










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkDYNA
Print version ISSN 0012-7353On-line version ISSN 2346-2183
Dyna rev.fac.nac.minas vol.78 no.166 Medellín Apr./June 2011
MODELLING THE DYNAMICS OF ANTIBODY PRODUCTION IN BIRDS
MODELO DE LA DINÁMICA DE PRODUCCIÓN DE ANTICUERPOS EN AVES
ANDRÉS OLARTE
Universidad Nacional de Colombia, Bogotá, Facultad de Ingeniería, faolarted@unal.edu.co
CARLOS CLAVIJO
Universidad Nacional de Colombia, Bogotá, Facultad de Ciencias, caclavijor@unal.edu.co
HERNANDO DÍAZ
Universidad Nacional de Colombia, Bogotá, Facultad de Ingeniería, hdiazmo@unal.edu.co
Received for review October 14th, 2009, accepted July 9th, 2010, final version August, 9th, 2010
ABSTRACT: This paper presents a dynamical model of avian immune response, for antibody production purposes. Antibodies are fundamental tools for research, diagnosis, and the treatment of several diseases. It has been recognized for some time that antibodies obtained from poultry are an efficient alternative for applications in medicine and biology. Therefore, a dynamical model for this process might be very useful. The model being proposed consists of seven non-linear ordinary differential equations with constant coefficients that represent the main cellular and molecular populations in avian immune response. Values for the parameters were obtained from avian and mammalian literature. In silico-generated responses in terms of antibody concentrations are presented and compared to reported kinetics.
KEYWORDS: Avian immunology, antibodies, dynamical models.
RESUMEN: En este artículo se presenta un modelo dinámico de la respuesta inmune humoral de aves para la producción de anticuerpos específicos. Los anticuerpos son fundamentales para la investigación, diagnóstico y tratamiento de diversas enfermedades. Se ha reconocido que los anticuerpos obtenidos de aves de corral son una opción eficiente para aplicaciones en medicina y biología. Por tanto, un modelo que represente la dinámica de este proceso resulta de gran utilidad. El modelo está compuesto de siete ecuaciones diferenciales ordinarias, no lineales, con coeficientes constantes y representa las principales poblaciones celulares y moleculares de la respuesta inmune de las aves. Los parámetros se obtuvieron a partir de datos reportados en el área de inmunología aviar y mamífera. Las respuestas obtenidas "in silico" en términos de la concentración de anticuerpos se presentan y se comparan contra la cinética reportada.
PALABRAS CLAVE: Inmunología aviar, anticuerpos, modelos dinámicos.
1. INTRODUCTION
The immune system protects organisms from a variety of threats (pathogens) [1]. Depending upon the type of invader, an immune system produces different responses that involve many components working in a coordinated fashion to eliminate the pathogen [2,3,4]. This system can recognize and destroy disease-producing agents using cells, molecules, and organs.
In most vertebrates, the immune system consists of innate and adaptive subsystems [5]. The former subsystem identifies microorganisms that share characteristics on their surfaces, whereas the latter recognizes each antigen (foreign agent that induces an immune response) specifically [2]. The adaptive immune system has cellular (produced by T cells) and humoral (produced by B cells) components. These two types of cells are known as lymphocytes. The specific recognition of the adaptive immune system lies in its capability to create millions of slightly different proteins, each one with the ability to recognize only a small set of antigens [5,6]. A very important feature of the adaptive immune system is memory, meaning it remembers previously encountered antigens. In this way, responses to reinfection (secondary response) are faster and stronger.
Immunoglobulins (Igs) or antibodies are molecules with a huge variability in their three dimensional shape, which are fundamental for immune response [7]. These molecules, produced by adaptive immune system B cells, can bind selectively to the antigen’s surface when their unique conformation fits on some foreign agent’s segments. In addition, antibodies have a constant region that is used as a signal for cells and immune molecules to destroy pathogens [5]. These features make antibodies valuable tools in biomedical and biotechnological applications, underscoring the search for efficient production processes [7,8].
Usually antibody production is carried out by inoculating a foreign agent into a vertebrate (typically a mouse or a rabbit), to elicit an immune system response, followed by one or more booster shots at different times, inducing secondary responses that are larger in terms of antibody levels [9,10]. Antibodies are then extracted from the blood. This implies bleeding or sacrificing the animal [11]. Recently, interest in antibody poultry-based production has grown since antibodies can be obtained from the egg yolks [9,10,12,13]. This technique has many advantages compared to the traditional way, including larger protein quantities, a wider range of application, and a reduction in animal stress [14,15].
