










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkDYNA
versão impressa ISSN 0012-7353
Dyna rev.fac.nac.minas vol.79 no.175 Medellín set./out. 2012
PRODUCCIÓN DE ACIDO CLAVULÁNICO POR FERMENTACIÓN DE Streptomyces clavuligerus: EVALUACIÓN DE DIFERENTES MEDIOS DE CULTIVO Y MODELADO MATEMÁTICO
PRODUCTION OF CLAVULANIC ACID BY FERMENTATION OF Streptomyces clavuligerus: EVALUATION OF DIFFERENT CULTURE MEDIA AND MATHEMATICAL MODELING
CLAUDIA PATRICIA SANCHEZ HENAO
Magister en. Ing. Química, Universidad de Antioquia, profesora Ingeniería de Alimentos, grupo de Bioprocesos, clapatriciasa@gmail.com
NATHALIA ANDREA GOMEZ GRIMALDOS
Ing. Química, Universidad de Antioquia, Corporación Académica Ambiental, natgogri@gmail.com
JUAN CARLOS QUINTERO DIAZ
Doctor en Ing. Química. Universidad de Antioquia, Profesor Asociado, grupo Bioprocesos, jcquinte@udea.edu.co
Recibido para revisar Marzo 5 de 2012, aceptado Mayo 14 de 2012, versión final Junio 19 de 2012
RESUMEN: El ácido clavulánico (AC) es un antibiótico b-lactámico con una potente capacidad inhibidora de la actividad b-lactamasa. En el presente trabajo se evaluaron cuatro medios de cultivo reportados en la bibliografía y uno propuesto por los autores, para la producción de AC. Se evaluó también la concentración de glicerol adecuada para el medio seleccionado y con base en los resultados obtenidos se realizó una evaluación de la producción de AC en un biorreactor de laboratorio. Se propuso un modelo matemático y se evaluó su ajuste a los resultados experimentales. Se observó que los medios complejos con presencia de una mezcla de aminoácidos libres, son adecuados para la producción de AC. El glicerol presentó un valor óptimo frente a la producción de AC en 50 g.L-1 y se observó el efecto inhibitorio a concentraciones de 100 g.L-1. Con el medio complejo y 50 g.L-1 de glicerol, la concentración de AC alcanzada fue de 994 mg.L-1. El modelo matemático propuesto que incluye una cinética tipo Contois para la biomasa y una cinética de formación de producto parcialmente asociado al crecimiento, presentan un ajuste significativo con un 95% de nivel de confianza.
PALABRAS CLAVE: Acido clavulnico, harina de soja, glicerol, modelo cinético, modelación
ABSTRACT: Clavulanic acid (CA) is a b-lactam antibiotic exhibiting potent inhibitory capacity towards b-lactamases. In this paper we assessed the production of CA in four culture media reported in the literature compared to a novel culture medium. The results of lab-scale bioreactor studies were used to establish the optimum glycerol concentration in the selected media. It was observed that the presence of complex media with a mixture of free amino acids, are suitable for the production of CA. Glycerol showed an optimum value compared to the production of CA in 50 g.L-1 and inhibitory effect was observed at concentrations of 100 g.L-1. With the complex medium and 50 g.L-1 of glycerol, the concentration of CA achieved was 994 mg.L-1. A mathematical model was proposed and their fit to the experimental results was evaluated. The proposed mathematical model includes a type Contois kinetics for biomass and product formation kinetics partially associated with growth, and showed a significant adjustment with 95% confidence level.
KEY WORDS: clavulanic acid, flour, glycerol, kinetic model, modelation
1. INTRODUCCIÓN
El primer antibiótico microbiano, la penicilina, fue descubierto en 1928 por Sir Alexander Fleming, pero fue hasta 1939 que se pudo usar en el tratamiento de enfermedades infecciosas. Desde entonces y hasta la actualidad, se han desarrollado tecnologías de producción de gran cantidad de antibióticos, que para el 2009 generaron un mercado global de 42 billones de dólares [1]. El desarrollo de resistencia microbiana a los antibióticos está directamente relacionado con su grado de exposición a ellos, y puesto que los antibióticos b-lactámicos (ej. Penicilinas, cefalosporinas, etc.) contribuyen al 40% del mercado total de antibióticos [2], se ha observado que los mecanismos de resistencia bacteriana a antibióticos b-lactámicos representan actualmente el principal problema clínico a nivel mundial. La producción de enzimas b-lactamasas es uno de los principales mecanismos de resistencia bacteriano a estos antibióticos. Estas enzimas destruyen la molécula de antibiótico y lo tornan inactivo [3].
