










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCaldasia
Print version ISSN 0366-5232On-line version ISSN 2357-3759
Caldasia vol.33 no.1 Bogotá Jan./June 2011
Sexual dimorphism and geographic variation of the blind snake Typhlops reticulatus (Scolecophidia: Typhlopidae) and distribution of species of the genus in Colombia
JOSÉ RANCES CAICEDO-PORTILLA
Laboratorio de Anfibios, Grupo de Cladística Profunda y Biogeografía Histórica, Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá, Colombia. jrcaicedop@unal.edu.co
RESUMEN
El género Typhlops es de distribución pantropical. Por sus hábitos fosoriales, son serpientes pobremente representadas en las colecciones, dificultando trabajos de tipo taxonómico debido a un tamaño insuficiente de la muestra. Se revisaron 56 especímenes depositados en las colecciones de Colombia. En la actualidad, sólo tres especies de Typhlops son conocidas para Colombia (T. brongersmianus, T. minuisquamus y T. reticulatus). Se amplía el área de distribución geográfica en Colombia de T. brongersmianus y T. minuisquamus alcanzando las estribaciones centro-orientales de la Cordillera Oriental. Se reportan especímenes adicionales de T. reticulatus con distribución transandina. Los rangos merísticos y cuantitativos se encuentran dentro lo reportado por la literatura para T. brongersmianus. T. minuisquamus presenta un mayor número de escamas dorsales totales y variación en las formulas de escamas dorsales con respecto a lo reportado, y el porcentaje de la fórmula de escamas dorsales en T. reticulatus varío considerablemente con lo reportado en la literatura. Se encontraron diferencias significativas en la longitud total del cuerpo, teniendo hembras con mayor tamaño que los machos, así mismo se hallaron diferencias significativas en el número de escamas dorsales totales entre las poblaciones cis y transandinas. A pesar de presentar variación geográfica, no se puede argumentar que las poblaciones separadas geográficamente puedan representar unidades evolutivas independientes. Pos esta razón, es necesario realizar estudios morfológicos de mayor profundidad que puedan ofrecer mayor información y evidencia, ya que T. reticulatus representa un complejo de especies que necesita una revisión urgente dentro del rango de distribución geográfica de esta especie.
Palabras clave. Typhlops, Colombia, Dimorfismo sexual, Variación geográfica, Distribución geográfica.
ABSTRACT
Typhlops has a pantropical distribution. Due to their fossorial habits, these snakes are poorly represented in collections, making it difficult to take taxonomic data. I verified all Colombian collections and registered 56 specimens. Only three species of Typhlops are known for Colombia: Typhlops brongersmianus, T. minuisquamus, and T. reticulatus. The geographic distribution of T. brongersmianus and T. minuisquamus are extended to the eastern flank of the Oriental Cordillera. Additional records of T. reticulatus are given. The meristics and quantitative ranges are in agreement with literature data for T. brongersmianus. Typhlops minuisquamus has a higher number of total dorsal scales and variation in the dorsal scales formulae than previously reported as in T. reticulatus. I found significant intraspecific differences in the total body length, in which females are greater than males, and a cis- and trans-Andean difference in dorsal scale counts. Despite exhibiting a distinct geographical variation, it is not possible to argue that the geographically separate populations are independent evolutionary units. Future morphological studies including more specimens across the species distribution range are necessary.
Key words. Typhlops, Blind-snakes, Colombia, Sexual dimorphism, Geographic variation.
Recibido: 12/03/2010
Aceptado: 12/02/2011
El género Typhlops tiene una distribución pantropical; en el neotrópico el género se encuentra en Centroamérica, Suramérica y las Indias Occidentales (McDiarmid et al. 1999). Son serpientes de hábitos troglobíticos por lo cual son difíciles de recolectar y están pobremente representadas en las colecciones del país.
Los estudios sobre estas serpientes en Colombia son muy pocos, el primer trabajo fue elaborado por Dunn (1944), quien hace un corto comentario sobre Typhlops reticulatus. Posteriormente, Dixon & Hendricks (1979) realizan un estudio sobre el arreglo sistemático, distribución y variación geográfica del género en el neotrópico, excluyendo las especies que se encuentran en las Antillas; en este trabajo, los autores reportan tres especies para Colombia: Typhlops brongersmianus Vanzolini 1972 de la isla Santa Sofía en el extremo sur del departamento del Amazonas; Typhlops minuisquamus Dixon & Hendricks 1979 reportada en la localidad de Timbó en el departamento de Vaupés y Typhlops reticulatus Linnaeus 1758, reportada al oriente de la cordillera de los Andes, la Orinoquía y Amazonía en Colombia (Dixon & Hendricks 1979), estos autores reportan un único espécimen de T. reticulatus al occidente de los Andes en el Ecuador. Aunque Dixon & Hendricks (1979) documentaron variación geográfica en T. reticulatus, ellos no hicieron arreglos taxonómicos, por la posibilidad que la variación geográfica podría estar causada por el dimorfismo sexual.