Avian antibody production has been the subject of several studies including: bird handling, inoculation procedures, antigen type and concentration, immunoglobulin uses, and DNA vaccines [8,9,12,14,15,16,17]. However, a common characteristic of these experiments is an empiric timetable for booster injections.
On the other hand, mathematical models have been proposed in the context of several biological fields in order to explain complex interactions between organisms, cells, and molecules [18,19,20]. One of these fields is theoretical immunology whose main objective is to build mathematical or computational models that account for several immune system processes [6,21]. The models most often used are written as Ordinary Differential Equations (ODE) that represent population changes as functions of time. This type of model has been employed in descriptions of adaptive immune cells interactions [22,23,24] and the dynamics of antibody production by B cells [25,26,27,28].
2. AVIAN IMMUNE SYSTEM
The avian immune system resembles the better known mammalian one [14,29,30,31,32]. More than 30 years ago it was known that chicken immune systems consist of innate, cellular, and humoral responses, just like mammals [33]. Nevertheless, some differences at the molecular and organic levels are clearly distinguishable. Chickens generate three types (isotypes) of immunoglobulins, i.e., IgM, IgA, and IgG (called IgYs when in the yolk) [34].
The innate response is mediated by macrophages, dendritic cells, natural killer cells, and complement proteins which respond immediately after foreign agent invasion. These cells and molecules use mechanisms similar to those used by their mammalian counterparts [30,35].
Cellular response involves T helper and cytotoxic T cells [32]. T lymphocytes oversee and coordinate other cells’ intracellular activity. In particular, some cells of the innate system (antigen presenting cells, APC), such as macrophages and dendritic cells, expose or "present" antigenic molecules to T helper cells, to activate them.
On the other hand, humoral response, produced by B lymphocytes, begins when a molecule similar to an antibody, located on its plasmatic membrane, recognizes an antigen segment. In addition, active T helper cells release a stimulatory signal for B cell activation. After this process, the B cell population rises and produces antibodies against this particular antigen. This mechanism is called clonal expansion.
3. YOLK ANTIBODY PRODUCTION
Immunoglobulins are fundamental proteins for the adaptive immune response and are valuable tools in biomedical research. Using antibodies, it is possible to mark cells, or components of them, detect, analyze, or purify proteins etc [11]. Some antibodies have also been utilized as a treatment for bacterial infections [8].
Nowadays, the importance of producing antibodies in a short time, and in a non-invasive and cost-effective fashion is recognized [9]. Avian, particularly poultry, immunoglobulins fulfill these requirements. Large amounts of antibodies can be obtained from egg yolks (as much as 100 to 250 mg per egg) and a hen can lay 300 eggs a year [13,36].
Several reports about the characterization of immunological responses and antibody production processes in birds have been published. Erhard et al, carried out many experiments using 972 laying chickens and analyzed 10,692 eggs. They measured serum and yolk antibody concentrations in response to several antigen types and amounts as well as in response to inoculation methodologies [16]. Reviews of these procedures for poultry and wild birds are presented in [14,37]. The primary response (obtained after first inoculation) kinetics has also been characterized for different chicken lines and antigens describing the time to reach maximum IgY levels [38].
However, none of these works or reviews considers booster schedules. The question of when a second antigen dose should be applied, to improve the immune response, has not been answered anywhere. No arguments have been
presented that establish booster times. On the contrary, the schemes that have been published are completely empirical or based on typical immune responses that can differ among different antigens.
4. DYNAMICAL MODEL
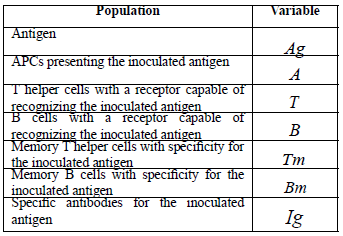
In this section, a mathematical model for avian humoral immune response is presented. This process is described for the cellular and molecular populations that participate in adaptive immune response. These populations are listed in Table 1. Units used are concentrations in cells per milliliter and micrograms per milliliter.
Table 1. Cellular and molecular populations considered in the model.
4.1 Antigen
Equation (1) describes the dynamics of antigen concentration and has three terms. The first one represents the antigen decay due to the effect of the immune system with a rate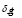 . The second term corresponds to the opsonization process; i.e., the increase of the phagocytes’ activity due to antibody signaling. The antigen elimination rate
. The second term corresponds to the opsonization process; i.e., the increase of the phagocytes’ activity due to antibody signaling. The antigen elimination rate 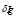 (by antibody effect) and the product of those two populations determine the speed of antigen degradation (mass action effect).
