










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.30 no.1 Santa Marta Jan./Dec. 2001
CARACTERIZACION ECOLOGICA DE LAS ASOCIACIONES DE PECES DEMERSALES DEL CARIBE COLOMBIANO NORTE, CON ENFASIS EN LOS PARGOS (LUTJANIDAE)
ECOLOGICAL CARACTERIZARION OF THE ENSAMBLAGES OF DEMERASAL FISHES IN THE COLOMBIAN CARIBBEAN, WITH EMPHASIS IN SNAPPERS (LUTJANIDAE)
Luis. Manjarrés1, Camilo García2 y Arturo Acero3
1 Instituto nacional de pesca y acuicultura INPA. Santa Marta, Colombia. E-mail: propargos@celcaribe.net.co (LM).
2 Universidad Nacional de Colombia, Departamento de biología - INVEMAR. A.A. 1016. Cerro Punta Betín , Santa Marta, Colombia. E-mail: cgarcia@invemar.org.co (C.B.G.).
3 Universidad Nacional de Colombia. Instituto de Ciencias Naturales - INVEMAR. A.A. Cerro Punta Betin, Santa Marta, Colombia (A.A.P.).
RESUMEN
El presente estudio se centró en los objetivos de establecer la existencia de asociaciones de peces demersales en la plataforma y el talud superior de la zona norte del Caribe colombiano, y determinar si los patrones bióticos determinados por estas asociaciones se correlacionan con los patrones estructurados por las condiciones ambientales evaluadas. Los datos del estudio provinieron de tres campañas de evaluación de recursos demersales del Programa INPA-VECEP/UE, efectuadas durante el período 1995-1996. La característica dominante de los grupos de estaciones y de las asociaciones de peces demersales fue la carencia de persistencia temporal. Únicamente fue factible establecer la persistencia de algunas asociaciones parciales durante un máximo de dos períodos de muestreo, como fue el caso de Rhomboplites aurorubens/Priacanthus arenatus, Lutjanus analis/Lutjanus synagris y Eucinostomus argenteus/Upeneus parvus. Se presentaron cambios estacionales en el conjunto de variables ambientales que explicaron de mejor forma la estructura comunitaria, a excepción de la profundidad, que siempre figuró en la combinación que produjo la correlación óptima. Los cambios en los patrones bióticos parecen señalar que la co-existencia de las especies se debe básicamente a una respuesta común a los gradientes ambientales, antes que a interacciones entre especies. Sin embargo, el hecho de que estas correlaciones fuesen relativamente bajas parece indicar que a una escala de tiempo menor otros factores, tales como posibles migraciones por razones biológicas, también podrían afectar la estructura y composición de las asociaciones de peces demersales.
PALABRAS CLAVE: Demersales, Caribe colombiano, Ensamblajes, Ecología, Lutjanidae.
ABSTRACT
The mean objectives of the present study were to establish the existence of demersal fish associations in the shelf and superior slope of the colombian Caribbean northern zone and determine if the biotical patterns determineted by these associations are correlationed with the patterns structured by the environmental conditions evaluated. Data were taken from three demersal resources assessment surveys of the INPA-VECEP/UE Program, carried out during the period 1995-1996. The station groups were not consistent in terms of species composition along the different periods. The dominant characteristic of demersal species associations was the abcense of temporal persistence. Only It was possible to find some partial associations which remain during a maximum of two sampling periods: Rhomboplites aurorubens/Priacanthus arenatus, Lutjanus analis/Lutjanus synagris y Eucinostomus argenteus/Upeneus parvus. It were observed seasonal changes in the set of environmental variables which better explain the community structure, but depth always appears in the optimal correlation set. Apparently, species co-existence is rather due to a common answer to environmental gradients, than to species interactions. Nevertheless, the relatively low correlations seem to indicate that other factors, like migrations for biological reasons, could also affect the structure and composition of demersal fish associations, on a smaller time scale.
KEY WORDS: Demersal, Colombian Caribbean, Assemblages, Ecology, Lutjanidae.
INTRODUCCION
Hay una tendencia general, tanto en aguas tropicales como templadas, a desarrollar métodos para el manejo de sistemas multiespecíficos. Para ello, resulta ideal definir las asociaciones de peces demersales tal manera que se minimicen las influencias externas de otras especies y se logre una persistencia temporal en sus estructuras (Mahon y Smith, 1989). En la mayoría de los estudios que han discutido la persistencia de dichas asociaciones, se ha encontrado que éstas son espacial y temporalmente dinámicas (Arancibia, 1992; Bianchi, 1992a,c; Ansari et al., 1995; Fariña et al., 1997). Esta variabilidad ha puesto en discusión la real existencia de estas asociaciones. En el caso de los peces demersales de plataformas continentales, el problema generalmente se ha abordado usando análisis de clasificación y/o ordenación de datos de arrastres de fondo, los que permiten determinar grupos de especies que tienden a coincidir a lo largo de un grupo de estaciones, usualmente relacionados con cambios graduales en las condiciones ambientales (IMR, 1989; Bianchi, 1991; Bianchi, 1992a,b,c; Clarke y Warwick, 1994; Wolff, 1996).
Otros estudios recientes enfatizan el carácter dinámico de las agregaciones de peces y en el hecho de que en un momento dado no sólo responden a los gradientes ambientales, sino también a factores reproductivos (migraciones o concentraciones para el desove) o tróficos (migraciones por alimento), de forma que los factores ambientales y los biológicos se combinan para producir asociaciones estacionalmente variables. No obstante, parece que este tipo de correlaciones podrían explicar comportamientos a una escala temporal menor que los derivados del análisis de los factores ambientales (Caddy y Sharp, 1988; Wolff, 1996).
A pesar de la importancia biológica y pesquera de la zona norte del Caribe colombiano, son pocos las investigaciones sobre las características ecológicas de las asociaciones de peces demersales, su persistencia temporal y su distribución espacial. Aunque se registra un rápido crecimiento en la cantidad de literatura relacionada con la taxonomía, la dinámica poblacional y las pesquerías de la región, aún es muy limitado el conocimiento de las asociaciones de peces demersales y sus respuestas a los gradientes ambientales. Si bien existe información puntual sobre algunos de estos aspectos, sólo se encuentran dos trabajos en los que se han llevado a cabo análisis conjuntos de dicha información, en tiempo y en espacio (IMR, 1989; Duarte y von Schiller, 1997).
El enfoque integrado de las investigaciones sobre asociaciones de peces demersales y el marco físico ambiental donde se organizan biológicamente ha tenido un claro desarrollo en los últimos años. Cada vez es mayor la tendencia a valorar la coincidencia de especies como debida a variaciones en parámetros abióticos, tales como profundidad, temperatura y/o calidad del sedimento del fondo (Caddy y Sharp, 1988; Bianchi, 1992c). Por ello, el presente estudio se fundamentó en torno a las hipótesis de que, en primera instancia, era factible establecer tales asociaciones en la plataforma y el talud superior de la zona norte del Caribe colombiano y, en segundo lugar, que las mismas eran determinadas por una respuesta común a los gradientes ambientales.