Recientemente, Carvajal-Cogollo (2006) reportó el segundo espécimen de T. reticulatus con distribución al occidente de los Andes, al norte de la Cordillera Occidental en Colombia, argumentando que su registro es el que se encuentra más al norte en la distribución de esta serpiente, sin embargo, Carvajal-Cogollo (2006) no revisó el trabajo de Dixon & Hendricks (1979), quienes tienen dos reportes en Paujil en el estado Falcón de Venezuela. Otros dos estudios que tratan a cerca de estas especies fueron los realizados por Pérez-Santos & Moreno (1988) y Sánchez et al. (1995). El primer trabajo fue fuertemente criticado por Cadle (1992), debido principalmente a poca citación de especímenes de museo que soporten la mayoría de los datos sistemáticos, de distribución e historia natural que aparecen en el libro; dudas a cerca de los datos de distribución e historia natural pues estos no son documentados y son fuertemente cuestionables; y su trabajo se basó en la consulta de fuentes secundarias en el tratamiento de las especies, este error se dio principalmente porque ellos basaron su trabajo en los datos consignados en los catálogos de los museos consultados por estos autores o terceras personas, sin una revisión directa de los especímenes depositados y de los datos consignados en estos museos (Lynch, com. pers.). El segundo trabajo es un listado, que se realizó con base en la consulta y recopilación de una gran cantidad de trabajos donde se citan las especies y localidades hasta ese momento registradas para Colombia (a pesar de esto, Sánchez et al. 1995 no citan el trabajo de Dixon & Hendricks 1979; por lo que estos autores no reportan a T. brongersmianus y T. minuisquamus para Colombia), Sánchez et al. (1995) no realizaron una revisión directa de los especímenes depositados en las colecciones herpetológicas del país. Finalmente, Castaño-Mora et al. (2004), en su trabajo sobre los reptiles del Chocó biogeográfico argumentan que la familia Typhlopidae muestra una distribución solo cis-andina, no teniendo en cuenta el registro de Dixon & Hendricks (1979) y la consulta de otras colecciones del país como la del Instituto Alexander von Humboldt, donde se encontraba un espécimen de la costa Pacífica colombiana.
Teniendo en cuenta lo anterior, se hace necesaria la revisión de los especimenes depositados hasta el momento en las colecciones colombianas.
En la revisión de los ejemplares depositados en las colecciones de Colombia, se encontró que la mayoría de serpientes objeto de este estudio estaban catalogadas como T. reticulatus, no diferenciándose las otras dos especies del género que se encuentran en Colombia, también se encontró serpientes de los géneros Tricheilostoma, Epictia, LioTyphlops y Typhlophis en los frascos donde se encontraban los tiflópidos mencionados anteriormente. Aunque las colecciones herpetológicas Colombianas tienen depositadas gran cantidad de material y de especies (obs. pers.), muchos de los registros que se encuentran escritos en los catálogos son identificaciones incorrectas, subestimando la verdadera riqueza de ofidios que están presentes en el país.
El objetivo de este trabajo fue realizar la identificación de los especímenes depositados en las diferentes colecciones de Colombia, realizar los mapas de distribución de las especies del género que se encuentran en el país y analizar si hay variación geográfica entre las poblaciones cis y transandinas de T. reticulatus, y en caso de que se encuentre dicha variación, analizar si esta variación se ve influenciada por el dimorfismo sexual de los especímenes estudiados.
MATERIALES Y MÉTODOS
Se revisó el material depositado en las colecciones herpetológicas de las siguientes instituciones: Instituto de Ciencias Naturales de la Universidad Nacional de Colombia, Bogotá (ICN); Museo de la Universidad de La Salle, Bogotá (MLS); Museo de Historia Natural de la Universidad Industrial de Santander, Bucaramanga (UIS-R); Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, Villa de Leyva (anteriormente IND-R, ahora IAvH); Museo de Historia Natural de la Pontificia Universidad Javeriana, Bogotá (MUJ) y del museo de Historia Natural de la Universidad de Antioquia, Medellín (MHUA). La colección personal de Hollman Miller, Mitú (HM) fue revisada por John D. Lynch.
Las medidas que se tomaron fueron la longitud total (Lt), longitud de la cola (Lc) y ancho en la mitad del cuerpo (Ac), utilizando cinta métrica y calibrador píe de rey marca Koala® a 0.1 y 0.02 mm de precisión respectivamente; también se analizaron dos radios relacionados con estas medidas: Lt/Lc y Lt/Ac. Se tomo igualmente tres caracteres cuantitativos: número total de escamas dorsales, número total de escamas subcaudales y número de escamas alrededor del cuerpo en la parte anterior, media y posterior (formula de escamas dorsales). Las medidas y caracteres cuantitativos se tomaron siguiendo a Dixon & Hendricks (1979).
