










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista U.D.C.A Actualidad & Divulgación Científica
Print version ISSN 0123-4226
rev.udcaactual.divulg.cient. vol.14 no.2 Bogotá Jul./Dec. 2011
CIENCIAS AGROPECUARIAS - Artículo Científico
EVALUACIÓN DEL RENDIMIENTO Y SUS COMPONENTES EN GENOTIPOS DE CEBADA (Hordeum vulgare L.) DIFERENCIADOS POR SU TIPO DE ESPIGA Y GRANO
EVALUATION OF YIELD AND ITS COMPONENTS IN BARLEY (Hordeum vulgare L.) GENOTYPES DIFFERENTIATED FOR SPIKE AND KERNEL TYPES
Diana Katherinne Rios1; Rodrigo Britto2; Hernando Delgado3
1Ingeniero Agrónomo. Grupo Manejo Biológico de Cultivos. GMBC. Facultad de Ciencias Agropecuarias, Universidad Pedagógica y Tecnológica de Colombia UPTC. Av. Central del Norte, Tunja, Colombia. Correo electrónico:dianakatherinne.rios@uptc.edu.co
2Ing. Agrónomo. Fitomejorador Ph.D. Programa Mejoramiento Genético de Cereales Menores, FENALCE, Tunja, Colombia. Correo electrónico:rbritto2@yahoo.com
3Ing. Agrónomo. M.Sc. Fitotecnia: Genética y Fitomejoramiento. Profesor Asistente. Grupo Manejo Biológico de Cultivos. GMBC. Facultad de Ciencias Agropecuarias, Universidad Pedagógica y Tecnológica de Colombia UPTC. Av. Central del Norte. Tunja, Colombia. Correo electrónico:hernando.delgado@uptc.edu.co
Rev. U.D.C.A Act. & Div. Cient. 14(2): 55 - 63, 2011
RESUMEN
Con el objeto de poder determinar en el futuro los cruzamientos más deseables al combinar cuatro tipos de cebada, se estudió en Duitama y en Samacá (Boyacá, Colombia), el efecto diferencial del tipo de espiga y de grano en el rendimiento final y sus componentes, de doce genotipos, en un diseño de parcelas divididas, con cuatro repeticiones. Se efectuó análisis de varianza para cada una de las localidades y características evaluadas, tomando el promedio del grupo de los genotipos involucrados. La comparación de medias de tratamientos, se realizó por el método de Diferencia Mínima Significativa. Los genotipos con espigas de dos carreras produjeron mayor número de macollas efectivas (Samacá) y granos más pesados (Duitama, Samacá), mientras que los de seis carreras tuvieron un mayor número de granos por espiga (Duitama, Samacá). Adicionalmente, en Duitama, los genotipos con granos desnudos produjeron mayor número de granos, pero los de granos cubiertos dieron granos más pesados. La interacción entre el número de carreras y el tipo de grano solo presentó significancia para el número de granos por espiga, en la localidad de Duitama. En cuanto al rendimiento de grano seco en Duitama, ninguna de las fuentes de variación mostró efectos significativos. Respecto al número de carreras, se pudo deber a compensación entre los componentes del rendimiento, algunos mayores y otros menores en uno u otro tipo, entre las cebadas de dos y seis carreras evaluadas.
Palabras clave: Cebada, características, carreras, grano.
SUMMARY
In order to avoid no desired crosses to combine four types of barley, the differential effect of spike type and grain on final yield and its components was studied in twelve genotypes at Duitama and Samacá (Boyacá, Colombia) in a split-plot design with four replications. An analysis of variance was practiced, locally, for each variable taking the mean genotype groups as a treatment and not an individual genotype. Differences among treatments were determined by a Least Significance Difference. Two-rowed barley genotypes gave a higher number of effective tillers (Samacá) and heavier kernels (Duitama, Samacá); while, six-rowed barley genotypes had a greater number of kernels/spike (Duitama, Samacá). Additionally, in Duitama, hull absent kernel barley genotypes produced a greater number of kernels/spike; but, hulled kernel barley genotypes gave heavier kernels. The row number x hull prevalence interaction only affected the kernels/spike number (Duitama). As for the yield of dry grain in Duitama, none of the sources of variation presented significant effects. With regard to the row number, this may be due to compensation between the yield components, some higher than others in one or another type, between the two-rowed and six-rowed barleys evaluated.