AREA DE ESTUDIO
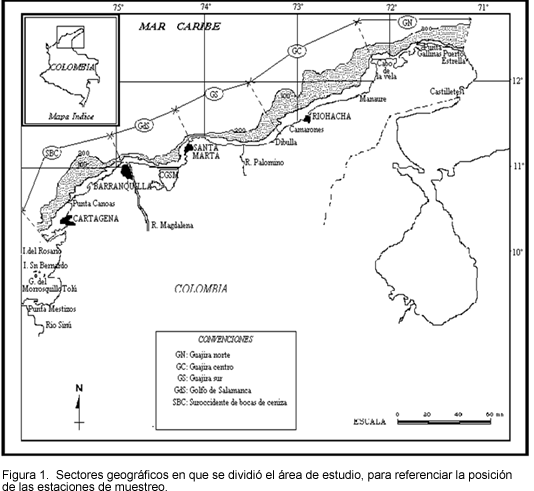
El área de estudio se ubica en la plataforma caribeña colombiana, entre las longitudes 71°14´ (frente a Puerto Estrella) y 75° 45´ W (frente a Islas del Rosario). En sentido costa-mar, el área de cobertura de las campañas oceanográficas (2729 mn²) se extendió desde la actual frontera pesquera artesanal, (alrededor de los 13 m de profundidad, en el sector más somero), y la cota batimétrica de los 200 m (Figura 1). Aunque de manera convencional la isolínea de profundidad de los 200 m define el límite externo de la plataforma continental, para el área de estudio este límite se encuentra realmente a menor profundidad. Según Vernette (1985), este límite se halla generalmente entre
130 y 150 m. No obstante, los registros batimétricos hechos en desarrollo de las campañas oceanográficas INPA-VECEP/UE indican que una característica recurrente es encontrar el inicio del talud a los 100 m de profundidad (Manjarrés et al., 1995, 1996a,b).
Tanto desde el punto de vista meteorológico como del oceanográfico, estacionalmente se presentan cambios muy pronunciados (Blanco, 1988). Los vientos Alisios del noreste (a través de la surgencia) y la descarga de aguas continentales caracterizan las estaciones de la región y condicionan la productividad de los ecosistemas que sustentan un alto porcentaje de los recursos pesqueros artesanales del área de estudio (Márquez, 1990; CORPES 1992).
En razón a la condición de paralelismo que debe existir entre la orientación de la costa y la dirección del viento, la extensión espacial de la surgencia se limita básicamente al tramo costero ubicado entre Punta Gallinas (alta Guajira), al oriente, y el Cabo de la Aguja (sector de Santa Marta), al occidente (72-74oW). Se presentan dos centros de intensidad máxima o núcleos del afloramiento: al norte de la Península de La Guajira (entre el Cabo de la Vela y Punta Gallinas) y frente al cabo de La Aguja, en el sector de Santa Marta (Donoso, 1992).
En lo que respecta a las descargas continentales, son varias las que fertilizan estacionalmente el área de estudio, determinando cambios en las condiciones físico-químicas del agua (principalmente la salinidad y la turbidez), y constituyendo un factor determinante en las condiciones de vida de los organismos (Márquez, 1982). Se destacan los aportes del río Magdalena -el mayor en drenar al Mar Caribe, a través de Bocas de Ceniza, con 650 m3/s, según Blanco (1988)-, la Ciénaga Grande de Santa Marta -CGSM- (167.2 m3/s), el Canal del Dique y los vertimentos de los ríos menores de la Sierra Nevada de Santa Marta, los cuales originan pequeños sectores estuarinos desde el Parque Natural Tayrona hasta la Guajira (CORPES, 1992).
El efecto diferencial de los vientos, las corrientes y surgencias resultantes y las descargas continentales permiten configurar entonces cuatro épocas climáticas para el área de estudio, las cuales en el presente trabajo se referencian de la siguiente manera: seca o de verano mayor (mediados de diciembre a abril), lluviosa menor (mayo-junio), seca menor (julio-mediados de agosto) y lluviosa mayor (mediados de agosto a mediados de diciembre).
Diversos autores han planteado la existencia de diferencias ecológicas a lo largo del área de estudio. En CORPES (1992) se menciona la ocurrencia de dos situaciones de carácter general en el ambiente del área: condiciones típicas de un mar tropical (mar cálido, con arrecifes coralinos y manglares como ecosistemas característicos) hacía la parte suroccidental, principalmente en cercanías de Islas del Rosario; y condiciones similares a las de aguas subtropicales en la zona frente a los departamentos Magdalena y Guajira (áreas fértiles sometidas a fuertes ciclos estacionales).
Márquez (1990) propone un esquema de regionalización ecológica que incorpora mayor información ambiental, el cual considera un sector intermedio, que denomina Región II, desde Punta Gloria hasta Punta Galeras (Atlántico), influido por las descargas fértiles y sedimentos del río Magdalena y la CGSM. Por tanto, dentro del área de estudio los otros dos sectores serían Guajira-Punta Gloria (Región I) y Punta Galeras-Islas del Rosario (parte de la Región III). El primero se caracteriza por la influencia directa de la corriente Caribe y la surgencia estacional fertilizante. El segundo es una zona de "transición" hidrográfica y climática modificada por la contracorriente proveniente del Golfo de Urabá (Andrade et al., 1988).
MATERIALES Y METODOS
Procedencia de los datos
Los datos provinieron de las campañas oceanográficas de evaluación de recursos demersales efectuadas por el Programa INPA-VECEP/UE durante el período 1995-1996, dos a bordo del B/I ANCON y una final a bordo del B/O "HERMANO GINES" (Tabla 1). Las campañas se inscribieron en las siguientes temporadas climáticas: seca menor (período 1), lluviosa mayor (período 2) y seca mayor (período 3). Los resultados también permiten caracterizar cada uno de estos períodos en función de la intensidad relativa del afloramiento, así: 1: período de afloramiento débil, 2: período de ausencia de afloramiento, y 3: período de afloramiento fuerte (Manjarrés et al., 1995; Manjarrés et al., 1996a,b).
Para evitar sesgos por diferencias en la capturabilidad y el esfuerzo, el tiempo efectivo del arrastre (red sobre el fondo) se procuró estandarizar en 30 minutos y la velocidad en 3.5 nudos. Se trabajó con la relación longitud cable/profundidad recomendada para lograr una abertura horizontal de la red del 60%. Debido a las inconsistencias del muestreo nocturno, derivadas de la migración vertical de especies demersales (Doubleday, 1981; Bartels et al., 1983), las estaciones de pesca consideradas en el estudio se circunscribieron a horas diurnas.

Preparación de los datos
Inicialmente, se seleccionaron las especies demersales, con base en los trabajos de Cervigón et al. (1992), Cervigón (1991, 1993, 1994, 1996) y los catálogos de especies de la FAO (Collette y Nauen, 1983; Compagno, 1984; Allen, 1985) . En vista de la dificultad de escoger las especies "realmente demersales", sólo se excluyeron del análisis las reconocidamente semipelágicas -algunas veces referenciadas como bentopelágicas-, las cuales, a pesar de que están relacionadas con la cadena trófica asociada al fondo, no son cuantitativamente representativas de la abundancia relativa de especies en la comunidad (Bianchi, 1992c). Además, en la tercera campaña se usó una red con una abertura vertical significativamente mayor que la empleada en las dos campañas previas, lo que implica diferencias apreciables en la capturabilidad de las especies excluidas. Para la preparación de las tres matrices de abundancia de especies por estaciones, el siguiente paso fue calcular las abundancias por "arrastre tipo" (at) de 0.01168 mn², área resultante de considerar la longitud de la relinga superior de la red del "ANCON" (20,6 m) y las condiciones de operación de pesca descritas anteriormente.
Definición de las asociaciones
La estrategia adoptada para establecer tanto las asociaciones de peces como los subsistemas o habitats (agrupamiento de estaciones de muestreo) que presentaron estructuras faunísticas comunes, fue la de utilizar como técnica básica el análisis multivariado de clasificación jerárquica aglomerativa, mediante el índice cuantitativo de similaridad de Bray-Curtis y el algoritmo de ligamiento promedio no ponderado (Clarke y Ainsworth, 1993; Clarke y Warwick, 1994; Wolff, 1996). Este método se empleó en conjunción con la técnica de ordenación conocida como "non-metrical multidimensional scaling" (NMDS), a fin de efectuar una validación de los resultados (Field et al., 1982; Potter et al., 1997). La aplicación de estas técnicas se hizo con la ayuda del programa PRIMER (Plymouth Routines in Multivariate Ecological Research) (Carr, 1996).