El ácido clavulánico (AC), es una molécula que exhibe una potente actividad inhibitoria de las enzimas b-lactamasas, por lo que se ha convertido en un importante producto comercial disponible en combinaciones con antibióticos b-lactámicos como amoxicilina (Augmentin TM), tetraciclina (Timentín TM) [4] y también como producto genérico. Su efecto inhibidor fue descubierto en 1976 y desde entonces se han realizado exhaustivos estudios con el fin de mejorar los títulos de antibiótico empleando procesos de fermentación con la bacteria que naturalmente lo produce, Streptomyces clavuligerus (Sc).
La productividad del proceso, depende de muchos factores, entre ellos del tipo y concentración de nutrientes, de las condiciones de operación de la fermentación y de los propios mecanismos intracelulares de biosíntesis. A pesar de los esfuerzos de los investigadores que trabajan en el tema alrededor del mundo, aún no se ha conseguido obtener valores de producción (titulo) de AC superiores a 1g.L-1 [5, 6], e incluso en la gran mayoría de trabajos y después de estudios de optimización estos valores han sido inferiores a 500 mg L-1 [7-9]. De acuerdo con información de patentes registradas a nombre de importantes empresas farmacéuticas, se han reportado valores de producción de AC entre 561 y 1000 mg L-1 [10, 11].
Por tanto, es de gran importancia continuar abordando estudios tendientes a mejorar la producción del AC, lo que permitirá reducir sus costos y hacerlo más accesible a la población de escasos recursos.
En diferentes estudios publicados, se ha observado el empleo de una gran variedad de formulaciones de medios para el cultivo de S c [5, 10-15]. Algunos de ellos son químicamente definidos y otros son medios complejos. Estos últimos favorecen la producción de AC puesto que la bacteria requiere para su biosíntesis la presencia de aminoácidos [16], los cuales están presentes en sustratos complejos como harina de cereales o digeridos de proteínas, que presentan la ventaja de ser nutrientes de bajo costo. Sin embargo, debido a la complejidad del medio y la baja concentración del producto, su purificación se torna compleja y costosa [11, 17]. Los carbohidratos son la fuente de carbono y energía preferida, por ser las más simples, para el crecimiento y producción de metabolitos secundarios; pero el Sc presenta una inusual incapacidad de consumir glucosa y otros carbohidratos simples, su principal fuente de carbono son los lípidos [18, 19, 20]. En estas formulaciones se emplea como fuente de carbono almidón o glicerol [20, 21], sin embargo, dado que el glicerol ha sido identificado como el precursor de 3 átomos de carbono para la síntesis de AC [6, 21], en la gran mayoría de trabajos se emplea esta sustancia como la fuente de carbono. Sc es incapaz de consumir glucosa y otros azúcares simples.
Debido a la gran heterogeneidad en la composición de los medios de cultivo reportados en la literatura para la producción de AC, los cuales han sido desarrollados y evaluados, no siempre a las mismas condiciones, se hace necesario evaluarlos bajo las mismas condiciones y con la misma especie bacteriana, con el fin de determinar la verdadera capacidad de estos medios para la producción de AC.
En este trabajo se evaluaron 5 diferentes composiciones de medios de cultivo para producción de AC bajo las mismas condiciones de cultivo de S. clavuligerus. Una vez seleccionado el medio de mayor producción se evaluaron diferentes concentraciones de la fuente de carbono, y el medio seleccionado se empleó para producir AC en un biorreactor a escala de laboratorio. También se propuso un modelo matemático para describir el comportamiento cinético de la fermentación.
2. MATERIALES Y MÉTODOS
2. 1. Técnicas microbiológicas
2.1.1. Microorganismo y medio de conservación.
Se empleó un liofilizado de la cepa de Streptomyces clavuligerus ATCC 27064. La activación se realizó en caldo TSB® (Tryptic Soy Broth) a 28 ºC por 36 h. La cepa activada se almacenó a -80ºC en tubos Eppendorf con medio TSB® y glicerol al 40%. Estos tubos se emplearon como semilla para todos los ensayos [22].
2.1.2. Medios de cultivo
Los inóculos empleados para cada ensayo se prepararon mezclando en Erlenmeyers de 500 mL, el contenido de un tubo semilla con 100 mL de medio TSB® , e incubándolos a 28 ºC y 220 rpm por 36 h. A estas condiciones, la DO600nm obtenida osciló entre 0,6 y 0,8[22].