Para determinar el sexo de los individuos se realizó una incisión ventral en el tercio posterior del cuerpo observándose la presencia de testículos y hemipenes evertidos, en el caso de los machos; y folículos ováricos, folículos vitelogénicos y huevos oviductales en el caso de las hembras; estas observaciones se basaron en los trabajos realizados por Robb (1960), Fox & Dessauer (1962) y Fox (1965).
Para analizar el dimorfismo sexual y la variación geográfica entre los especímenes de distribución cis y transandina de Typhlops reticulatus (figura 1), se utilizó estadística univariada no paramétrica (prueba de U-Mann Whitney) cuando los datos no mostraban una distribución normal y paramétrica (Prueba de T) cuando mostraban una distribución normal para poder realizar dichas pruebas. Para todos los análisis estadísticos se considero una significancia de a = 0.05.
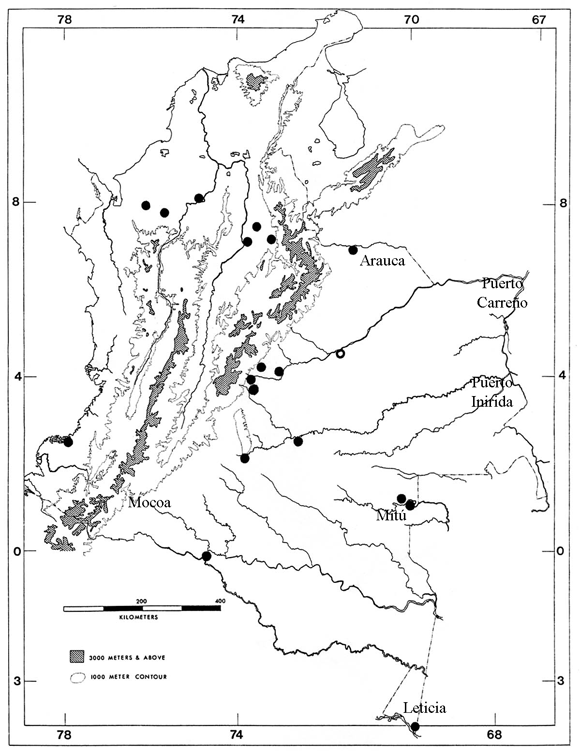
Figura 1. Distribución cis y transandina de T. reticulatus en Colombia. Los círculos abiertos corresponden a registros de literatura.
Un total de 27 localidades fueron georeferenciadas utilizando principalmente el gacetero en línea GEONAMES: http://www.geonames.org, utilizando el sistema de coordenadas WGS84 (World Geodetic System 1984); los mapas de distribución fueron elaborados en ArcGis versión 9.1 (ESRI, GIS and Mapping Software).
RESULTADOS
En total se examinaron 56 (más nueve Typhlops minuisquamus, revisados por John D. Lynch) especímenes de las tres especies registradas hasta el momento para Colombia (seis de T. brongersmianus, doce de T. minuisquamus y 47 de T. reticulatus) (Anexo 1).
Se encontraron especímenes del flanco oriental de la Cordillera Oriental, en la región de Villavicencio de T. brongersmianus y de T. minuisquamus¸ así como especímenes adicionales con distribución transandina de T. reticulatus, con lo que se amplía considerablemente la distribución de estos tres taxones dentro del país, así mismo se amplia la distribución altitudinal de T. reticulatus a 812 m, sobrepasando en casi 300 metros lo reportado por Dixon & Hendricks (1979), los cuales reportan una altura de 500 m aproximadamente (figura 1, figura 2 y figura 3).
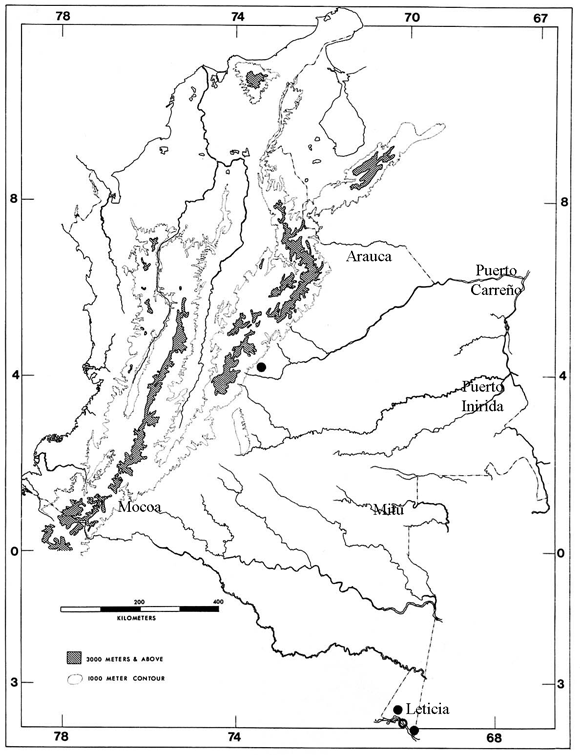
Figura 2. Distribución de T. brongersmianus en Colombia. Los círculos abiertos corresponden a registros de literatura.