(by antibody effect) and the product of those two populations determine the speed of antigen degradation (mass action effect).
The input 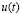 refers to the system input (inoculated antigen concentration). This function represents the event that elicits the immune response and it is the signal that can be manipulated by the designer.
refers to the system input (inoculated antigen concentration). This function represents the event that elicits the immune response and it is the signal that can be manipulated by the designer.
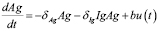 (1)
(1)
4.2 Antigen presenting cells (APC)
APC’s (mainly macrophages and dendritic cells) dynamics is modeled by an equation with three components:
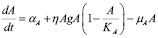 (2)
(2)
This equation refers exclusively to the population of APC that has ingested the inoculated antigen and presents it to T helper cells. A constant production rate of APCs is measured by the 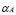 parameter, representing phagocytes generation from marrow bone stem cells. However, this rate corresponds only to mature APCs that are found near the inoculation region, not the circulating ones. Finally, a natural death rate
parameter, representing phagocytes generation from marrow bone stem cells. However, this rate corresponds only to mature APCs that are found near the inoculation region, not the circulating ones. Finally, a natural death rate 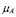 is included.
is included.
The second term represents the increase in APC population due to innate immune response to the specific antigen. Since this effect appears only in the presence of the antigen, a typical mass-action term can be included. However, this allows for an unbounded population size. A sigmoidal 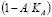
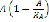 factor has been added to account for two aspects of this stage of the immune response: 1) a progressive increase in the growth rate of APC concentration, and 2) a limited size APC population. The
factor has been added to account for two aspects of this stage of the immune response: 1) a progressive increase in the growth rate of APC concentration, and 2) a limited size APC population. The 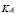
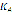 parameter sets the population maximum size and
parameter sets the population maximum size and 
 measures how effectively phagocytes move and ingest the antigen.
measures how effectively phagocytes move and ingest the antigen.
4.3 Active T-helper cells
T lymphocytes have a more complex set of interactions affecting the population growth.
The model described by (3) includes activation, clonal expansion, and immunological memory.
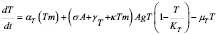 (3)
(3)
The number of specific T cells carrying a receptor capable of recognizing the antigen is kept very low in its absence. The function 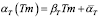 represents a memory cell-dependent production rate. Before exposure to the specific antigen, the memory cell population is zero. New lymphocytes are produced at a constant rate,
represents a memory cell-dependent production rate. Before exposure to the specific antigen, the memory cell population is zero. New lymphocytes are produced at a constant rate, 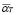 .
. 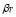 describes the increase in T cell concentration as the result of immunological memory.
describes the increase in T cell concentration as the result of immunological memory. 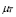 is a death rate.
is a death rate.
T-helper lymphocyte activation initiates the adaptive immune response. This process depends on APC and T cell concentrations as well as on the probability 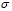 of their encounter. Clonal expansion begins after the activation stage. The coefficient
of their encounter. Clonal expansion begins after the activation stage. The coefficient 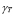 defines the speed of clone generation from activated T lymphocytes. The mass action term of T cells and the antigen is necessary because the expansion persists only while the antigen is present. Finally, the memory effect is represented by a product of antigen concentration, memory T cell population, and the parameter
defines the speed of clone generation from activated T lymphocytes. The mass action term of T cells and the antigen is necessary because the expansion persists only while the antigen is present. Finally, the memory effect is represented by a product of antigen concentration, memory T cell population, and the parameter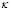 . The latter defines the speed of T cell differentiation.
. The latter defines the speed of T cell differentiation.
A sigmoidal function is included in this equation for the same reasons explained for the APC dynamics description. The maximum population of T lymphocytes is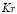 .
.
4.4 Active B cells (Plasma cells)
The cells that secrete antibodies are modeled in this section. An equation is used for the dynamical system description that is, basically, the same as the one used for T lymphocytes. In this case,  describes the effect of memory B cells. The other terms have a similar role as in the T cell case, with corresponding parameters bearing the B subscript.
describes the effect of memory B cells. The other terms have a similar role as in the T cell case, with corresponding parameters bearing the B subscript.
