. & Satinoff, E. (1997). Suprachiasmatic nuclei lesions do not eliminate homeostatic thermoregulatory responses in rats. Journal of Biological Rhythms, 12, 226-234. [ Links ]
Weinert, D. (2005). The temporal order of mammals. Evidence for multiple central and peripheral control mechanisms and for endogenous and exogenous components: some implications for research on aging. Biological Rhythm Research, 36, 293-308. [ Links ]
Weinert, D., Weinandy, R. & Gattermann, R. (2007). Photic and non photic effects on the daily activity pattern of Mongolian gerbils. Physiology & Behavior, 90, 325-333. [ Links ]
Wynne-Edwards, K. E. (2007). Paternal care in rodents: Weakening support for hormonal regulation of the transition to behavioral fatherhood in rodent animal models of biparental care. Hormones and Behavior, 52, 114-121. [ Links ]
Yerushalmi, S. & Green, R. M. (2009). Evidence for the adaptive significance of circadian rhythms. Ecology Letters, 12, 970-981. [ Links ]