Aunque en el presente trabajo el análisis de la recurrencia espacio-temporal de las asociaciones de peces demersales es efectuada a nivel de todo el área de estudio, vale decir, no hubo estratificación "a priori" del área de estudio, para visualizar y analizar el grado de coherencia espacial de los agrupamientos de estaciones en cada período de muestreo, se tomaron como referencia dos sistemas de sectorización geográfica del área de estudio: el primero delimitado por líneas perpendiculares a la costa (Figura 1) y el segundo por la profundidad. En el primer caso, se consideraron los siguientes sectores de referencia, establecidos a partir de la sectorización propuesta por Márquez (1990):
GN = Guajira Norte (Puerto Estrella - Cabo de la Vela)
GC = Guajira Central (Cabo de la Vela - Camarones)
GS = Guajira Sur (Camarones - Río Piedras)
GdS = Golfo de Salamanca (Santa Marta - Bocas de Ceniza)
SBC = Suroccidente de Bocas de Ceniza (Bocas de Ceniza - Islas del Rosario)
En el segundo caso, se consideró una variante de la propuesta de Bianchi (1992c), quién para diferentes regiones tropicales encontró que los ensamblajes de peces demersales guardaban relación con la siguiente estratificación: plataforma somera (<30 m), plataforma intermedia (30-50 m) y plataforma profunda (>50 m). Esta misma investigadora reconoce que en diferentes sectores de la costa norte de Suramérica, el borde de la plataforma se ubica alrededor de los 100 m de profundidad, situación que también se presenta en el área de estudio (Manjarrés et al., 1996a). Por ello, el cambio consistió en introducir un cuarto sector batimétrico, denominado talud superior, para las estaciones realizadas a más de 100 m de profundidad, tal como hizo Wolff (1996) en la costa pacífica de Costa Rica. De esta forma, para identificar el comportamiento de los agrupamientos con relación a la profundidad, se tuvo la siguiente zonación de referencia:
plataforma somera................10-30 m
plataforma intermedia............31-50 m
plataforma profunda..............51-100 m
talud superior......................101-160 m
Antes de la construcción de las matrices de similaridad, las matrices iniciales fueron sometidas a diferentes operaciones, dependiendo del tipo de análisis, así:
- En el caso del análisis normal (asociaciones de estaciones), se aplicó la transformación logarítmica (ln(x+1)) para balancear la contribución de especies raras y comunes en las representaciones multivariadas no paramétricas, vale decir, remover cualquier efecto dominante producido por capturas extremas o anómalas, tal como lo recomienda Clarke y Warwick (1994) y lo han aplicado Field et al. (1982); Bianchi (1992a,b,c) y Salaverría (1996).
- En el caso del análisis inverso (asociaciones de especies), en lugar de la transformación de las matrices iniciales, se procedió a establecer los porcentajes de cada especie con respecto al total de cada fila. Al trabajar con datos "relativizados" se evita que especies perfectamente correlacionadas, en el sentido de que sus conteos están en estricta proporción a través de las muestras, pudieran ser separadas en los análisis multivariados no paramétricos, por presentar abundancias diferentes (Field et al., 1982).
Análisis del patrón de distribución espacial de las asociaciones de especies
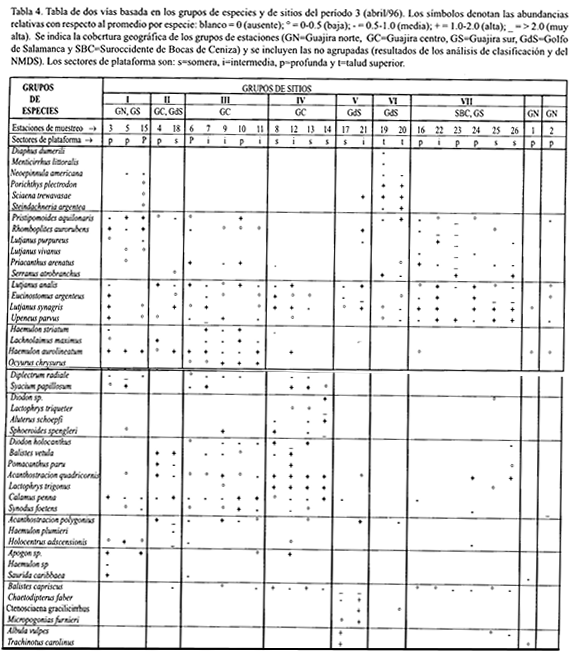
Para analizar la coherencia de las asociaciones de especies con algún patrón de distribución espacial de las estaciones, a partir de los dendrogramas de estaciones y de especies, se construyeron tablas de coincidencia de dos vías u ordenación de dos vías para cada período, vale decir, matrices de datos biológicos con las filas (especies) y las columnas (estaciones) agrupadas según los resultados de los dendrogramas correspondientes (Mahon y Smith, 1989).
Relación con las condiciones ambientales
Se consideraron las variables temperatura y salinidad del agua en el fondo, profundidad, tipo de fondo, latitud, longitud y fecha (ubicación relativa en los 365 días del año). Siguiendo la metodología de Clarke y Ainsworth (1993), antes del cálculo de la matriz de distancias euclidianas, se elaboraron diagramas de dispersión de puntos ("Draftsman Plots") de las variables ambientales, para evaluar el pre-requisito de distribución normal multivariada. Los resultados de los "draftsman plots" indicaron la conveniencia de realizar transformaciones logarítmicas con las variables profundidad y distancia a la costa, y logarítmica inversa con la salinidad. El alto grado de colinearidad (r=-97.3% ; p<0.001) que se presentó entre las variables latitud y longitud llevó a excluir ésta última variable del análisis de correlación. Después, los datos fueron normalizados, de modo que los ejes de las ordenaciones tuviesen escalas comparables (adimensionales).
Para examinar en que medida los patrones abióticos se relacionaban con el patrón biológico observado, se empleó la rutina BIO-ENV del Programa PRIMER (Clarke y Ainsworth, 1993; Clarke y Warwick, 1994; Potter et al., 1997). Mediante este método, para cada período de muestreo en particular, se pudo establecer cuál fue la combinación de variables que conformó el patrón abiótico de mayor ajuste a la ordenación biótica, lo que se determinó mediante el coeficiente de correlación de rangos de Spearman (no paramétrico) entre las dos ordenaciones. La ordenación de los datos bióticos se hizo con base en similaridades de Bray-Curtis de las abundancias transformadas logarítmicamente (Clarke y Ainsworth, 1993).
RESULTADOS
Para ilustrar los cambios temporales que se operan en los diferentes aspectos abordados en este trabajo, se utilizan como punto de referencia los resultados del análisis espacial del período 3 (abril/96), caracterizado por la vigencia de un afloramiento relativamente fuerte.
Agrupamiento de estaciones
En la Figura 2 se presenta el dendrograma de clasificación jerárquica de las estaciones de muestreo del período 3. En este período se identificaron siete agrupamientos de estaciones de muestreo, los cuales mostraron niveles de similaridad relativamente altos (la mayoría por encima del 40%), en comparación con los otros dos períodos. Sólo dos estaciones se ubicaron por fuera de cualquier agrupamiento, como se infiere de la siguiente relación:
Grupo I. Plataforma profunda de los sectores Guajira norte (3), Guajira central (5) y Guajira sur (15).
Grupo II. Plataforma profunda del sector Guajira central (4) y somera del sector Golfo de Salamanca (18).
Grupo III. Plataforma intermedia (7,9,11) y profunda (6,10) del sector Guajira Central.
Grupo IV. Plataforma somera (8,13,14) e intermedia (12) del sector Guajira central.