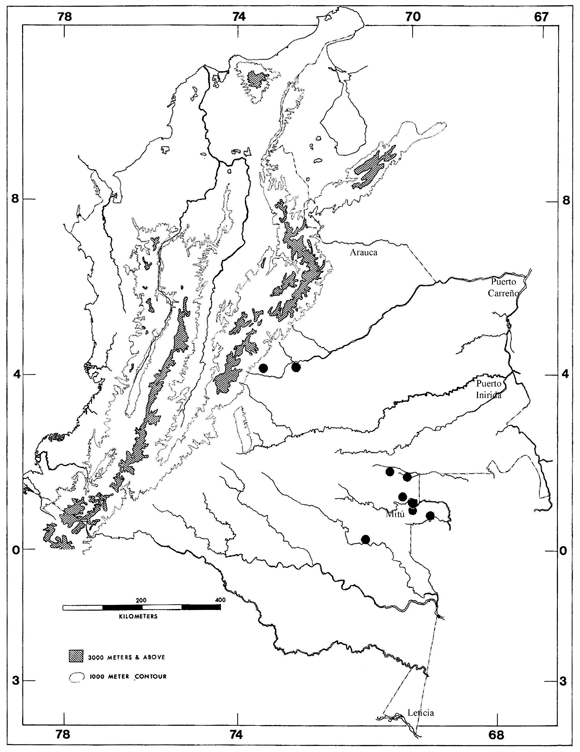
Figura 3. Distribución de T. minuisquamus en Colombia.
De los 47 especímenes preservados de T. reticulatus, 38 son de la región cisandina y nueve de la región transandina (figura 1); de estos especímenes, 28 corresponden a hembras y 16 son machos. Tres especímenes no se pudieron sexar.
De los especímenes examinados, 26 tienen conteos alrededor del cuerpo 20-20-20 (55.32%); 17 tienen 20-20-18 (36.17%) y cuatro especímenes tienen conteos de 20-20-19 (8.51%), porcentajes muy disímiles a lo que encontraron Dixon & Hendricks (1979), quienes reportan sólo el 2% de los especímenes estudiados por ellos con conteos de 20-20-20, es decir, de los 120 especímenes analizados por estos autores, sólo cuatro tenían conteos de 20-20-20 ó 20-20-19 (J. R. Dixon, com. pers.). Al separar estos conteos entre las poblaciones transandinas y cisandinas, se encuentra que el porcentaje de los conteos alrededor del cuerpo es el siguiente: en la muestra de 10 especímenes de la población transandina (incluyendo el espécimen de Ecuador reportado por Dixon & Hendricks) se encontró que el 70% de la muestra tiene conteos de escamas de 20-20-20, el 20% tienen conteos de 20-20-18 y el 10% tiene conteos de 20-20-19 (Figura 6); de los 38 especímenes de la región cisandina encontrados en este estudio, el 50% tienen conteos de 20-20-20, el 42% tienen conteos de 20-20-18 y el 8% tienen conteos de 20-20-19.
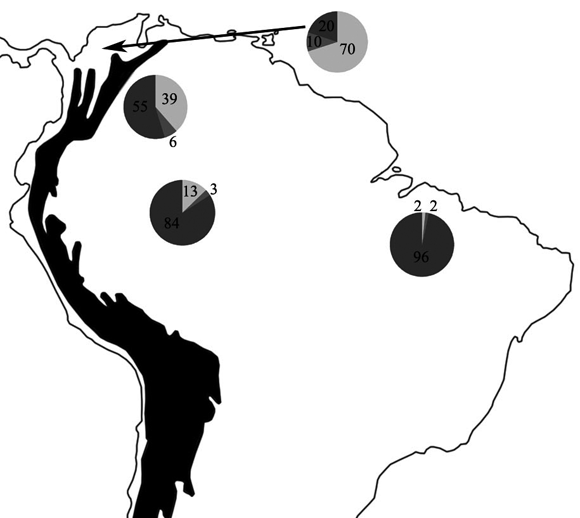
Figura 6. Distribución del porcentaje de las diferentes fórmulas de escamas dorsales en T. reticulatus en Suramérica. 20-20-20; 20-20-19; 20-20-18. Los números representan los porcentajes de cada una de las fórmulas.