Key words: Barley, features, rows, kernel.
INTRODUCCIÓN
La cebada (Hordeum vulgare L.) pertenece al grupo de cereales cultivados desde la antigüedad, 7.000 años a.C. En Colombia, desde su introducción, en 1539, fue uno de los principales cultivos de tierra fría, llegando a ocupar hasta 75.600ha, en 1975, que produjeron 122.000t (Rico, 1980). El área sembrada, en el 2009, fue de 6.372ha, con una producción total de 9.034t, reduciéndose en 27,1%, respecto del 2008, por los efectos del fenómeno del niño. Las importaciones de este mismo año alcanzaron 189.186t (FENALCE, 2009), por lo que se está muy lejos de suplir la demanda, a nivel nacional.
En el país, se siembran los cuatro tipos de cebada, que resultan de la combinación del número de carreras en la espiga y la persistencia de las glumillas del grano: las de dos y seis carreras, cada una con granos desnudos y cubiertos. El objetivo principal del fitomejoramiento en cereales menores en Colombia, ha sido el obtener genotipos con adecuados rendimientos de grano, resistentes a las enfermedades, especialmente roya amarilla, y con una buena calidad. El rendimiento es quizá la parte más importante, ya que determina que se siembre o no un genotipo dado, por parte de los agricultores.
El rendimiento de grano ha sido descrito como el volumen de un paralelepípedo, en el cual, el número de espigas por unidad de superficie, el número de granos por espiga y el peso de los granos, constituyen las aristas. Un incremento en cualquiera de los tres componentes determinará un aumento del rendimiento, siempre y cuando no haya una disminución correspondiente en los otros dos (Barriga, 1974; Ataei, 2006).
Okuyama et al. (2005) encontraron que en trigo, el diámetro del tallo, la longitud de la espiga y la altura de planta, fueron los caracteres más relacionados con el mayor rendimiento de grano. Aproximadamente, el 72% de la variación en el rendimiento de los cereales está relacionada al número de espigas fértiles, número de granos por espiga y el peso del grano. Así mismo, estos componentes tienen una correlación positiva y altamente significativa con el rendimiento en cebada (Ataei, 2006). Según Castañeda-Saucedo et al. (2009), el número de espigas por metro cuadrado es el componente que mayor contribución tiene en el rendimiento en cereales de grano pequeño.
El primer componente del rendimiento de grano es el número de macollas, que causa un incremento de área foliar o fotosintética. El segundo componente es el número de granos por espiga; este componente del rendimiento de grano en cebada identifica la etapa de crecimiento y el estado reproductivo y va a depender del número de flores que son polinizadas y del número de flores polinizadas que se llenan y se convierten en grano (Ataei, 2006). El tercer componente es el peso del grano; diversos genes individuales afectan el peso del grano (Rasmusson, 1985). Se ha demostrado que la translocación de materiales asimilados puede responder por cerca del 30% de peso final del grano (Gallagher et al. 1975).
Eshghi & Akhundova (2009) plantean que la elección de un procedimiento eficaz de mejoramiento depende, en gran medida, del conocimiento del control genético del caracter a seleccionar. En este sentido, estos mismos autores, a través de un análisis dialélico, en variedades de cebada de grano desnudo, resaltaron los efectos genéticos aditivos, para altura de planta, número de macollas y días a madurez, coincidiendo con lo encontrado por Islam & Darrah (2005) y Verma et al. (2007), en genotipos de cebada desnudos y cubiertos; mientras tanto, para el número de granos por espiga, se registraron efectos genéticos de sobredominancia. Dado que la heredabilidad, en sentido estricto del rendimiento de grano por planta fue baja, concluyeron que la mejora directa de éste es, de alguna manera problemática, porque los factores ambientales contribuyen, en gran medida, en su control (Eshghi & Akhundova, 2009). Resultados similares para altura de planta, número de macollas, días a madurez y rendimiento de grano por planta hallaron Singh et al. (1999) y Budak (2000). Entre tanto, Rohman et al. (2006) reportan que ambos componentes, aditivos y no aditivos, fueron importantes en el control de la herencia de estas características, pero los componentes de dominancia fueron más predominantes que los aditivos.