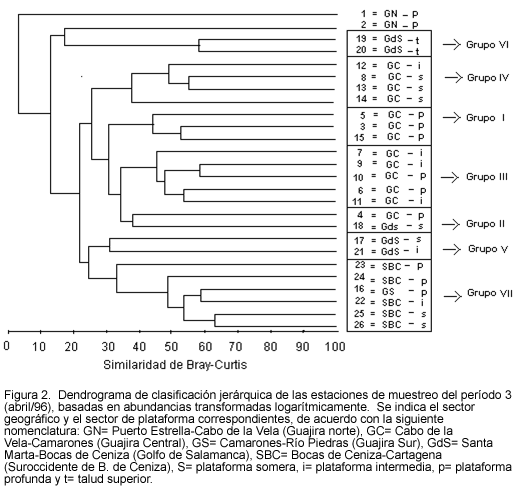
Grupo V. Plataforma somera (17) e intermedia (21) del sector Golfo de Salamanca.
Grupo VI. Talud superior del sector Golfo de Salamanca (19,20).
Grupo VII. Plataforma variada del sector Suroccidente de Bocas de Ceniza (22,23,24,25,26) y profunda del sector Guajira sur (16).
La ordenación global del período 3 (Figura 3) muestra una tendencia de las estaciones de los sectores GC y SBC a agruparse. Otro comportamiento deducible es la delimitación entre la plataforma somera y la intermedia-profunda, y entre ésta y el talud superior (líneas rectas discontinuas). También se destacó la marcada disimilaridad entre las estaciones del GdS: las estaciones del talud superior de este sector mostraron asociaciones notablemente diferentes a las de los otros estratos de plataforma del mismo sector.
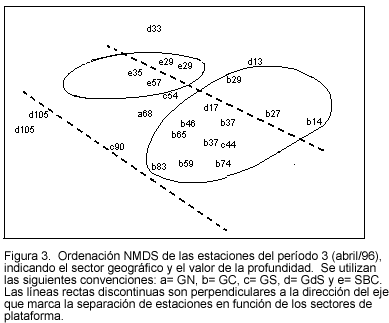
Los grupos de estaciones no fueron estables a través de los diferentes períodos de muestreo. En el aspecto espacial, solamente las estaciones de plataforma intermedia y profunda del sector GC y las de plataforma intermedia del sector SBC mostraron algún grado de coherencia geográfica, aunque en el caso del sector SBC fue también evidente la tendencia a asociarse con estaciones de plataforma intermedia y profunda de la Guajira.
La mayor coherencia geográfica se observó durante el período 3, en el que primaron los agrupamientos de estaciones pertenecientes al mismo sector geográfico (agrupamientos intra-sectores), pero de diferentes sectores de profundidad. Un caso similar, pero en menor proporción, ocurrió en el período 1, en tanto que en el período 2 el predominio correspondió a los agrupamientos de estaciones pertenecientes a sectores no adyacentes, la mayoría pertenecientes al mismo sector de plataforma, lo que sugiere la influencia de la profundidad en el patrón de agrupamiento.
Importancia relativa de las especies
En la Tabla 2 se consignan las 20 especies demersales más importantes del período 3 (afloramiento fuerte), de acuerdo con diferentes índices (%F, A, B e IIR). Nueve especies ocurrieron en más del 50% de las estaciones de muestreo. Los mayores porcentajes de aparición correspondieron a las especies L. synagris, L. analis, U. parvus, E. argenteus y C. penna. En orden de abundancia, las primeras cinco especies colectadas fueron H. striatum, E. argenteus, H. aurolineatum, P. aquilonaris y U. parvus, las cuales reunieron el 55.7% del total de ejemplares colectados en el período. La abundancia de L. synagris triplicó la de L. analis, como se infiere de los respectivos valores de abundancia por arrastre tipo (11.2 ind./at y 3.5 ind./at) y de porcentaje de abundancia (4.5 y 1.4%). En términos de biomasa, dominó L. analis (14.7%), especie que junto con E. argenteus, B. capriscus, L. synagris, C. penna, D. holocanthus (pez globo)y D. guttata (raya) sumaron el 50% de la biomasa de especies demersales colectada. La biomasa por arrastre tipo de L. analis fue significativamente superior a la de L. synagris (7.15 y 3.06 kg/at). De acuerdo con el IIR, las especies más
representativas de este período fueron, en su orden, E. argenteus, L. analis, L. synagris, B. capriscus y C. penna.
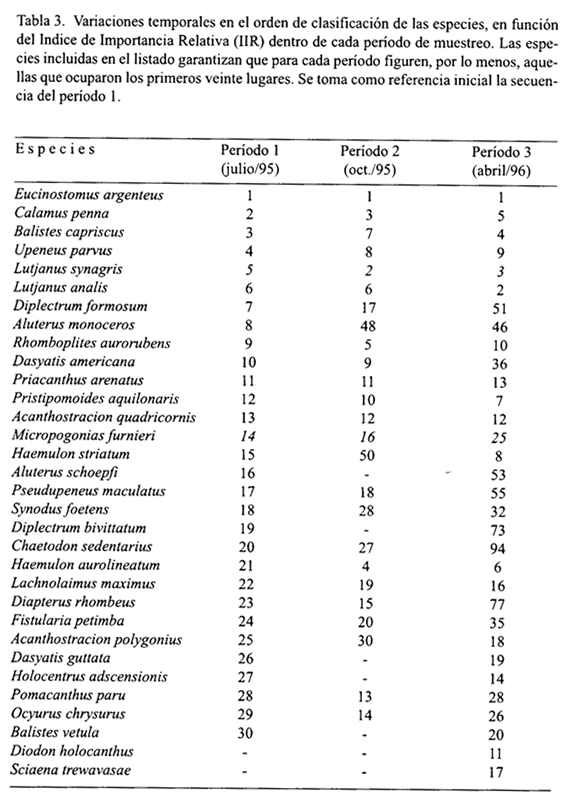
En cuanto al comportamiento temporal, los resultados del IIR evidenciaron cambios en la composición de la ictiofauna demersal a lo largo de los tres períodos de muestreo (Tabla 3). Sólo nueve de las 20 especies que dominaron en las capturas de cada período de muestreo conservaron esta dominancia a lo largo del estudio (E. argenteus, C. penna, B. capriscus, U. parvus, L. synagris, L. analis, P. arenatus, P. aquilonaris y A. quadricornis). Sin embargo, se pueden identificar dos características comunes a los tres períodos: 1) la condición de especie más representativa de E. argenteus, y 2) la ubicación de C. penna y L. synagris dentro de los primeros cinco lugares de importancia.
Asociaciones de especies
El dendrograma de la Figura 4 muestra las asociaciones de especies demersales establecidas para el período 3, las cuales se describen a continuación, teniendo en cuenta la relación entre asociaciones de especies y grupos de estaciones, la cual se planteó con la ayuda de una tabla de dos vías (Tabla 4).
Diaphus dumerili/ Menticirrhus littoralis/ Neoepinnula americana/ Porichthys plectrodon/ Sciaena trewavasae/Steindachneria argentea. Esta asociación se basó en su coincidencia en estaciones del talud superior del sector GdS (Tabla 4), hasta el punto de ser exclusiva del Grupo VI , aún cuando no presentó una constancia alta. De las seis especies que conformaron esta asociación, las dos primeras se agruparon directamente (100% de similaridad), como resultado de su única aparición en una estación del sector GdS (abundancia media). Las restantes cuatro especies se unieron de manera indirecta a las anteriores (45% de similaridad), y basaron su agrupamiento en su coincidencia en las dos estaciones del Grupo VI y, con menores abundancias, en una estación profunda del sector GS (Grupo I).