Dimorfismo sexual
Typhlops reticulatus muestra diferencias significativas en la longitud total del cuerpo (t41, 0.05= -2.14; P= 0.03), siendo las hembras más grandes que los machos (figura 4); pero no presento dimorfismo sexual en: número de escamas dorsales del cuerpo (t41, 0.05= -1.23; P= 0.23); número de escamas ventrales de la cola (U16, 28= 170; P= 0.19); longitud de la cola (U16, 28= 204; P= 0.63) tampoco en el radio Lt/Ac (t41, 0.05= 1.32 ; P= 0.19) ni en el radio Lt/Lc (t41, 0.05= -1.11 ; P= 0.27).
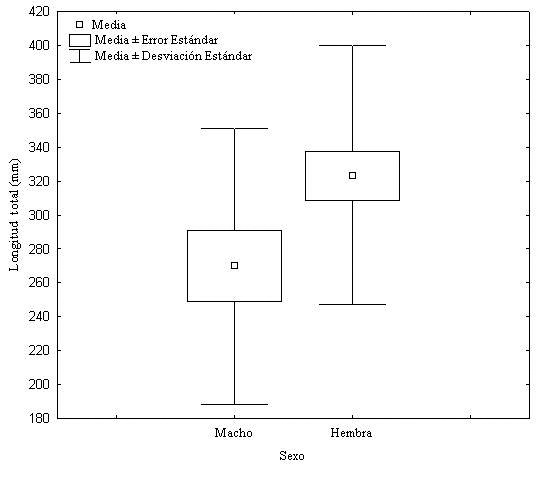
Figura 4. Dimorfismo sexual en la longitud total del cuerpo (en mm) en T. reticulatus en Colombia.
Variación geográfica
Los especímenes con distribución cis y transandina de Typhlops reticulatus presentaron diferencias altamente significativas en el número de escamas dorsales del cuerpo (t44, 0.05= 8.06; P= 0) y en el radio Lt/Ac (t44, 0.05 = 3.46; p=0.0011), teniendo la muestra de distribución cisandina mayor número de escamas dorsales y valor en el radio Lt/Ac que la muestra con distribución transandina (figura 5); la muestra no presentó diferencias significativas en: longitud total del cuerpo (U44, 0.05= -0.79; P= 0.42); el número de escamas ventrales de la cola (t45, 0.05 = 1.465; p = 0.14); la longitud de la cola (t45, 0.05= -0.7727; p = 0.44) y tampoco en el radio Lt/Lc (t44,0.05 = 0.69; p = 0.49).
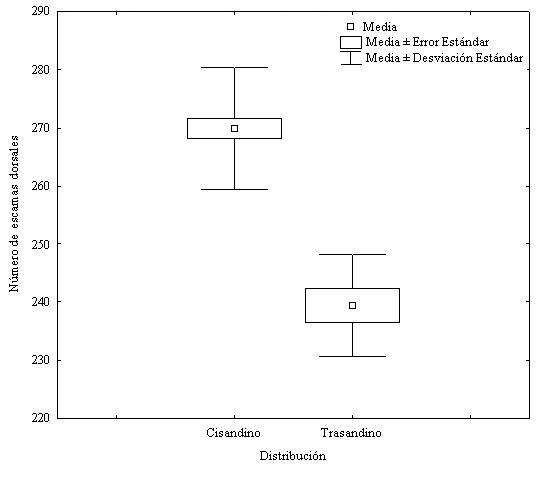
Figura 5. Variación Geográfica en el número total de escamas dorsales del cuerpo de las poblaciones cis y transandinas de T. reticulatus en Colombia.
Los conteos de escamas de los seis especímenes analizados de Typhlops brongersmianus como escamas dorsales totales varía de 243-261 (X= 247.5); número de subcaudales 11-13 (X= 12.2); formula de escamas dorsales (83.3% con 20-20-20 y 16.3% con formula 20-20-18), los porcentajes y promedios presentados en el presente estudio difieren a lo reportado por Dixon & Hendricks (1979), pero los conteos están dentro de los rangos reportados por estos autores, esta diferencia se puede deber al poco material disponible para este estudio; el tamaño y coloración de los especímenes estudiados son similares a lo reportado por Dixon & Hendricks (1979).
Los conteos de escamas dorsales totales de los doce especímenes analizados de T. minuisquamus varía de 214-268 (X= 240.1); el número de subcaudales 7-11 (X= 8.6); fórmula de escamas dorsales totales (dos individuos con conteos 20-18-16, uno con 18-16-14, uno con 18-18-15, cinco con 18-18-14 y tres con 20-18-14). Los rangos y porcentajes del número de escamas dorsales totales varía de acuerdo a lo reportado por Dixon & Hendricks (1979), quienes dan un rango de 221-253; y la fórmula de escamas dorsales totales también varían considerablemente a lo reportado por Dixon & Hendricks (1979), quienes sólo reportan las fórmulas 20-18-16 y 18-16-14; el resto de conteos, medidas y patrón de coloración están dentro del rango reportado por estos autores.