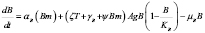 (4)
(4)
All processes associated with B cells are analogous to the T cell case, except for the first term. This represents the activation of B cells as the result of the recognition of the antigen by its receptor and a simultaneous signal received from a T-helper cell. In this case, 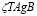 is the result of such an activation where
is the result of such an activation where 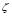 measures the probability that it will occur. The plasma cell kinetics exhibits a behavior fully compatible with a sigmoidal function. That is the reason for the inclusion of such a term in the equation.
measures the probability that it will occur. The plasma cell kinetics exhibits a behavior fully compatible with a sigmoidal function. That is the reason for the inclusion of such a term in the equation.
4.5 Memory cells
The equations modeling the dynamics of both memory lymphocytes are similar:
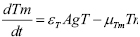 (5)
(5)
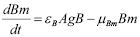 (6)
(6)
Memory cell generation is the result of the activation of T and B cell activation. A mass action term is included to account for the cessation of memory cell generation after the disappearing of the antigen exposure. The proportion of cells that upon activation become memory cells are given by the parameters  and
and 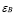 . These cells have long life expectations, characterized by a small
. These cells have long life expectations, characterized by a small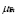 and
and 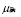 .
.
4.6 Immunoglobulins
IgYs are transferred from the blood to the yolk during egg formation. The levels of serum and yolk antibodies take around a week to reach equilibrium [17]. This means that only one equation is required for both types of antibodies.
Equation (7) presents the serum antibody dynamics. Plasma cells secrete those soluble proteins, hence antibody generation is directly related to the size of the B cell population, with a constant rate 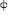
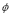 . The parameter
. The parameter 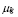 is the rate of natural decay of Igs. Thus, the model is:
is the rate of natural decay of Igs. Thus, the model is:
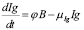 (7)
(7)
5. PARAMETERS OF THE MODEL
The parameters’ values must be obtained from experiments. However, initial values are required for all parameter estimation algorithms. These were obtained from experimental results reported in the literature.
5.1 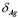 [days
[days ]
]
A half life for the antigen in mammals of a few days is proposed in [39]. Similar results have been obtained experimentally for birds. For example, the kinetic response of birds to sparfloxacin has been reported and this antigen showed a half life of one day [40]. Responses to different antigens can also vary due to the degree of contribution of the liver and the kidney to their elimination. We use ( =0.5).
=0.5).
5.2 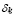 [
[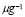 days
days ]
]
Generation of specific antibodies increases the capacity of phagocytes to internalize (i.e. digest) the antigen. Qureshi [32] found that the number of these cells in a culture of chicken macrophages increased 50-90% in the presence of antibodies. From the dynamics of antibodies and using the relation described above:
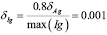 (8)
(8)
It is assumed that the maximum level of Igs in serum, is reached after two weeks and it is about 1000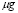 /ml.
/ml.
5.3 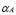 [Cell days
[Cell days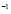 ] and
] and 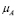 [days
[days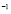 ]
]
Prior to inoculation (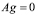 ), the cell population is at an equilibrium
), the cell population is at an equilibrium 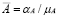 . The half life of APCs, in particular Dendritic Cells (DC), has been determined experimentally [41]. For human DCs, for instance, half life is about a few days [41]. We chose a value of four days, giving
. The half life of APCs, in particular Dendritic Cells (DC), has been determined experimentally [41]. For human DCs, for instance, half life is about a few days [41]. We chose a value of four days, giving 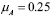 . The dynamics of APCs models only the concentration of cells presenting the antigen of interest. Before the inoculation there should be no such cells. However, we will assume that a single cell is present, so that the process can be initiated. This gives (
. The dynamics of APCs models only the concentration of cells presenting the antigen of interest. Before the inoculation there should be no such cells. However, we will assume that a single cell is present, so that the process can be initiated. This gives (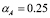 ).
).
5.4 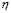 [
[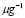 days
days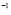 ] and
] and 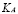 [Cell]
[Cell]
The speed of antigen internalization by DCs has been estimated experimentally with a value of about three hours [42]. Therefore, the rate of change of the population is determined mainly by the time required for cell migration and antigen recognition. 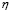 was found through simulation so that the peak level of antigen presentation occurs between six and twelve hours after inoculation [2]. Thus (
was found through simulation so that the peak level of antigen presentation occurs between six and twelve hours after inoculation [2]. Thus (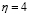 ).
).
The peak value reached by the population is not easily found in the literature because this model considers only those cells that have phagocytosed the antigen and presented it to T-cells. This varies according to the antigen being used. We used 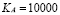 for the simulations.
for the simulations.
5.5 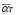 ,
, 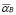 [Cell days
[Cell days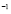 ],
], 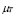 and
and 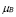 [days
[days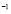 ]
]
At first, there is a small population of lymphocytes (T-helper and B) that have a receptor for a peptide resulting from antigen processing by APCs (or antigen segment). It is estimated that in hens there are 10-100 cells from a total cell population of 100 million [32]. This is about one cell in one ml of blood.