Pristipomoides aquilonaris/Rhomboplites aurorubens/Lutjanus purpureus/Lutjanus vivanus/Priacanthus arenatus/Serranus atrobranchus.Esta asociación, establecida a un nivel de similaridad del 45%, se presentó en el sector SBC, particularmente en

la plataforma profunda. Excluyendo a S. atrobranchus, también caracterizó las estaciones profundas del sector GN, y excluyendo adicionalmente a P. arenatus, a otra estación profunda del sector GS (Grupo I).
Como se observa, esta asociación incluyó cuatro especies de lutjánidos, cuyas mayores abundancias ocurrieron en la plataforma profunda y, en el caso de P. aquilonaris, en el talud superior. La primera dicotomía de este grupo muestra que P. aquilonaris se asoció de manera directa con R. aurorubens, en tanto que L. purpureus hizo lo propio con L. vivanus. Estas dos últimas especies de pargos mostraron una distribución más restringida que la primera, ya que sólo coincidieron en los sectores GS y SBC; P. aquilonaris y R. aurorubens lo hicieron en los sectores GN, GC, GS y SBC, siendo más abundantes que L. purpureus y L. vivanus en la plataforma profunda.
Lutjanus analis/Eucinostomus argenteus/Lutjanus synagris/Upeneus parvus. Esta fue la asociación de más amplia distribución en todo el área, aunque tuvo mayor constancia y abundancia en el sector SBC (Grupo VII), especialmente en estaciones de plataforma somera e intermedia. Esta asociación también caracterizó diversos subsectores de la Guajira (Grupos I , III y IV).
Haemulon striatum/Lachnolaimus maximus/Haemulon aurolineatum/Ocyurus chrysurus/ Diplectrum radiale/Syacium papillosum. Esta asociación se encontró exclusivamente en el sector GC (Grupo III), con mayores abundancias en estaciones de plataforma intermedia y, en menor grado, profunda. Exceptuando al único lutjánido de esta asociación -O. chrysurus-, este grupo de especies también coincidió en una estación profunda del sector GN (Grupo I). De las especies de esta asociación, O. chrysurus fue la que registró la distribución más localizada.
Diodon sp/Lactophrys triqueter/Aluterus schoepfi/Sphoeroides spengleri. Como la anterior asociación, ésta se presentó exclusivamente en el sector GC, específicamente en el Grupo IV, conformado por estaciones de plataforma somera e intermedia.
Diodon holocanthus/Balistes vetula/Pomacanthus paru/Calamus penna/Acanthostracion quadricornis/Lactophrys trigonus/Synodus foetens. Esta asociación fue exclusiva del sector GC, específicamente de un grupo de estaciones de plataforma somera e intermedia (Grupo IV). No obstante, en la tabla de dos vías (Tabla 4) se aprecia que si se exceptúa a Pomacanthus paru, resulta unaasociación que también estuvo representada en otro Grupo del sector GC (III).
Acanthostracion polygonius/Haemulon plumieri/Holocentrus adscensionis.Este grupo de especies sólo coincidió en una estación somera del sector GdS (Grupo II). De las tres especies que lo conformaron, la distribución más localizada correspondió a H. plumieri (sólo en el sector GdS). A. polygonius se encontró además en el sector GC, y H. adscensionis en todos los sectores del área de estudio.
Apogon sp/Haemulon sp/Saurida caribbaea. Estas especies sólo coincidieron en una estación profunda del sector GN. Esta asociación y la de L. analis/E. argenteus/L. synagris/U. parvus fueron las que caracterizaron el Grupo I (plataforma profunda de GN y GS), aunque esta última no fue exclusiva del Grupo. Apogon sp se capturó también en la plataforma intermedia de GC, y S. caribbaea en la plataforma profunda de GC.
Balistes capriscus/Chaetodipterus faber/Ctenosciaena gracilicirrhus/Micropogonias furnieri.La coincidencia de estas especies se dió únicamente en una estación de plataforma intermedia del sector GdS (Grupo V).
Albula vulpes/Trachinotus carolinus.El agrupamiento de estas especies se debió a su coincidencia en una de las dos estaciones someras del sector GdS (17). A. vulpes también se encontró en dos estaciones someras del sector SBC, mientras que T. carolinus lo hizo en una profunda del sector GN.
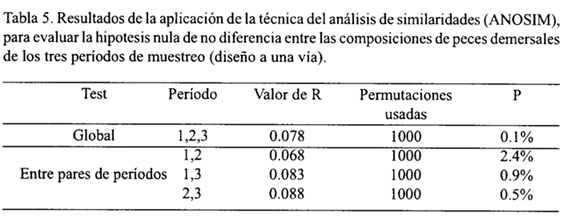
En lo relacionado con las diferencias estacionales en la composición de las asociaciones de especies, la técnica del ANOSIM (test global) demostró que hubo diferencias significativas entre los tres períodos de muestreo. Dado que el valor del estadístico R (0.078) no estuvo comprendido en la distribución simulada de los posibles valores de R, obtenidos mediante el re-rotulado aleatorio de las 77 muestras, hubo suficiente evidencia para rechazar la hipótesis nula "no hay diferencia entre períodos", a un nivel de significancia de P < 0.1% (P < 0.001), vale decir, se presentaron diferencias entre períodos en la estructura de la ictiofauna demersal (Tabla 5).
Para establecer entre qué par específico de períodos existió diferencia significativa, el procedimiento se repitió comparando las similaridades respectivas. En todos los casos, el análisis condujo a rechazar la hipótesis nula, a diferentes niveles de significancia. Por ello, la característica dominante fue la carencia de persistencia temporal de las asociaciones. Unicamente fue factible establecer la persistencia de algunas asociaciones parciales durante un máximo de dos períodos de muestreo. Tal es el caso de R. aurorubens y P. arenatus, especies que conformaron una asociación en el período de afloramiento débil (1), y en el período de afloramiento fuerte (3) se asociaron con P. aquilonaris, L. purpureus, L. vivanus y S. atrobranchus.
Otra asociación que persistió durante dos períodos fue la de L. analis y L. synagris. Durante el período de afloramiento fuerte estuvo asociada con E. argenteus y U. parvus, mientras que en el período de afloramiento débil se les agregó B. capriscus, y se perdió la asociación con E. argenteus y U. parvus. Precisamente la asociación B. capriscus/L. synagris constituyó otro caso de asociación vigente durante dos períodos (1 y 2). Aunque en el período 3 B. capriscus no se agrupó con L. synagris y L. analis, la tabla de dos vías (Tabla 4) muestra que en la gran mayoría de las estaciones donde se capturó B. capriscus también se presentaron las dos especies de lutjánidos mencionadas. Se atribuye el no agrupamiento a que L. synagris y L. analis están más ampliamente distribuidos. Otra asociación común a dos períodos fue la conformada por E. argenteus y U. parvus, la cual, además de presentarse en el período 2 -asociada con H. aurolineatum/P. aquilonaris/C. penna- , se encontró en el período 3, conjuntamente con L. analis y L. synagris.
Relación entre la biota y las condiciones ambientales
La Tabla 6 muestra las combinaciones de variables que dieron lugar a las mayores correlaciones de rango para el período 3, utilizando tanto el coeficiente estándar de Spearman (rs), como la versión ponderada del mismo, conocido como coeficiente armónico de rangos (rw). El tipo de coeficiente de correlación utilizado (rs o rw) no influyó en el orden de prioridad de las variables que "explicaron" de mejor manera el patrón biótico. Además, se presentó una estructura jerárquica en los dos niveles de variables, ya que la variable abiótica que mejor agrupó las estaciones de muestreo fue la profundidad (rw=0.369 y rs=0.462) y la mejor combinación de dos variables retuvo la profundidad y agregó la latitud (rw=0.447 y rs=0.516). Esta combinación significó el óptimo global, ya que la adición de una tercera variable disminuyó el valor de la correlación.