DISCUSIÓN
Al igual que Dixon & Hendricks (1979), en este trabajo se encontró que el número total de escamas dorsales desde la rostral hasta la espina de la cola, es el carácter que mostró diferencias significativas en cuanto a variación geográfica entre las muestras de las poblaciones cis y transandinas de Typhlops reticulatus, pero estos autores sospechan que sus datos pueden estar influenciados por un fuerte dimorfismo sexual en este carácter; no obstante, en la muestra estudiada se pudo analizar el dimorfismo sexual en esta especie, encontrando únicamente diferencias significativas en la longitud total del cuerpo, más no en el número total de escamas dorsales del cuerpo; por ejemplo, una hembra (UIS-R-1666) mide 502 mm de longitud total, pero sólo tiene 250 escamas dorsales totales; por lo que se puede argumentar que la variación geográfica en cuanto a este carácter no se ve influenciada por el dimorfismo sexual o el mayor tamaño de las hembras, como lo encontró Roux-Esteve (1975) en algunas especies de tiflópidos africanos.
En cuanto al patrón de variación geográfica encontrado por Dixon & Hendricks (1979) encontraron una clina geográfica que va incrementando el número de escamas dorsales totales desde el noroccidente al suroriente. En el presente estudio se encontró el mismo patrón, con la muestra de las poblaciones transandinas (noroccidente) con el menor número de escamas dorsales, y las poblaciones cisandinas (suroriente) con el mayor número de escamas dorsales, mostrando así un incremento de las escamas desde el noroccidente hacia el suroriente de Colombia.
El carácter de la fórmula de las escamas dorsales, también mostró grandes diferencias a lo que reportan Dixon & Hendricks (1979), se encontró que el 63.83% de la muestra tiene fórmulas de escamas 20-20-20 y 20-20-19, un porcentaje bastante alto a lo reportado por estos autores (sólo el 2% presentan estas formulas), al analizar la distribución de las formulas de las escamas dorsales no hay un patrón evidente de distribución en cuanto a este carácter, ya que se encuentran especímenes de una misma localidad con fórmulas diferentes (p. ej. en Leticia, Amazonas); así mismo, en la muestra transandina hay dos especímenes con formulas de 20-20-18 y 20-20-19 y en la muestra cisandina el carácter también esta distribuido de una manera azarosa, lo que puede indicar que hubo un flujo de genes entre las poblaciones cis y transandinas antes del levantamiento de Los Andes del norte de América del Sur. Pero al realizar un agrupamiento de los porcentajes de las fórmulas de las escamas dorsales analizados en este trabajo con los datos de Dixon & Hendricks (1979), exceptuando los tres registros de Venezuela; se puede observar un patrón de aumento en la aparición del porcentaje de la fórmula 20-20-18 (Figura 6). Se encontró que en los especímenes transandinos sólo el 20% de las serpientes estudiadas tienen esta fórmula (N=10); en las serpientes de los llanos Orientales de Colombia, el porcentaje de aparición de especímenes con la fórmula 20-20-18 aumenta a un 55% de la muestra analizada (N=31), mientras en los especímenes del occidente y centro de la Amazonía el porcentaje de esta fórmulas es del 84% (N=63) y en las serpientes que se encuentran distribuidas en el oriente de la Amazonía, y las Guyanas el porcentaje de esta fórmula es del 96% en la muestra estudiada (N=57); es decir, a medida que aumenta el porcentaje de la fórmula de escamas dorsales con 20-20-18, va disminuyendo el porcentaje de la fórmula con conteos de 20-20-20, encontrando las serpientes transandinas con un porcentaje del 70% con esta fórmula y las del oriente de la Amazonía y las Guyanas sólo con el 2% (Figura 6). Al igual que el aumento en el número de escamas dorsales totales, el aumento del porcentaje de la fórmula de escamas dorsales con arreglo 20-20-18, también aumenta del noroccidente al suroriente, demostrando así la clina geográfica que habían encontrado Dixon & Hendricks en su estudio.
Dixon & Hendricks (1979) se abstienen de nombrar subespecies de T. reticulatus debido principalmente a la sospecha de que sus datos sobre número total de escamas dorsales podrían estar influenciados por el dimorfismo sexual que puede presentar este carácter, aunque en el presente estudio se encontró que el dimorfismo sexual no influye sobre el número total de escamas dorsales, y a pesar de la distribución disyunta o separada por la Cordillera de los Andes entre las poblaciones cis y transandinas, y a la variación que muestra el número de escamas dorsales totales, esta separación no es total, pues hay un ejemplar de la región transandina que tiene 250 escamas dorsales totales (UIS-R-1666), al igual que un espécimen de la región cisandina que tiene 249 escamas (MLS-045). Este traslape en el número de escamas dorsales totales también puede indicar que hubo un flujo de genes entre las poblaciones cis y transandinas, antes de que empezara el levantamiento de la Cordillera Oriental colombiana en la época del Mioceno Medio, comprendido entre los 12.9 y 11.8 millones de años atrás (Guerrero 1997); ya que la familia Typhlopidae se puede considerar como un grupo relativamente antiguo y de distribución gondwánica (Thomas 1976), pues muchos de los caracteres anatómicos de la familia Typhlopidae y las familias de serpientes ciegas asociadas (Anomalepididae y Leptotyphlopidae) son considerados como primitivos (Dixon & Hendricks 1979), se puede considerar que antes del levantamiento de la Cordillera Oriental de Colombia, la serpiente T. reticulatus se encontraba ampliamente distribuida en la parte norte de Suramérica.