Eshghi et al. (2010) resaltan que efectos genéticos aditivos y de dominancia fueron importantes para la mayoría de los caracteres evaluados, pero dominancia e interacción no alélica tuvieron un efecto más pronunciado para número de granos por espiga en condiciones de sequía y para peso de 1.000 granos y rendimiento de grano en ambos ambientes. La mayor heredabilidad fue para número de granos por espiga en condiciones normales y número de macollas en ambos ambientes, indicando que estos caracteres son controlados por efectos aditivos. Debido a baja ganancia genética y baja heredabilidad para rendimiento de grano comparado con otros caracteres, indicaría que su efecto fenotípico es principalmente controlado por consecuencia ambiental, por lo que sería más recomendable la selección de genotipos superiores con base en componentes del rendimiento, más que por rendimiento de grano.
Varios estudios han identificado loci de caracteres cuantitativos (QTLs) para el rendimiento de grano y sus componentes en cebada. Cakir et al. (2003) reportaron tres QTLs en los cromosomas 2H, 3H y 5H, para rendimiento de grano en cebada. Li et al. (2006) identificaron varios QTLs para este mismo caracter y sus componentes, como el número de granos por espiga en el cromosoma 1H.
Arisnabarreta & Miralles (2006) estudiaron el efecto del N aplicado en el rendimiento de grano y sus componentes en cebadas de dos y seis carreras. En ambos tipos, las variaciones en rendimiento de grano fueron explicadas por cambios en granos por unidad de área más que por diferencias en el promedio del peso de grano. En la cebada de dos carreras, los cambios en el número de granos se debieron a variaciones en el número de espigas por m2, mientras que en la cebada de seis carreras, se debieron al número de granos por espiga. El hecho que la respuesta a nitrógeno mediante el número de granos por unidad de área en las cebadas de dos y seis carreras es mejor explicado por diferentes subcomponentes del rendimiento, lleva a estos autores a especular acerca que el periodo crítico para la determinación del rendimiento podría diferir entre estos dos tipos de cebada.
Por el papel que juegan el rendimiento de grano y sus componentes en la definición de una línea promisoria como variedad comercial, esta investigación tuvo como objetivo principal evaluar dichas características, en doce genotipos de cebada, diferenciados por tipos de espiga y de grano.
MATERIALES Y MÉTODOS
La investigación, se realizó en el segundo semestre de 2009, en los municipios de Duitama (Centro Demostrativo Surbatá - ICA, con coordenadas N 5°47”, W 73°04” y altitud de 2515 m.s.n.m.) y en Samacá (Vereda Tibaquira Alto, N 5°29’, W 73°30’ y altitud de 2665 m.s.n.m.), en el Departamento de Boyacá (Colombia). Se evaluaron doce genotipos de cebada: seis con espigas de seis carreras y los restantes de dos carreras. Cada uno de los grupos tenía tres genotipos con el grano desnudo y tres con el grano cubierto (cuadro 1). En las dos localidades, se utilizó un diseño de parcelas divididas con cuatro repeticiones, siendo la parcela principal, el número de carreras de la espiga y la subparcela, el tipo de grano. Cada unidad experimental constó de cuatro surcos de cinco metros de largo, con una separación de 30cm.