Aún cuando se presentaron cambios estacionales en el conjunto de variables ambientales que explicaron de mejor forma la estructura comunitaria, la variable profundidad siempre figuró en la combinación que produjo la correlación óptima global, en el procedimiento del BIO-ENV. En el período de afloramiento débil estuvo acompañada por la temperatura, la distancia a la costa y la latitud; durante el período de ausencia de afloramiento, la correlación óptima global se alcanzó agregando la variable distancia a la costa; y durante el período de afloramiento fuerte añadiendo la variable latitud.

BIOENV mostró que los valores de disimilaridad bióticos no estuvieron fuertemente correlacionados con las variables ambientales en ninguno de los tres períodos de muestreo, considerando tanto los valores del coeficiente estándar de Spearman (rs) como la versión ponderada del mismo (coeficiente armónico de rangos rw) (Clarke y Ainsworth, 1993; Potter et al., 1997). La mayor correlación se presentó en el período de afloramiento fuerte (rs=0.447; rw= 0.516), mientras que la menor se registró en el período de ausencia del afloramiento (rs=0.341; rw= 0.427).
DISCUSIÓN
Importancia relativa de las especies
Respecto a las especies comerciales de mayor abundancia, la descripción de la ictiofauna demersal hecha en el presente estudio confirma en gran medida los resultados de evaluaciones previas, aunque varía su importancia relativa en términos de biomasa. En general, las especies dominantes en biomasa para todo el área de estudio fueron las mismas que mencionan Duarte y von Schiller (1997) para el área del Golfo de Salamanca (B. capriscus, L. analis, L. synagris y C. penna). También se coincide en la dominancia en número de E. argenteus, B. capriscus y C. penna. Los índices de abundancia de L. synagris y L. analis son inferiores a los reportados por el IMR (1989). En contraste, especies que no figuran entre las comerciales, como B. capriscus, muestran una importancia relativa mayor a la registrada en las primeras evaluaciones de recursos demersales efectuadas en el Caribe colombiano, entre las cuales se cuentan las del B/I Chocó en el Proyecto PNUD-FAO-INDERENA (Ben-Tuvia y Ríos, 1969).
Existen antecedentes sobre descensos en las capturas de las especies explotadas e incrementos de otras no comerciales, como consecuencia de la actividad extractiva (Longhurst y Pauly, 1979; Fariña et al., 1997). Por tanto, es de suponer que en algunos sectores del área de estudio esta actividad, y particularmente la ejercida por los arrastreros camaroneros que iniciaron la pesquería industrial camaronera en 1968 (Mora, 1986; Zamarro, 1997; Weidner y Serrano, 1997), haya afectado la densidad y la estructura de las poblaciones de las principales especies comerciales (pargos), en mayor medida que a las especies no comerciales de talla pequeña, las cuales constituyen las capturas descartadas a bordo de los arrastreros. Estas especies han tenido un aumento en su abundancia debido a la disminución de la competencia y de la depredación por otras especies explotadas (Arancibia, 1992).
Persistencia temporal y variabilidad espacial de las asociaciones de peces demersales
De acuerdo con la escala espacial de los sectores geográficos y la escala temporal del presente estudio, resulta evidente que hay un bajo grado de persistencia temporal de los patrones bióticos observados. Esta característica es consistente con otros estudios de ensamblajes demersales tropicales. Bianchi (1992c) afirma que a pesar de las condiciones estables que priman en las regiones tropicales, grandes áreas de plataformas tropicales están sujetas a importantes fluctuaciones estacionales de diferente tipo. En el área de estudio, estas fluctuaciones están determinadas por la alternación cíclica de los vientos Alisios del noreste (por tanto de la surgencia) y la descarga de aguas continentales (Márquez, 1990; CORPES, 1992). De hecho, los resultados indican un fuerte condicionamiento de los grupos faunales dominantes a la vigencia o ausencia de la surgencia estacional.
Desde el punto de vista batimétrico, las discontinuidades en los grupos faunales fueron más marcadas, observándose una tendencia a la conformación de tres grandes ensamblajes: los de plataforma somera, los de plataforma intermedia y profunda y los del talud superior. Sin embargo, aún cuando fueron más frecuentes los agrupamientos de estaciones intermedias con las profundas, aquellas también se agruparon con algunas someras, lo que ocasionalmente le imprime a esta franja el carácter de zona de transición, como la denominaron Duarte y von Schiller (1997) para el caso del Golfo de Salamanca. Tanto desde el punto de vista de las asociaciones como de las especies en particular, la mayor discontinuidad se presentó entre la ictiofauna de la plataforma continental y la del talud superior.
En general, el esquema dominante fue el de superposición de especies entre los sectores, lo que creó un panorama de cambios graduales en la estructura de los ensamblajes de un sector a otro. Según Mahon y Smith (1989) este patrón es esperado cuando las especies se distribuyen de manera mutuamente independiente, en respuesta a gradientes ambientales, más que cuando los ensamblajes consisten de especies interdependientes, altamente co-evolucionadas.
La significación de las correlaciones entre los patrones bióticos y abióticos indican que las especies asociadas estarían respondiendo de la misma manera al ambiente físico (Mahon y Smith, 1989). No obstante, los resultados del presente estudio indican que si bien se presentaron correlaciones entre el patrón biótico y los factores ambientales, éstas no fueron lo suficientemente fuertes como para limitar las causas de la estructura comunitaria a los gradientes ambientales. Por otro lado, el hecho de que los trabajos existentes sobre hábitos alimentarios de algunas de las especies asociadas indiquen más una relación de competencia que de depredador-presa también contribuye a darle mayor confiabilidad al enfoque ambiental. Una muestra de ello la constituye la relativa persistencia de las asociaciones L. synagris/L. analis, P. arenatus/R. aurorubens y E. argenteus/U. parvus. Sin embargo, existen otros eventos biológicos que pueden influir en los patrones bióticos observados (migraciones de desove, por ejemplo).
Composición y distribución de las asociaciones de especies demersales
En otras regiones del Caribe se han registrado asociaciones similares a las establecidas estacionalmente en este estudio, aunque también se han encontrado algunas diferencias en la conformación y distribución batimétrica de estas asociaciones.
En lo que respecta a las asociaciones de habitats costeros (plataforma somera), se encuentra una coincidencia con los resultados de Bianchi (1992c) en el área de Guayana y Surinam. No obstante la cobertura incompleta de las aguas someras, se presentaron especies exclusivas de este estrato: los haemúlidos Anisotremus virginicus, Conodon nobilis y H. plumieri; losgerréidos Gerres cinereus y Diapterus auratus; loscarángidos Alectis ciliaris y Trachinotus goodei, elsinodóntido Trachinocephalus myops; elbótido Bothus maculiferus; elacantúrido Acanthurus bahianus; eldasiátido Himantura schmardae; y eltetradóndito Sphoeroides dorsalis.
Las asociaciones de plataforma intermedia correspondieron en gran medida a lo que Lowe-McConnell (1962) denominó peces "dorados" y "plateados". A estos grupos corresponden las siguientes asociaciones: H. striatum/H. aurolineatum/D. radiale y B. capriscus/C. faber/C. gracilicirrhus/M. furnieri (período 3); Peristedion sp/R. bicaudalis (período 1); A. rhomboidalis/D. rhombeus (período 2).
Además de los peces "plateados" y "dorados", en la plataforma intermedia también se dieron varias asociaciones de peces pertenecientes a la "zona roja", término que caracteriza los ensamblajes de la plataforma profunda (Lowe-McConnell, 1962). Es el caso de las asociaciones P. arenatus/R. aurorubens (período 1) y P. arenatus/R. aurorubens/L. purpureus (período 3).