Life expectancy for chicken B-lymphocytes is about three days (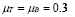 ). According to the model, the steady state cell population is
). According to the model, the steady state cell population is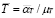 . Therefore,
. Therefore,
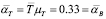 (9)
(9)
5.6 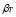 and
and 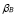 [days
[days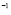 ]
]
After the immunization, the number of T cells reactive for a given antigen increases significantly, from 100 to 1000 times the initial number [5]. Based on this range and on the dynamics of T-cells 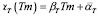 :
:
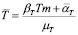 (10)
(10)
Furthermore, it is assumed that the secondary response is mainly made out of memory cells. Hence, 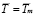 . This gives
. This gives 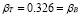 .
.
5.7 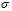 [Cell
[Cell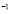 days
days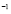 ]
]
A single dendritic cell can activate 10 to 20 cells per day [44]. Choosing the lower value gives 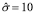 . This value, however, requires a correction according to the two terms (A and Ag) appearing in the equation. For an initial concentration of antigen concentration of 10
. This value, however, requires a correction according to the two terms (A and Ag) appearing in the equation. For an initial concentration of antigen concentration of 10 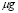 /ml and a maximum of 10,000 cells,
/ml and a maximum of 10,000 cells, 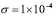 .
.
5.8 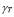 and
and 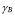 [
[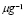 days
days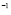 ]
]
Usually, the peak of lymphocyte population due to the adaptive response occurs around a week (seven days) after inoculation. It will be assumed that, in the primary response, clonal expansion is responsible for most of the specific T-cell population. It will also be assumed that the initial cell population is just one cell (in accordance with the initial concentration of lymphocytes having a receptor for the antigen in use) and that the population reaches 99% of the peak value within seven days. Therefore, from the solution of the logistic equation,
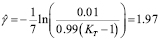 (11)
(11)
Adjusting for the amount of antigen concentration units 10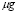 /ml:
/ml: 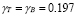
5.9 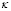 ,
, 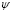 [Cell
[Cell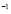 days
days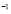 ] and
] and 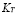 [Cel]
[Cel]
Naïve T-cells in mammals require a stimulation period between 6 and 30 hours before starting clonal expansion. Memory cells, on the other hand, respond in 0.5 to 2 hours after stimuli [41]. This effect has been modeled using three terms in the ODE, and parameter values were obtained through in silico experimentation. A plausible result was obtained using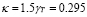 . By an argument similar to that used for T-cell memory,
. By an argument similar to that used for T-cell memory, 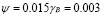 was chosen.
was chosen.
T-helper cell expansion is known to be in the range of 100 to 1000 [2]. 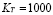 was chosen.
was chosen.
5.10 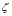 [
[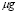 Cell
Cell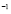 days
days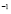 ]
]
The kinetics of B lymphocyte activation by T-helper cells has not been reported in the literature, to the authors’ knowledge. Due to the similarity of the dynamical equations, we will use the same activation parameter as that used for T lymphocytes, adjusted for taking into account the maximum population size, i.e., 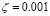 .
.
5.11 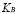 [Cell]
[Cell]
Effector T lymphocytes reach a peak value of around 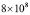 cells per ml [5]. This order of magnitude and the antibody production rate agree with experimental results in birds. Therefore,
cells per ml [5]. This order of magnitude and the antibody production rate agree with experimental results in birds. Therefore, 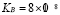 will be used.
will be used.
5.12 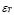 y
y 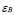 [
[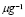 days
days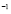 ]
]
In mammals, about 5% of effector T-cells remain as memory cells in circulation [45]. Thus, it will be assumed that the rate of memory lymphocyte production is 5% of clonal expansion. This value, as in the clonal expansion of naïve cells, is adjusted for the initial effect of the antigen concentration:
 (12)
(12)
5.13 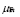 y
y 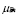 [days
[days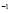 ]
]
The number of memory cells for a given antigen is severely limited and remains almost constant during the memory phase [5]. The life expectancy has been estimated to be several months, even years, in mammals [43]. In our model we will use (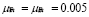 ).
).
5.14  [
[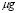 Cell
Cell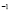 days
days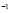 ]
]
The rate of antibody secretion has been estimated, for mammals, to be in the range of 8,000 to 20,000 molecules per second in a differentiated B cell [39]. We will choose the same value used by Oprea [39] in his work, i.e., 10,000 molecules per second. Adjusting for change of units (secs to days, and Daltons to kgs: 1D = 1.66053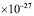 kg). Since a hen’s IgY weighs 180kDa [9]:
kg). Since a hen’s IgY weighs 180kDa [9]:
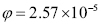 (14)
(14)
5.15 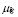 [days
[days ]
]
The half life of IgYs in serum for 2-9 month old hens was measured in [32] to be 3.3 days. This gives (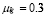 ).