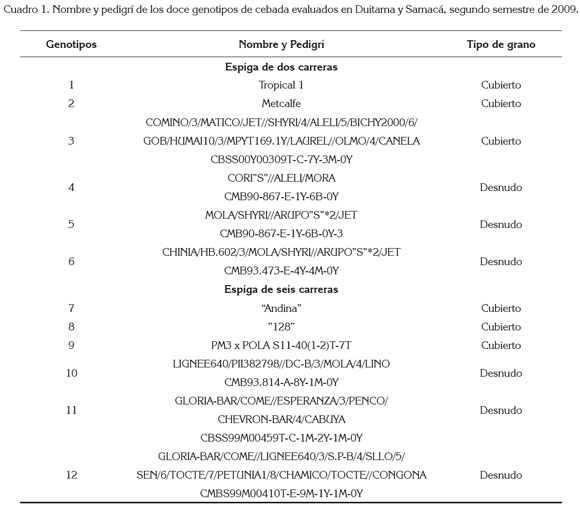
La densidad de siembra, se calibró por el peso de 1.000 granos y el porcentaje de germinación, de cada genotipo, tomando como base el testigo, la variedad "128", con una densidad de 60kg ha-1, de manera que cada uno de los genotipos quedara con igual número de plantas por surco. En los surcos borde, se sembró la selección especial "Tropical", para garantizar la competencia en los surcos iniciales y finales de cada repetición. La siembra se realizó a chorrillo, el 5 de septiembre de 2009, en Samacá y el 14 de septiembre, en Duitama. Las prácticas culturales de campo, se efectuaron de acuerdo con lo establecido por el ICA, para el manejo de pruebas regionales de cereales menores. La cosecha, se procedió con hoz, en seis fechas diferentes, comprendidas entre el 16 de enero y el 11 de febrero de 2010, debido a la presencia de genotipos precoces y tardíos. Cada uno de los manojos cosechados por unidad experimental fueron trillados separadamente; el grano obtenido fue limpiado y, posteriormente, se tomó peso y humedad, en un medidor electrónico marca Motomco Ion. Al finalizar el ciclo del cultivo, se determinaron las siguientes variables en cada unidad experimental:
Número de macollas efectivas por planta: En 20 plantas en competencia de los surcos centrales en cada unidad experimental, entendiendo como macolla efectiva aquella que produce una espiga que tenga como mínimo un grano.
Número de granos por espiga: De cada una de las 20 plantas seleccionadas, previamente, se tomó la espiga principal, se trilló, de forma individual y se contó el número de granos.
Peso del grano: De la producción total de cada unidad experimental, se tomó el peso de 1.000 granos, en dos replicaciones.
Rendimiento de grano: Con base en el peso y la humedad de la producción total de cada unidad experimental, se determinó en g/parcela al 15% de humedad.
Análisis estadístico: Por ser de mayor interés el inferir acerca de las características de la espiga (carreras) y del grano (desnudez o persistencia de las glumillas), que de un genotipo en particular, para el análisis de varianza, se tomó el promedio del grupo de los genotipos involucrados. Así, para las carreras, se trabajó con el promedio de los seis genotipos (cinco para los de seis carreras, por baja germinación del genotipo número 10) y para la desnudez del grano, con el promedio de los tres genotipos (dos para los de seis carreras, grano desnudo, por la misma razón anterior). Para cada variable y sitio, se realizó análisis de varianza, mediante el paquete estadístico SAS v.8e (SAS Institute Inc., Cary, NC) y para comparar las medias de los tratamientos, se aplicó el método de la Diferencia Mínima Significativa, sólo cuando la prueba de F del análisis de varianza indicó diferencias significativas. En todas las pruebas, se utilizó un nivel de significancia de 0,05 y 0,01.
RESULTADOS Y DISCUSIÓN
En las tabla 1 y 2, se registran los cuadrados medios y el nivel de significancia de las características: número de macollas efectivas por planta, número de granos por espiga y peso de 1.000 granos en las dos localidades. En el municipio de Samacá, el rendimiento de grano no pudo ser determinado, debido a baja población de plantas en las parcelas por el intenso verano, que no permitió aplicar el número de riegos adecuado.
Número de macollas efectivas por planta: Los análisis de varianza (tabla 1 y tabla 2), indican que el número de carreras, el tipo de grano y la interacción de número de carreras por tipo de grano no presentaron efecto significativo sobre este componente en Duitama, mientras que en Samacá (tabla 2) sólo mostró efecto significativo el número de carreras. En efecto, en Samacá las cebadas de dos carreras registraron mayor número de macollas con respecto a las de seis, cuya diferencia de 0,44 fue altamente significativa (tabla 3), lo cual, concuerda con lo reportado por Rasmusson (1985) y con García del Moral et al. (2003), en cuanto a que la cebada de seis carreras presentó una tasa de macollamiento reducida, incluso, bajo condiciones favorables de crecimiento, en comparación con la de dos carreras. Asociación positiva entre el número de espigas o macollas efectivas y el rendimiento de grano ha sido encontrada por Castañeda-Saucedo et al. (2009), Rasmusson (1985) y Barriga (1974). Para estos últimos autores es el componente de mayor influencia en el rendimiento.