En el presente estudio, Rhomboplites y Pristipomoides formaron parte de una asociación de plataforma profunda durante la temporada de afloramiento fuerte (P. aquilonaris/R. aurorubens/L. purpureus/L. vivanus/P. arenatus/S. atrobranchus), encontrada en los sectores Guajira norte, Guajira sur y Suroccidente de Bocas de Cenizas. Se encuentran notables coincidencias entre estos ensamblajes y lo que Bianchi (1992c) registró para la plataforma profunda de Surinam y Guyana, a los cuales denominó ensamblajes de fondos duros: R. aurorubens/P. macrophthalmus/P. arenatus/L. purpureus. Todos los lutjánidos de estas asociaciones fueron referenciados por Acero y Garzón (1985) como especies de habitats rocosos y/o arenosos, en zonas relativamente profundas, debajo de los 30-50 m.
Variables ambientales de mayor relación con el patron biótico
El hecho de que en las tres temporadas la profundidad haya figurado en el arreglo abiótico que explicó de mejor forma el patrón de asociación de los peces demersales representa un resultado similar a los de la mayoría de los estudios que han enfocado este aspecto (Mahon y Smith, 1989 ; IMR, 1989; Arancibia, 1992; Bianchi, 1992a,c ; Blaber et al., 1994; Gray y Otway, 1994; Duarte y von Schiller, 1997; Fariña et al., 1997). Como lo advierten Potter et al. (1997), esto debe interpretarse en el sentido de que el patrón biótico estuvo mediblemente asociado (pero no necesariamente causalmente asociado) con la profundidad. En efecto, Longhurst y Pauly (1987) advierten que la profundidad viene a significar un indicador de la masa de agua presente sobre el fondo.
En su análisis de las estaciones de arrastre de fondo efectuadas entre Surinam y Colombia, el IMR (1989) encontró que los principales cambios ocurren especialmente con la profundidad, mientras que no detectó diferencias zoogeográficas reales entre las diferentes regiones (utilizando la longitud). Esto último no puede afirmarse en el caso del presente estudio, por cuanto en las dos temporadas influidas en mayor o menor grado por el afloramiento (períodos 1 y 3) la latitud figuró en el patrón abiótico de mejor ajuste al biótico. No debe perderse de vista que en el área de estudio tanto la latitud como su variable fuertemente correlacionada, la longitud (R>95%), determinan prácticamente las mismas posibilidades de zonación geográfica a lo largo de la costa. La influencia de la latitud, asociada con otros variables también registra antecedentes en estudios efectuados en regiones tropicales. Al respecto, cabe mencionar el trabajo de Bianchi (1992a) en Angola y el de Martin et al. (1995) en el Golfo de Carpentaria.
Una diferencia notable respecto a resultados presentados por Longhurst y Pauly (1979), Bianchi (1992c) y CORPES (1992), radicó en la ausencia de la variable relacionada con el sedimento dentro de los arreglos abióticos que explicaron de mejor forma el patrón biótico. En estos trabajos se menciona que además de la profundidad, los factores que determinan la subdivisión adicional en sub-comunidades, parecen relacionarse con el tipo de sustrato. En principio, se atribuye este hecho a que los valores de la variable utilizada (% arena) se registraron a partir de cartas sedimentológicas, las cuales sólo permitieron el uso de valores discretos (cuatro categorías de sedimentos). Sin embargo, conviene anotar que Duarte y von Schiller (1997) tampoco encontraron relación entre los patrones de distribución de especies demersales en el Golfo de Salamanca y el tipo de fondo, no obstante haberlo caracterizado mediante unidades phi (diámetro del grano en milímetros).
AGRADECIMIENTOS
Los autores desean expresar su gratitud al Programa de Pesca INPA-VECEP/UE, por el apoyo brindado a las campañas de evaluación de recursos demersales que generaron los datos para este artículo; a los integrantes de Programa, en particular a Inés Patricia Gómez y Jorge Viaña, quienes colaboraron en la edición de algunas gráficas; a las tripulaciones del B/I ANCON (INVEMAR) y del ARC MALPELO (CIOH), a la Dra. Gabriella Bianchi, del IMR (Noruega), por el suministro de varios de sus trabajos; al Dr. Eskild Kirkegaard, del Danish Institute for Fisheries Research- DIFRES-, por el envío de varios trabajos solicitados; y a Mrs. Linda Horsfield, de Plymouth Marine Laboratory (UK), quien aportó la última versión del Programa PRIMER.
BIBLIOGRAFÍA
1 Acero, A. y J. Garzón. 1985. Los pargos (Pisces: Perciformes: Lutjanidae) del Caribe colombiano. Actual. Biol., 14 (53): 89-99. [ Links ]
2 Allen, G.R. 1985. FAO Species Catalogue. Snappers of the world. FAO Fish. Synop., 125 (6), 208 p. [ Links ]
3 Andrade, C., F. Arias e I. Thomas. 1988. Estudio temporal de los sólidos en suspensión entre la desembocadura del río Magdalena y el Canal del Dique, Mar Caribe, Colombia. Mem. VI Sem. Nal. Cienc. Tecn. Mar, CCO, Bogotá: 387-397. [ Links ]
4 Ansari, Z.A., B.S. Chatterti, A. Ingole, R.A. Sreepada, C.U. Rivankar y A.H. Parvlekar. 1995. Community structure and seasonal variation of an inshore demersal fish community at Goa, West coast of India. Est. Coast. Shelf Sci., 41(5): 593-610. [ Links ]
5 Arancibia, H. 1992. Distribution patterns of the demersal fish assemblage off Central Chile. Biol. Pesq., 21: 43-53. [ Links ]
6 Bartels, C.E., W.A. Bussing, M.I. López y K.S. Price. 1983. Ocurrence, distribution, abundance and diversity of fishes in the Gulf of Nicoya, Costa Rica. Rev. Biol. Trop., 31 (1): 75-101. [ Links ]
7 Ben-Tuvia, A. y C. E. Ríos. 1969. Investigación sobre peces y pesca exploratoria del B/I CHOCO a lo largo del Caribe Colombiano durante 1969. Proyecto para el Desarrollo Pesca Marítima en Colombia (PNUD-FAO- INDERENA). Div. Pesq., 1( II)., INDERENA, Bogotá, 47 p. [ Links ]
8 Bianchi, G. 1991. Demersal assemblages of the continental shelf and slope edge between the Gulf of Tehuantepec (Mexico) and the Gulf of Papagayo (Costa Rica). Mar. Ecol. Prog. Ser., 73: 121-140. [ Links ]
9 __________. 1992a. Demersal assemblages of the continental shelf and upper slope of Angola. Mar. Ecol. Prog. Ser. , 81: 101-120. [ Links ]
10 __________. 1992b. Study of the demersal assemblages of the of the continental shelf and upper slope off Congo and Gabon, based on the trawl surveys of the RV "Dr. Fritdjof Nansen". Mar. Ecol. Prog. Ser., 85: 9-23. [ Links ]
11 __________. 1992c. Demersal assemblages of tropical continental shelves. Thesis Dr. Sciant. Univ. Bergen, Noruega, 217 p. [ Links ]
12 Blaber, S.J., D.T. Brewer y A.N. Harris. 1994. Distribution, biomass and community structure of demersal fishes of the Gulf of Carpentaria. Australia. Aust. J. Mar. Freshw. Res., 45(3): 429-443. [ Links ]