).
6. SIMULATION RESULTS
The time response of the model was simulated, using Matlab® and the results were qualitatively compared to experimental results reported in the literature. An initial antigen concentration of 10 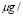 ml was used. This represents the typical value of concentration found in the blood after inoculation [37,40].
ml was used. This represents the typical value of concentration found in the blood after inoculation [37,40].
Figure 1 shows the primary response in terms of the concentration of specific antibodies in serum. The arrow marks the time of inoculation. Antibody production starts after five days. This is consistent with the presentation, expansion, and differentiation periods of lymphocytes. The peak value occurs about a week later. For qualitative comparison purposes, Figure 2 illustrates the responses as measured by Parmentier et al. [46], see Figure 1, page 250. The vertical axis represents the concentration in titers (a base two logarithmic scale).
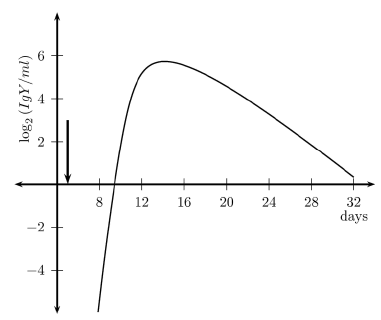
Figure 1. Antibody concentration IgY (Primary response)
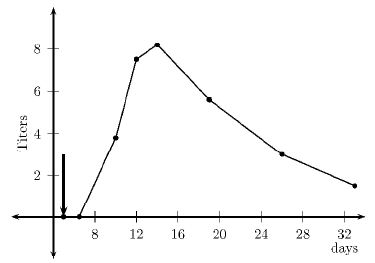
Figure 2. Serum antibody concentration (Primary response [46])
The results of the simulation of secondary response are shown on Figure 3. It can be seen that secondary response is both faster and more intense. An important fact is that this response consists mainly of G-type antibodies compared to the basically IgM-formed primary response. The ratio of the peak values is between 1,000 and 10,000 as shown in the curve. The effect of memory is also evidenced by a shortened time between stimuli and peak immune response.
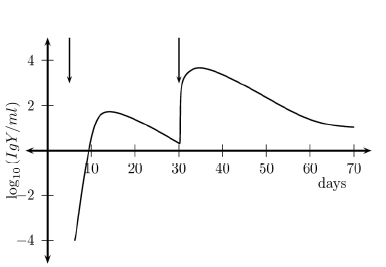
Figure 3. Antibody concentration IgY (Primary and secondary response)
7. CONCLUSIONS
A dynamical model consisting of seven nonlinear ordinary differential equations was introduced to represent the avian immune response to antigens. The derivation of the model was based on the current knowledge of the behavior of the immune system in mammals and birds. The resulting simulated response shows good qualitative agreement with experimental data in published works [2, 46].
Parameter values were chosen based on experimental results published and on simulation results. The concentration of antibodies obtained from simulations was contrasted to experimental kinetics reported by several researchers with good agreement. Full model validation will require the results of experimental immune response of chickens to a specific antigen.
The experimental results published to date show great variability in the response time and the peak value reached. Therefore, it is very important to establish parameter tuning strategies before using the model for a bird lineage and a specific antigen. Currently, experimental work is under way to allow for the validation of the model.
ACKNOWLEDGEMENTS
This work has been supported in part by the Direccion de Investigación, Sede Bogotá (under Grant DIB: 8003271) and by the Vice-Dean of Research and the Department of Electrical Engineering, of the School of Engineering, of the National University of Colombia.