Los medios de cultivo utilizados para evaluar la producción de AC fueron: Medio 1: Almidón (10 gL-1), L-asparagina (2 gL-1), MgSO4.7H2O (0,6 gL-1), K2HPO4 (4,4 gL-1), FeSO4.7H2O (1 gL-1), MnCl2.4H2O (1mgL-1), ZnSO4.7H2O (1 mgL-1) y CaCl2.3H2O (1,3 mgL-1). El medio se reguló con MOPS (100 mM) y se ajustó a pH 6,9 [5]. Medio 2: Es una modificación del medio 1, empleando en lugar de almidón, glicerol a una concentración de 20 gL-1 [23]. Medio 3: glicerol (10 gL-1), harina de soja (40 gL-1), aceite de soja (16 gL-1), K2HPO4 (1,2 gL-1), MnCl2.4H2O (1 mgL-1), ZnSO4.7H2O (1mgL-1) y CaCl2.3H2O (1,3 mgL-1). El medio se reguló con MOPS (100 mM) y se ajustó a pH 6,8 [12]. Medio 4: glicerol (20 gL-1), harina de soja (5,5 gL-1) y K2HPO4 (0,8 gL-1). El medio se reguló con MOPS (100 mM) y se ajustó a pH 7,0 [13]. Medio 5: glicerol (20 gL-1), harina de soja (15 gL-1) y casaminoácidos (5 gL-1). El medio se reguló con MOPS (100 mM) y se ajustó a pH 7,0 [24] . Antes de su inoculación, los medios de cultivo se esterilizaron en autoclave a 121 ºC durante 20 min.
2.1.3. Métodos experimentales
Se realizaron cinco cultivos de fermentación por triplicado en Erlenmeyers de 500 mL. A cada frasco se adicionó 100 mL del medio de cultivo a evaluar y la biomasa contenida en 10 mL de inóculo. Esta biomasa se obtuvo centrifugando 10 mL de inóculo a 5000 rpm por 10 min., eliminando el sobrenadante y resuspendiendo la biomasa con 10 mL de medio de prueba del Erlenmeyer y retornándolo nuevamente al Erlenmeyer. Los frascos se sellaron con tapones de algodón para permitir la difusión del oxígeno al cultivo y se incubaron a 28 ºC y 220 rpm por 36 h.
Con el medio de cultivo seleccionado, se evaluaron tres concentraciones de glicerol: 20 gL-1 (2%), 50 gL-1 (5%) y 100 gL-1 (10%). Los cultivos se realizaron por triplicado bajo las mismas condiciones operacionales ya descritas.
Para ambos ensayos, al final del periodo de incubación se tomaron muestras para cuantificar la concentración de biomasa y de AC. Los valores medios de cada ensayo se compararon estadísticamente mediante la prueba de rangos múltiples de Duncan, con el fin de determinar las diferencias significativas entre ellas [25,26].
La producción de AC se evaluó por duplicado en un biorreactor de laboratorio de 5 L (Biostat B Braun), con un volumen de trabajo de 3 L e inóculo al 10% v/v. La fermentación se llevó a cabo a 28ºC, 400 rpm y un flujo de aire de 1 vvm. El pH se controló en 6,8 +/- 0,2 con soluciones de NaOH (2N) y HCL (2N) adicionadas automáticamente. La espuma se controló adicionando 2 mL de antiespumante (antifoam 204) al inicio de la fermentación. El cultivo se desarrolló por 144 h y se tomaron muestras periódicamente para determinar las concentraciones de biomasa, glicerol y AC.
2.2. Métodos analíticos
Las muestras de la fermentación se centrifugaron a 14000 rpm por 10 min. a 4ºC. Los pellets se emplearon para determinar la concentración de biomasa y los sobrenadantes se diluyeron con solución de MOPS (pH 6,8 y 21 mM) y se filtraron a través de membranas de PTFE de 0.2 mm previo al análisis de AC y glicerol [22].
2.2.1. Determinación de biomasa: El micelio se lavó dos veces con 10 mL de solución salina (NaCl 0,9%) y su biomasa se determinó indirectamente mediante la técnica colorimétrica de DNA, basada en lectura de la absorbancia a 600 nm del color azul generado por la reacción de la difenilamina con la desoxirribosa [22, 27].
2.2.2. Determinación de ácido clavulánico: La concentración de AC se determinó por HPLC empleando el método descrito por Foulstone and Reading (1982) [28]. El AC presente en cada muestra, fue derivatizado con imidazol y analizado en un HPLC Agilent/Hewlett Packard Chemstation 1100, operado con columna C18 m-Bondapack fase reversa (Waters), una fase móvil compuesta por una solución de 94% v/v de KH2PO4 (50mM, pH 3,2) y 6% v/v de metanol, aun flujo de 1ml/min y un detector de arreglo de diodos. El AC derivatizado se detectó a 311 nm.
2.2.3. Determinación de glicerol: La concentración de glicerol se determinó por espectrofotometría a 412 nm, basado en la oxidación de ácido perclórico [16].