Por lo tanto, en este estudio, no es conveniente hablar de subespecies pues esta es una práctica que se ha utilizado a principios y mitad del siglo XX, y muy común entre ornitólogos, mastozoólogos y algunos herpetólogos, y de acuerdo con lo expuesto por Wilson & Brown (1953) y Thorpe (1980), quienes argumentan que los patrones de diferenciación entre poblaciones (p. ej. variación geográfica, coloración y tamaño, entre otros) se ven oscurecidos por el nombramiento de un gran número de subespecies sin sentido evolutivo alguno, por lo que ha habido una tendencia en seccionar las clinas geográficas en categorías artificiales y la delimitación de subespecies arbitrariamente con base en rasgos fisiográficos, como la separación de poblaciones por cadenas montañosas o ríos. Thorpe (1980) cuestiona que muchos herpetólogos europeos basan sus subespecies en la diferencia de un solo carácter, no teniendo en cuenta el factor evolutivo, en el cual diferentes caracteres pueden mostrar diferentes patrones de variación geográfica, entonces por el simple hecho de que una población difiera de otra, por ejemplo, en el conteo de escamas ó en el patrón de coloración, no es un argumento suficiente para que estas poblaciones sean tratadas como subespecies.
A pesar de que en la actualidad se pudiera hablar de dos unidades evolutivas independientes, debido a la separación geográfica de las poblaciones por la cordillera de los Andes, esto no se puede argumentar con T. reticulatus, teniendo en cuenta la muestra estudiada. Por lo anterior es importante que a futuro se realicen estudios más detallados de morfología interna (p. ej. estructura del cráneo y hemipenes), genética de poblaciones o estudios moleculares que puedan dar mayor información a cerca del flujo de genes, la deriva génica y los procesos de selección natural que puedan influir o no en la conformación de estas unidades evolutivas independientes separadas por la cadena montañosa del norte de Los Andes, ya que al parecer T. reticulatus puede representar un complejo de especies en todo su areal de distribución en Sur América (Passos com. pers.).
La distribución disyunta o separada que presentan los registros de T. brongersmianus y T. minuisquamus en el presente trabajo se debe a la falta de muestreos sistemáticos en la región cisandina de Colombia; por lo que se deben realizar estudios a cerca de la distribución real de este grupo de serpientes en el país, esto con el fin de poder entender sus patrones de distribución, y si la variación geográfica que presenta T. brongersmianus está o no influenciada por el sexo; y si se presenta variación geográfica o no en T. minuisquamus, especie distribuida principalmente en la cuenca amazónica (Dixon & Hendricks 1979). Aunque las muestras en los museos colombianos son relativamente escasas, tal vez debido a los hábitos secretivos de estas especies, en muchas regiones donde se han realizado mayores esfuerzos de muestreo y con las técnicas apropiadas (p. ej. Trampas de caída con cercos de aluminio), se han podido encontrar especies que son relativamente comunes en ciertas áreas de su distribución (Passos et al. 2005, 2006).
Por lo tanto, se hace necesaria la recolecta de material adicional de estas serpientes tiflópidas para poder obtener más datos y evidencias, ya que Colombia hace parte de la distribución del extremo occidental de estas tres especies.
AGRADECIMIENTOS
A los curadores de las siguientes instituciones, quienes permitieron el acceso y la revisión de los especímenes: John D. Lynch (ICN), Martha Patricia Ramírez (UIS), Vivian Páez (MHUA), Diego Perico (IND-R y IAvH), hermano José Espitia y "Arturito" Rodríguez (MLS) y Julio Mario Hoyos (MUJ). Agradecimientos especiales a John D. Lynch, mi maestro y amigo, por el préstamo de bibliografía y los mapas de Colombia; así como a las nutridas discusiones acerca del tema. A Sandy Arroyo del Laboratorio de Anfibios del ICN, Mónica Jiménez Castro y Jorge Gualdrón de Conservación Internacional-Colombia por la realización de los mapas de distribución usados o no en este estudio. A Claudia "Julis" Dulcey por la realización del mapa de la figura 6 y a las correcciones de forma del trabajo, a Diego "cachorro" Rincón y Henry Alberto Suárez por la realización de las gráficas y la asesoría estadística. A Martha Patricia Ramírez y David Sánchez por la búsqueda y préstamo de literatura clave en la realización de este trabajo, a Gustavo Adolfo Ballén por las correcciones realizadas al resumen en inglés, finalmente agradecerle a los dos revisores anónimos por los comentarios y correcciones constructivas al manuscrito.