REFERENCES
[1] AMEMIYA CHRIS, SAHA T., NIL RATAN AND ZAPATA AGUSTIN. Evolution and development of immunological structures in the lamprey. 2007, Current Opinion in Immunology, Vol. 19, pp. 535-541. [ Links ]
[2] ABBAS ABUL K. AND LICHTMAM ANDREW H. Basic Immunology Functions and disorders of the Immune System. Saunders 2004. [ Links ]
[3] BENDELAC ALBERT, SAVAGE PAUL B. AND TEYTON, LUC. The Biology of NK T Cells 2007, Annu. Rev. Immunol., Vol. 25, pp. 297-336. [ Links ]
[4] SERBINA NATALYA V., et al Monocyte mediated defense against microbial pathogen. 2008, Annu. Rev. Immunol., Vol. 26, pp. 421-452. [ Links ]
[5] JANEWAY C. et al. Immunobiology. 6th. Garland Science, 2005. [ Links ]
[6] PERELSON ALAN S. AND WEISBUCH GERARD. Immunology for physicists. 1997, Reviews of Modern Physics, Vol. 69, pp. 1219-1267. [ Links ]
[7] MAYNARD JENNIFER AND GEORGIOU GEORGE. Antibody Engineering. 2000, Annu. Rev. Biomed. Eng., Vol. 2, pp. 339-376. [ Links ]
[8] BERGHMAN L. R., et al. Antibodies: An Alternative for Antibiotics? 2005, Poultry Science, Vol. 84, pp. 660-666. [ Links ]
[9] COVA LUCYNA. DNA-designed avian IgY antibodies: novel tools for research, diagnostics and therapy. 2005, Journal of Clinical Virology, Vol. 34 Suppl.1, pp. 70-74. [ Links ]
[10] TINI M. et al. Generation and application of chicken egg-yolk antibodies. 2002, Comp Biochem Physiol A Mol Integr Physiol, Vol. 13, pp. 569-574. [ Links ]
[11] HARLOW ED AND LANE DAVID. Antibodies a Laboratory Manual. 1988. [ Links ]
[12] HAMAL K. R. et al. Maternal Antibody Transfer from Dams to Their Egg Yolks, Egg Whites, and Chicks in Meat Lines of Chickens. 2006, Poultry Science, Vol. 85, pp. 1364-1372. [ Links ]
[13] IVARIE ROBERT. Competitive bioreactor hens on the horizon. 2006, Trends in Biotechnology, Vol. 24, pp. 99-101. [ Links ]
[14] SCHADE R. et al. Chicken Egg Yolk Antibodies, Production and Application: IgY-Technology (Springer Lab Manuals). Springer, 2000. [ Links ]
[15] ERHARD MICHAEL H. et al. Lipopeptide Pam3Cys-Ser-(Lys)4: An Alternative Adjuvant to Freund's Adjuvant for the Immunisation of Chicken to Produce Egg Yolk Antibodies. 1997, ATLA, Vol. 25, pp. 173-181. [ Links ]
[16] ERHARD MICHAEL H. et al. Evaluation of Various Immunisation Procedures in Laying Hens to Induce High Amounts of Specific Egg Yolk Antibodies 2000, ATLA, Vol. 28 pp.63-80. [ Links ]
[17] HAU J. AND HENDRIKSEN C. F. Refinement of polyclonal antibody production by combining oral immunization of chickens with harvest of antibodies from the egg yolk. 2005, Ila[18] MURRAY J.D. Mathematical Biology. Springer, 2002. Vol. 1. [ Links ] [ Links ]
[18] MURRAY J.D. Mathematical Biology. Springer, 2002. Vol. 1.
[19] NOWAK MARTIN AND MAY ROBERT Virus Dynamics. Oxford University Press, 2004. [ Links ]
[20] SCHNELL SANTIAGO, GRIMA RAMON AND MAINI PHILIP Multiscale Modeling in Biology. 2007, American Scientist, Vol. 95, pp. 134-142. [ Links ]
[21] DE BOER, ROB. Theoretical Immunology. Available: http://www-binf.bio.uu.nl/rdb/books /ti.pdf, [citado Agosto de 2010]. [ Links ]
[22] ASQUITH BECCA AND BANGHAM CHARLES The dynamics of T-cell fratricide: application of a robust approach to mathematical modelling in immunology. 2003, Journal of Theoretical Biology, Vol. 222, pp. 53-69. [ Links ]
[23] DE BOER ROB J. Mathematical models of human CD4+ T-cell population kinetics. 2002, Netherlands Journal of Medicine, Vol. 60, pp. 17-26. [ Links ]
[24] CARNEIRO JORGE et al. A Model of the Immune Network with B-T Cell Cooperation I Prototypical Structures and Dynamics. 1996, J. theor. Biol, Vol. 182, pp. 513-529. [ Links ]