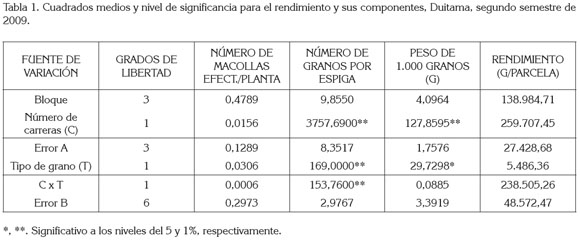
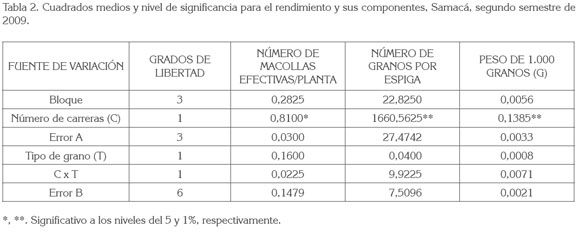
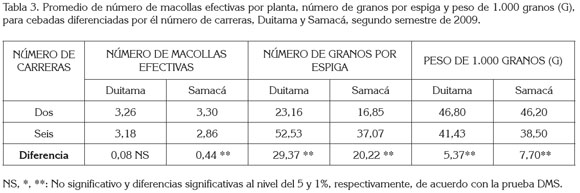
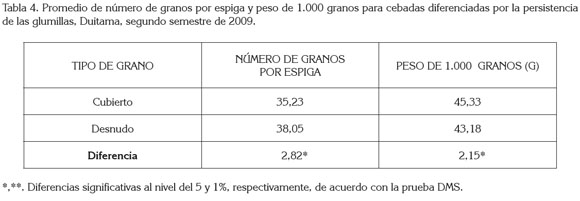
Número de granos por espiga: El número de carreras, el tipo de grano y la interacción entre ellos, mostraron efecto significativo (p<0.001) sobre este componente, en el municipio de Duitama (tabla 1 ). En Samacá, solo mostró efecto significativo (p<0.001) el número de carreras (tabla 2). En efecto, en las dos localidades, las cebadas de seis carreras mostraron un mayor número de granos por espiga que las de dos carreras. Las diferencias en cada localidad de 29,37 y 20,22 fueron altamente significativas (tabla 3); esto coincide con lo reportado por Kjaer & Jensen (1996) y por García del Moral et al. (2003), quienes hallaron resultados similares en evaluación de estos dos tipos de cebadas. Lo anterior es debido a que las cebadas de seis carreras presentan mayor número de espiguillas formadas (tres), en comparación con las de dos carreras, en las que solo se aprecia desarrollada la espiguilla intermedia, mientras abortan las laterales. Las cebadas de grano desnudo en Duitama dieron un mayor número de granos por espiga que las cebadas de grano cubierto; la diferencia de 2,82 granos por espiga fue significativa (tabla 4).
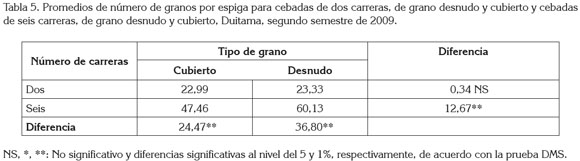
En cuanto a la interacción altamente significativa del número de carreras por el tipo de grano en Duitama (tabla 1), al estudiar las comparaciones pertinentes (tabla 5), se encontró lo siguiente, considerando el Número de Carreras: el número de granos por espiga en las cebadas de dos carreras no mostró diferencias significativas, debido al tipo de grano cubierto o desnudo, pero sí en las cebadas de seis carreras, con diferencias altamente significativas, a favor del grano desnudo. Según Bothmer & Jacobsen (1985), como las cebadas cubiertas dieron origen a las desnudas, la masa final de la espiga estaba programada genéticamente para ser la misma en ambos tipos, pero con la evolución, las de dos carreras emplearon la energía no utilizada en la formación de las glumillas, para formar un mayor número de granos. Mientras tanto, considerando el Tipo de Grano: el número de granos por espiga, tanto en las cebadas de grano cubierto como en las cebadas de grano desnudo fue diferente dependiendo del número de carreras de la espiga; las diferencias altamente significativas favorecieron a las cebadas de seis carreras; esto en la práctica, parece bastante lógico por el mayor número de flores (espiguillas), fecundadas en estas últimas.