13 Blanco, J.A. 1988. Las variaciones ambientales estacionales en las aguas costeras y su importancia para la pesca en la región de Santa Marta, Caribe Colombiano. Tesis M.Sc., Univ. Nal. Col., Bogotá, 50 p. [ Links ]
14 Caddy, J.E. y G.D. Sharp. 1988. Un marco ecológico para la investigación pesquera. FAO. Roma. Doc. Téc. Pesca, 283, 155 p. [ Links ]
15 Carr, M.R. 1996. Primer User Manual. (Plymouth Routines in multivariate ecological research). Plym. Mar. Lab, 41 p. [ Links ]
16 Cervigón, F. 1991. Los peces marinos de Venezuela. Vol. I. 2a. Ed. Los Roques. Caracas, 425 p. [ Links ]
17 __________. 1993. Los peces marinos de Venezuela. Vol. II. 2a. Ed. Los Roques. Caracas, 497 p. [ Links ]
18 __________. 1994. Los peces marinos de Venezuela. Vol. III. 2a. Ed. Los Roques. Caracas, 295 p. [ Links ]
19 __________. 1996. Los peces marinos de Venezuela. Vol. IV. 2a. Ed. Los Roques. Caracas, 254 p. [ Links ]
20 Clarke, K.R. y M. Ainsworth. 1993. A method of linking multivariate community structure to environmental variables. Mar. Ecol. Prog. Ser., 92: 205-219. [ Links ]
21 __________, y R.M. Warwick. 1994. Change in marine communities: an approach to statistical analysis and interpretation. Natural Environment Research Council, UK, 144 p. [ Links ]
22 Collete, B.B. y C.E. Nauen. 1983. FAO Species Catalogue. Scombrids of the world. FAO Fish. Synop., 125 (2), 137 p. [ Links ]
23 Compagno, L.J.V. 1984. FAO Species Catalogue. Vol. 4. Sharks of the world. Part 2. FAO Fish. Synop., 125 (4): 251-655. [ Links ]
24 Corpes (Consejo Regional de Planificacion de la costa Atlantica). 1992. El Caribe colombiano. Realidad Ambiental y Desarrollo. Cons. Reg. Planif. Costa Atlánt., Santa Marta, 275 p. [ Links ]
25 Donoso, M.C. 1992. Circulación de las aguas del mar Caribe. Mem. VII Sem. Nal. Ciencias Tecn. Mar., CCO, Bogotá: 345-356. [ Links ]
26 Doubleday, W.G. (Ed.). 1981. Manual on groundfish surveys in the Northwest Atlantic, NAFO Sci. Coun. Studies, 2, 55 p. [ Links ]
27 Duarte, L. y D. von Schiller. 1997. Comunidad de peces demersales del Golfo de Salamanca (Caribe colombiano): estructura espacio-temporal y caracterización trófica con énfasis en los habitos alimentarios de Lutjanus analis (Cuvier, 1828), Lutjanus synagris (Linnaeus, 1758), Balistes capriscus (Gmelin, 1788) y Balistes vetula (Linnaeus, 1758). Tesis de grado. Univ. Jorge Tadeo Loz., Santa Marta, 215 p. [ Links ]
28 Fariña, A.C., J. Freire y E.González-Gurriáran. 1997. Estructura espacial y cambios a largo plazo de las comunidades megabentónicas de la plataforma continental y talud superior de Galicia (noreste de España): análisis comparativo de peces y crustáceos decápodos. Publ. Espec. Inst. Esp. Oceanog., 23: 53-70. [ Links ]
29 Field, J.G., K.R. Clarke y R.M. Warwick. 1982. A practical strategy for analysing multispecies distribution patterns. Mar. Ecol. Prog. Ser. 8: 37-52. [ Links ]
30 Gray, C.A. y N.M. Otway. 1994. Spatial and temporal differences in assemblages of demersal fishes on the inner continental shelf off Sidney, South Eastern Australia. Aust. J. Mar. Freshwater. Res., 45(4): 665-676. [ Links ]
31 IMR (Institute of Marine Research). 1989. Prospecciones de los recursos pesqueros de las áreas de plataforma entre Surinam y Colombia. NORAD/UNDP/FAO Programe, Bergen, 160 p. [ Links ]
32 Longhurst, A.R. y D. Pauly. 1987. Ecology of tropical oceans. Academic Press, New York, 407 p. [ Links ]
33 Lowe-McConnell, R.H. 1962. The fishes of the British Guyana continental shelf with notes on their natural history. J. Linn. Soc. London Zool. 44 (301): 669-700. [ Links ]
34 Mahon, R y R.W. Smith. 1989. Demersal fish assemblages on the Scottian Shelf, northwest Atlantic: Spatial distribution and persistence. Can J. fish. Aquat. Sci. 46 (suppl.1): 134-152. [ Links ]
35 Manjarres, L., A. Vergara, J. Torres, G. Rodríguez, E. Arteaga, J. Viaña, J. Arévalo, R. Galvis y J. Rodríguez. 1995. Crucero de evaluación de recursos demersales en el Caribe colombiano. Informe Final. Rev. No. RA-006. INPA-VECEP/INVEMAR/ DEMER/9501. Julio 1995. Santa Marta, 72 p. [ Links ]
36 __________, G. Rodríguez, J. Torres, A. Vergara, E. Arteaga, J. Arévalo, R. Galvis, J. Rodríguez y J. Viaña. 1996a. Crucero de evaluación de recursos demersales en el Caribe colombiano. Rev. RA-007. Informe Final. INPA-VECEP/INVEMAR/ DEMER/9502. Octubre-Noviembre de 1995. Santa Marta, 30 p. [ Links ]
37 __________, G. Rodríguez, A. Vergara, E. Arteaga, J. Rodriguez, J. Arévalo, J. Viaña, R. Galvis, J. Torres y M. Barros. 1996b. Crucero de evaluación de recursos demersales en el Caribe colombiano. Rev. RA-007. Informe Final. INPA-VECEP/INVEMAR/ DEMER/9502. Octubre-Noviembre de 1995. Santa Marta, 30 p. [ Links ]
38 Márquez, G.E. 1982. Los sistemas ecológicos marinos del sector adyacente a Santa Marta, Caribe colombiano I: Generalidades. Ecol. Trop., 2(1): 5-18. [ Links ]
39 Márquez, G. 1990. Ecosistemas marinos. Caribe Colombia. Fondo FEN Colombia, Bogotá: 115-133. [ Links ]
40 Martin, T.J., D.T. Brewer y S.J. Blaber. 1995. Factors affecting distribution and abundance of small demersal fishes in the Gulf of Carpentaria. Aust. J. Mar. Freshwater Res., 40(6): 909-920. [ Links ]
41 Mora, J.H. 1986. La industria pesquera en el Caribe colombiano. Memorias del V Seminario Nacional de Ciencias del Mar, Buenaventura. Nov. 24-27/86, 246 p. [ Links ]
43 Potter, I.C., P.N. Claridge, G.A. Hyndes y K.R. Clarke. 1997. Seasonal, annual and regional variations in the compositions of the icthyofauna in the Inner Severn Estuary and Inner Bristol Channel. Plym. Mar. Lab., Plymouth, 36 p. [ Links ]
43 Salavarria, A. 1996. Ecología de las comunidades demersales presentes en Puerto Quetzal durante las prospecciones de marzo y sept. de 1995 y marzo de 1996. CEMA-USAC. Guatemala, 31 p. [ Links ]
44 Vernette, G. 1985. Síntesis del proyecto Bahía de Cartagena. Bol. Cient. CIOH. 4, Cartagena: 49-110. [ Links ]
45 Weidner, D y J. Serrano. 1997. World Swordfish Fisheries. Latin America. Part A. South America. US Departm. Comm., NOAA, Vol. IV. 428 p. [ Links ]
46 Wolff, M. 1996. Demersal fish assemblages along the Pacific coast of Costa Rica: a quantitative and multivariate assessment based on the Victor Hensen Costa Rica Expedition (1993/1994). Rev. Biol. Trop., 44 (Suppl.3): 187-214. [ Links ]
47 Zamarro, J. 1997. Revisión de la investigación pesquera en Colombia. Programa INPA-VECEP. Doc. inéd., 59 p. [ Links ]
FECHA DE RECEPCION 01/07/1999 FECHA DE ACEPTACION 10/05/2001