[25] FLORES LUIS E. et al. A graph model for the evolution of specificity in humoral immunity. 2004, Journal of Theoretical Biology, Vol. 229, pp. 311-325. [ Links ]
[26] KESMIR CAN AND DE BOER ROB J. A Mathematical Model on Germinal Center Kinetics and Termination. 1999, The Journal of Immunology, Vol. 163, pp. 2463-2469. [ Links ]
[27] KLEINSTEIN STEVEN AND SEIDEN PHILIP. Simulating the Immune System. 2000, Computing in Science & Engineering, Vol. 2, pp. 69-77. [ Links ]
[28] MO HONGWEI, GUO MAOZU AND XU LIFANG. Simulating the Mutation of Antibody of Human Immune System. 2005, Neural Networks and Brain ICNN &B, Vol. 3, pp. 1415-1419. [ Links ]
[29] PARMENTIER HENK K. et al. Differences in distribution of lymphocyte antigens in chicken lines divergently selected for antibody responses to sheep red blood cells. 1995, Vet. Immunology and Immunopathology, Vol. 48, pp. 155-168. [ Links ]
[30] SHARMA J. AND TIZARD I. Avian Cellular Immune Effector Mechanisms: A Review. 1984, Avian Pathology, Vol. 13, pp. 357-376. [ Links ]
[31] ERF, G. F. Cell-Mediated Immunity in Poultry. 2004, Poultry Science, Vol. 83, pp. 580-590. [ Links ]
[32] DAVISON F., KASPERS B. AND SCHAT K. Avian Immunology. Academic Press, 2008. [ Links ]
[33] HAMMER D. K. The Immune System in Chickens. 1974, Avian Pathology, Vol. 3, pp. 65-78. [ Links ]
[34] FAIRBROTHER, ANNE, SMITS, JUDIT AND GRASMAN, KEITH A. Avian Immunotoxicology. 2004, Journal of Toxicology and Environmental Health, Part B, Vol. 7, pp. 105-137. [ Links ]
[35] QURESHI MUQUARRAB, HEGGEN CHERILYN L. AND HUSSAIN IRSHAD. Avian macrophage: effector functions in health and disease. 2000, Developmental and Comparative Immunology, Vol. 24, pp. 103-119. [ Links ]
[36] GROSSMAN M. AND KOOPS W. J. A Model for Individual Egg Production in Chickens. 2001, Poultry Science, Vol. 80, pp. 859-867. [ Links ]
[37] SNOEIJS TINNE et al. Kinetics of primary antibody responses to sheep red blood cells in birds: a literature review and new data from great tits and European starlings. 2007, Animal Biology, Vol. 57, pp. 79-95. [ Links ]
[38] WEIGEND S., MIELENZ N. AND LAMONT S. Application of a Nonlinear Regression Function to Evaluate the Kinetics of Antibody Response to Vaccines in Chicken Lines Divergently Selected for Multitrait Immune Response. 1997, Poultry Science, Vol. 76, pp. 1248-1255. [ Links ]
[39] OPREA MIHAELA AND PERELSON A. Exploring the Mechanisms of Primary Antibody Responses to T Cell Dependent Antigens 1996, J.theor.Biol., Vol. 181, pp. 215-236. [ Links ]
[40] SRIRANJANI D. et al. Pharmacokinetics of sparfloxacin in broiler chicken. 2006, British Poultry Science, Vol. 47, pp. 720-725. [ Links ]
[41] SALLUSTO F. AND LANZAVECCHIA A. The instructive role of dendritic cells on T-cell responses. 2002, Arthritis Res, Vol. 4, pp. 127-132. [ Links ]
[42] INABA KAYO et al. Dendritic Cells Pulsed with Protein Antigens In Vitro Can Prime Antigen-specific, MHC-restricted T Cells In Situ. 1990, J. Exp. Med., Vol. 172, pp. 631-640. [ Links ]
[43] SPRENT JONATHAN AND TOUGH DAVID. Lymphocyte Life-Span and Memory. 1994, Science, Vol. 265, pp. 1395-1400. [ Links ]
[44] ROMANI N. et al. A small number of anti-CD3 molecules on dendritic cells stimulate DNA synthesis in mouse T lymphocytes. 1989, J. Exp. Med, Vol. 169, pp. 1153-1168. [ Links ]
[45] WILLIAMS MATTHEW A. AND BEVAN MICHAEL J. Effector and Memory CTL Differentiation. 2007, Annu. Rev. Immunol., Vol. 25, pp. 171-192. [ Links ]
[46] PARMENTIER HENK AND MECHTELD WALRAVEN. Antibody Responses and Body Weights of Chicken Lines Selected for High and Low Humoral Responsiveness to Sheep Red Blood Cells. Poultry Science, 1998, Vol. 77, pp. 248-255. [ Links ]