Peso de 1.000 granos: El análisis de varianza para este componente de rendimiento (tablas 2 y 3) mostró diferencias altamente significativas para número de carreras en los dos municipios y significativas para el tipo de grano, exclusivamente, en la localidad de Duitama. En efecto, el peso de 1.000 granos fue mayor, en ambos sitios, en las cebadas de dos carreras, cuyas diferencias de 5,37g (Duitama) y 7,7g (Samacá) fueron altamente significativas (tabla 4), lo cual, es explicable por la menor competencia por los asimilados en estas últimas cebadas, al tener menor número de granos. Kjaer & Jensen (1996) encontraron resultados similares al mencionar que en las variedades de dos carreras el peso de grano fue mayor que en las de seis, pero que estas últimas superaron a las primeras, en el número de granos por espiga.
Gallagher et al. (1975) y Rasmusson (1985) plantean que el peso del grano dentro de un grupo determinado de cebadas es un componente de rendimiento de grano bastante estable, porque se produce movilización de los asimilados de reservas del tallo y otros tejidos vegetales, que pueden compensar la carencia de los fotoasimilados corrientes. Por esto, para algunos autores, granos por espiga y espigas por planta tienen un mayor efecto directo en el rendimiento que el peso del grano (Rasmusson, 1985) y, para otros, el efecto de peso del grano en el rendimiento es despreciable o inconsistente (Sinebo, 2002).
Como se expresó anteriormente, la fuente de variación tipo de grano presentó diferencias significativas en Duitama (tabla 4), siendo más pesados los granos de las cebadas cubiertas, cuya diferencia con los granos de cebadas desnudas, 2,15g, fue significativa al nivel del 5%, lo cual, se debe, en parte, a que este tipo de grano presenta el peso adicional de las glumillas adheridas.
Rendimiento de grano: En el análisis de varianza (tabla 1), ninguna de las fuentes de variación: número de carreras de la espiga (C), tipo de grano (T) y la interacción C x T presentó efectos significativos en el rendimiento (g/parcela), en el municipio de Duitama, donde se pudo medir esta característica. En cuanto al número de carreras, se pudo deber a la compensación entre los componentes del rendimiento de grano, algunos mayores y otros menores, en uno u otro tipo, entre las cebadas de dos y seis carreras evaluadas. Estos efectos compensatorios han sido reportados por Mitchell et al. (1996) y Sheehy et al. (2001), quienes plantean que con frecuencia la utilidad de los componentes del rendimiento, como criterio de selección, se puede ver limitada por los efectos de compensación y que se ha constatado que existe una correlación negativa entre los componentes del rendimiento, de manera que no se puede seleccionar positivamente para todos ellos, al mismo tiempo. Lo más conveniente parece ser lograr el equilibrio entre dichos componentes, mejorando aquel que sea más eficiente en un determinado genotipo, mientras los demás, se mantienen constantes. Similarmente, la compensación entre los componentes del rendimiento en cebada ha sido reportada también por García del Moral et al. (1991) y por Conry (1994).
De forma general, se podría concluir que la variable número de carreras por espiga presentó efectos significativos sobre los tres componentes de rendimiento; macollas efectivas por planta (Samacá), número de granos por espiga (Duitama, Samacá) y peso de 1.000 granos (Duitama, Samacá). Es así como los genotipos con espigas de dos carreras produjeron un mayor número de macollas efectivas (Samacá) y granos más pesados (Duitama, Samacá); no obstante, los genotipos con espigas de seis carreras fueron superiores en cuanto al número de granos por espiga (Duitama, Samacá). El tipo de grano solo presentó los efectos más significativos en el número de granos por espiga y peso del grano: los genotipos de cebada con granos desnudos produjeron mayor número de granos por espiga (Duitama); pero los genotipos con granos cubiertos dieron granos más pesados (Duitama).
La interacción entre el número de carreras de la espiga y el tipo de grano, solo afectó con alta significancia el número de granos por espiga en Duitama. Dentro del número de carreras de la espiga, los genotipos de granos desnudos y cubiertos fueron similares en número de granos en las de dos carreras; pero en las de seis carreras fueron superiores con alta significancia, en número de granos, los genotipos de granos desnudos. Y dentro del tipo de grano, los genotipos de seis carreras sobrepasaron con alta significancia en número de granos a los de dos carreras, tanto en los granos desnudos como en los cubiertos.