LITERATURA CITADA
1. CADLE, J.E. 1992. On Colombian Snakes. Herpetologica 48 (1): 134-143. [ Links ]
2. CARVAJAL-COGOLLO, J.E. 2006. Geographic Distribution: Typhlops reticulatus, Herpetological Review 37(4): 501-502. [ Links ]
3. CASTAÑO-MORA, O.V., G. CÁRDENAS-A., E.J. HERNÁNDEZ-R. & F. CASTRO-H. 2004. Reptiles en el Chocó biogeográfico. En: Rángel-Ch., J. O. (ed.), Colombia Diversidad Biótica IV: El Chocó biogeográfico/Costa Pacífica. Universidad Nacional de Colombia, Bogotá, p.p. 599-632. [ Links ]
4. FOX, W. 1965. A comparison of the male urogenital systems of blind snakes, Leptotyphlopidae and Typhlopidae. Herpetologica 21(4): 241-256. [ Links ]
5. FOX, W. & H.S. DESSAUER. 1962. The single right oviduct and other urogenital structures of female Typhlops and LeptoTyphlops. Copeia 3: 590-597. [ Links ]
6. DIXON, J.R. & F.S. HENDRICKS. 1979. The Wormsnakes (Family Typhlopidae) of the Neotropics, Exclusive of the Antilles, Zoologische Verhandelingen (Leiden) 173: 1-39. [ Links ]
7. DUNN, E.R. 1944. A Review of the Colombian Snakes of the Families Typhlopidae and Leptotyphlopidae, Caldasia 3 (11): 47-55. [ Links ]
8. GUERRERO, J. 1997. Stratigraphy, Sedimentary, Environments and Miocene Uplift of the Colombian Andes. en: R. F. kay, R. H. Madden, R. L. Cifelli & J. J. Flynn (eds), Vertebrate Paleontology in the Neotropics: The Miocene Fauna of La Venta, Colombia. Smithsoniam Institution Press, Washington, D. C. pp 5-43. [ Links ]
9. MCDIARMID, R. W., J. A. CAMPBELL & T. A. TOURÉ. 1999. Snake Species of the World: a Taxonomic and Geographic Reference. The Herpetologists' League, Washington D.C. 1: 511. [ Links ]
10. PASSOS, P., U. CARAMASCHI & R.R. PINTO. 2005. Rediscovery and Redescription of LeptoTyphlops salgueiroi Amaral, 1954 (Squamata, Serpentes, Leptotyphlopidae). Boletim do Museu Nacional, nov. série Zoologia 520: 1-10. [ Links ]
11. PASSOS, P., U. CARAMASCHI & R.R. PINTO. 2006. Redescription of LeptoTyphlops koppesi Amaral, 1954, and Description of a New Species of the LeptoTyphlops dulcis group from Central Brazil (Serpentes: Leptotyphlopidae). Amphibia-Reptilia. 27: 347-357. [ Links ]
12. PÉREZ-SANTOS, C. & A.G. MORENO. 1988. Ofidios de Colombia. Monografie VI, Museo Regionale di Scienze Naturali di Torino. 517pp. [ Links ]
13. ROBB, J. 1960. The internal anatomy of Typhlops Schneider (Reptilia). Australian Journal of Zoology 8(2): 181-216. [ Links ]
14. ROUX-ESTEVE, R. 1975. Recherches Sur la Morphologie, la Biogéographie et la Phylogenie des Typhlopidae d´Afrique. Bulletin de I´nstitut Fondamental d´Afrique Noire 36: 428-508. [ Links ]
15. SÁNCHEZ-C, H., O.V. CASTAÑO-M., & G. CÁRDENAS-A. 1995. Diversidad de los Reptiles en Colombia. En: J.O. Rangel-Ch. (ed.). Colombia Diversidad Biótica I. Universidad Nacional de Colombia, Bogotá, pp. 277-325. [ Links ]
16. THOMAS, R. 1976. Systematics of Antillean snakes of the genus Typhlops (Serpentes: Typhlopidae). Dissertation, 288 pp. Department of Zoology. Loussiana State University 46: 1-5. [ Links ]
17. THORPE, R. S. 1980. Microevolution and Taxonomy of European Reptiles With Particular References to the Grass Snakes Natrix natrix and the Wall Lizards Podarcis sicula and P. melisellensis. Biological Journal of the Linnean Society 14: 215-233. [ Links ]
18. WILSON, E.O. & W.L. BROWN JR. 1953. The Subspecies Concept and its Taxonomic Application. Systematic Zoology 2 (2): 97-111. [ Links ]