El presente trabajo permitió resaltar las características propias de los componentes del rendimiento en cada tipo de cebada, según su número de carreras y tipo de grano, lo cual, será de utilidad para la selección en campo y para orientar las futuras hibridaciones.
Conflictos de interés: El manuscrito fue preparado y revisado con la participación de todos los autores, quienes declaramos que no existe ningún conflicto de intereses que ponga en riesgo la validez de los resultados presentados.BIBLIOGRAFÍA
1. ARISNABARRETA, S.; MIRALLES, D.J. 2006. Yield responsiveness in two- and six-rowed barley grown in contrasting nitrogen environments. J. Agron. Crop Sci. 192(3): 178-185. Disponible desde Internet en: http://onlinelibrary.wiley.com/doi/10.1111/j.1439-037X.2006.00203.x/abstract (con acceso el 20/03/11). [ Links ]
2. ATAEI, M. 2006. Path analysis of barley (Hordeum vulgare L.) yield. Tarim Bilimleri Dergisi. Ankara Universitesi Ziraat Fakultesi (Irán). 12(3):227-232. [ Links ]
3. BARRIGA, P. 1974. Análisis de causa y efecto para rendimiento y componentes del rendimiento en trigo de primavera. Agro Sur. 2(1):01-05. Disponible desde Internet en: http://mingaonline. uach.cl/scielo.php?script=sci_arttext&pid=S0304- 88021974000100001&lng=es&nrm=iso> (con acceso el 10/03/10). [ Links ]
4. BOTHMER, R. von; JACOBSEN, N. 1985. Origin, Taxonomy and Related Species. In: Rasmusson, D.C. Ed. Barley. American Society of Agronomists, Madison, Wis. p.19-56. [ Links ]
5. BUDAK, N. 2000. Heterosis, general and specific combining ability estimates at F1 and F2 generations of 8´8 diallel population in barley. Turk. J. Field Crops. 5:61-70. [ Links ]
6. CAKIR, M.; POULSEN, D.; GALWEY, N.W.; ABLETT, G.A.; CHALMERS, K.J.; PLATZ, G.J.; PARK, R.F.; LANCE, R.C.M.; PANOZZO, J.F.; READ, B.J.; MOODY, D.B.; BARR, A.R.; JOHNSTON, P.; LI, C.D.; BOYD, W.J.R.; GRIME, C.R.; APPELS, R.; JONES, M.G.K.; LANGRIDGE, P. 2003. Mapping and QTL analysis of the barley population Tallon × Kaputar. Australian J. Agric. Res. 54:1155-1162. [ Links ]
7. CASTAÑEDA-SAUCEDO, M.C.; LÓPEZ-CASTAÑEDA, C.; COLINAS-DE LEÓN, M.T.; MOLINA, J.; HERNÁNDEZ, A. 2009. Rendimiento y calidad de la semilla de cebada y trigo en campo e invernadero. Interciencia. 34(4):286-292. Disponible desde Internet en: . http://www.scielo.org.ve/scielo.php?script=sci_ arttext&pid=S0378-18442009000400013&lng=es&n rm=iso ISSN 0378-1844 (con acceso el 18/02/10). [ Links ]
8. CONRY, M.J. 1994. Comparative effect of six cultivars at four rates of nitrogen on the grain yield and grain quality of spring-sown malting barley in Ireland. J. Agric. Sci. 122:343-350. [ Links ]
9. ESHGHI, R.; AKHUNDOVA, E. 2009. Genetic analysis of grain yield and some agronomic traits in hulless barley. African J. Agric. Res. 4(12):1464-1474. Disponible desde Internet en: http://www.academicjournals.org/ AJAR (con acceso el 18/02/10). [ Links ]
10. ESHGHI, R.; OJAGHI, J.; RAHIMI, M.; SALAYEVA, S. 2010. Genetic characteristics of grain yield and its components in barley (Hordeum vulgare L.) under normal and drought conditions. American-Eurasian J. Agric. & Environ. Sci. 9(5):519-528. [ Links ]
11. FENALCE. 2009. Indicadores sectoriales. Febrero 18 de 2009. Disponible desde Internet en: http://www.fenalce.org/archivos/Indicadores.pdf (con acceso el 15/03/10). [ Links ]
12. GALLAGHER, N.; BISCOE, P.; SCOTT, R. 1975. Barley and its environment. V. Stability of Grain Weight. J. Appl. Eco. 12(1):319-336. Disponible desde Internet en: http://www.jstor.org/stable/2401735 (con acceso el 25/02/10). [ Links ]
13. GARCÍA DEL MORAL, L.F.; RAMOS, J.M.; GARCÍA DEL MORAL, M.B.; JIMÉNEZ-TEJADA, M.P. 1991. Ontogenetic approach to grain production in spring barley based on path-coefficient analysis. Crop Sci. 31:1179-1185. [ Links ]
14. GARCÍA DEL MORAL, L.F.; GARCÍA DEL MORAL, M.B.; MOLINA-CANO, J.L.; SLAFER G.A. 2003. Yield stability and development in two- and six-rowed winter barleys under Mediterranean conditions. Field Crops Res. 81:109-119. [ Links ]
15. ISLAM, A.A.; DARRAH, M. 2005. Estimation of gene effects for seed yield and component traits in hulless barley. Turk. J. Field Crops. 8:85-92. [ Links ]
16. KJAER, B.; JENSEN, J. 1996. Quantitative trait loci for grain yield and yield components in a cross between a six-rowed and a two-rowed barley. Euphytica. 90(1):39-48. Disponible desde Internet en: http://www. springerlink.com/content/u61x267854t51444/fulltext. pdf?page=1 (con acceso el 27/03/10). [ Links ]
17. LI, J.Z.; HUANG, X.Q.; HEINRICHS, F.; GANAL, M.W.; RODER, M.S. 2006. Analysis of QTLs for yield components, agronomic traits, and disease resistance in an advanced backcross population of spring barley. Genome (Canada). 49:454-466. [ Links ]
18. MITCHELL, J.H.; FUKAI, S.; COOPER, M. 1996. Influence of phenology on grain yield variation among barley cultivars grown under terminal drought. Australian J. Agric. Res. 47:757-774. [ Links ]
19. OKUYAMA, L.A.; FEDERIZZI, L.C.; BARBOSA NETO, J.F. 2005. Plant traits to complement selection based on yield components in wheat. Ciência Rural, Santa Maria. (Brazil). 35(5):1010-1018. [ Links ]
20. RASMUSSON, D.C. 1985. Barley. American Society of Agronomy, Crop Science Society of America, Soil Science Society of America. Publishers, Madison, Wisconsin. 522p. [ Links ]
21. RICO, E. 1980. El cultivo de la cebada cervecera en Colombia. Bavaria S.A. p.2-4. [ Links ]
22. ROHMAN, M.M.; SULTANA, R.; PODDER, R.; TANJIMUL, A.T.M.; ISLAM, M.K.; ISLAM, M.S. 2006. Nature of gene action in barley (Hordeum vulgare L.). Asian J. Plant. Sci. 5:170-173. [ Links ]
23. SHEEHY, J.E.; DIONORA, M.J.A.; MITCHELL, P.L. 2001. Spikelet numbers, sink size and potential yield in rice. Field Crops Res. 71:77-85. [ Links ]
24. SINEBO, W. 2002. Crop ecology, production & management. Yield relationships of barleys grown in a tropical highland. Crop Sci. 42:428-437. Disponible desde Internet en: http://www.highbeam.com/doc/1G1-84214183.html (con acceso el 12/01/09). [ Links ]
25. SINGH, I.; DASHORA, S.L.; SHARMA, S.N.; SASTRY, E.V.D.; SINGH, I. 1999. Inheritance of some quantitative characters in six-rowed barley. Indian. J. Genet. 59:99- 101. [ Links ]
26. VERMA, A.K.; VISHWAKARMA, S.R.; SINGH, P.K. 2007. Genetic architecture for yield and quality component traits over two environments in barley (Hordeum vulgare L.). Barley Genet. Newslett. 37:24-28. [ Links ]
Recibido: Mayo 7 de 2011; Aceptado: Noviembre 3 de 2011