2.3. Modelo matemático
El modelo propuesto para el proceso de producción de AC a partir de Sc, se obtiene realizando balances de masa en un sistema por lote para la biomasa, el sustrato y el producto. Para la velocidad de crecimiento específica se propuso el modelo de Contois (1). Se ha observado que en cultivos de Sc la viscosidad del caldo se incrementa con el aumento de la concentración de biomasa [29], la viscosidad genera reducción en la transferencia de masa y esto puede limitar la velocidad de crecimiento. Estos fenómenos son representados por la cinética tipo Contois, en la que la velocidad específica de crecimiento es inversamente proporcional a la biomasa. La velocidad de crecimiento microbiano se representa por (2). La velocidad de consumo de glicerol se representa como una función directa de la velocidad de crecimiento microbiano (3) y la velocidad de producción de AC se representa como una función aditiva de la velocidad de crecimiento microbiano y la concentración de biomasa (4) es decir, se representa como un metabolito producido con un aporte del metabolismo primario y un aporte del metabolismo secundario. Adicionalmente, se ha observado que durante las primeras horas de cultivo, el AC no se produce, y a partir de un tiempo determinado cuando ya ha iniciado el crecimiento exponencial, éste se detecta en el medio. Este fenómeno se representa con la función de paso w (t), siendo cero desde el inicio del cultivo hasta un tiempo to y para tiempos superiores a to, la función toma un valor de 1.
La estimación de los parámetros del modelo cinético y la solución numérica de las ecuaciones diferenciales que representan el modelo, se realizó empleando el método de optimización no lineal lsqnonline, basado en el algoritmo de Levenberg-Marquardt, asociado con la herramienta ODE45 de Matlab 2008b.
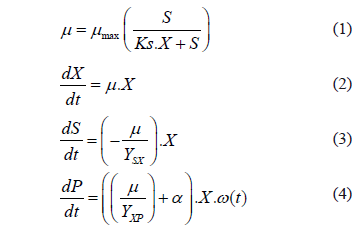
Donde X es la concentración de la biomasa (gL-1), S es la concentración de glicerol (gL-1), P es la concentración de AC (mgL-1), m es la velocidad de crecimiento específica (h-1), mmax es la velocidad de crecimiento máxima específica (h-1), KS es la constante de saturación de Monod (gL-1), YXS es el rendimiento biomasa en glicerol, YXP es el rendimiento de biomasa en AC(gmg-1), a que es una productividad de AC específica (mgg-1h-1) y w(t) es una "función de paso" que asume un valor de 0 en los rangos de tiempo entre 0-48 h y el valor de 1 en el rango de tiempo entre 48-136 h.
3. RESULTADOS Y DISCUSIÓN
3.1. Evaluación de los medios de cultivo
Los resultados de la evaluación de los cinco medios de cultivo se presentan en la Figura 1. Se determinó la concentración de AC y el rendimiento de producto en biomasa como variables para seleccionar el mejor medio. Al realizar un análisis estadístico empleando el test de rangos múltiples de Duncan se observó que hay diferencia significativa entre todos los medios evaluados. Los medios sintéticos (medios 1 y 2) presentan una baja producción de AC y un bajo rendimiento en producto. Igual comportamiento se observó en el medio 3 aunque en este caso el medio contiene harina de soja. El medio 4 presentó un alto rendimiento de AC pero una baja concentración durante el cultivo. El medio de cultivo que reportó los más altos valores de producción de AC fue el medio 5 con 335 mg mgL-1 mientras que con los medios químicamente definidos se alcanzaron valores de 140 mgL-1.
De acuerdo con resultados de la bibliografía, la producción de AC con los medios químicamente definidos se encuentran entre 5,6 y 25,4 mg.L-1 [5], mientras que con medios complejos se han alcanzado valores de entre 135 y 742 mg.L-1 [12,13,30], lo que coincide con los resultados encontrados en este trabajo. La baja producción de AC en los medios 1 y 2 se puede explicar por la carencia de suficiente fuente de nitrógeno y la ausencia de aminoácidos esenciales para su síntesis como leucina, serina, valina, arginina, ornitina, entre otros, que han sido reportados por diferentes autores y sus funciones están definidas en la ruta metabólica de biosíntesis de AC [13, 31]. Actualmente para la formulación de medios de cultivo para la producción de AC se emplean mezclas de aminoácidos con el fin de proveer los nutrientes necesarios para la síntesis. Es por esto que se evaluaron los medios 4 y 5. El medio 4 contiene una mezcla compleja de aminoácidos presentes en las proteínas de la harina de soja, mientras que el medio 5, además de tener los aminoácidos presentes en harina de soja se suplementa con aminoácidos libres presentes en los casaminoácidos. Como se muestra en la Figura 1, parece ser que la presencia de aminoácidos libres incrementa la producción de AC de 100 a 330 mgL-1. Un fenómeno similar fue observado en cultivos con harina de soja y con proteína hidrolizada de soja, donde con este último componente se incrementó la concentración en un 100% [12]. La presencia de fuentes complejas de aminoácidos al parecer desfavorece la formación de biomasa incrementando el rendimiento de producto.
3.2. Evaluación de la concentración de glicerol.
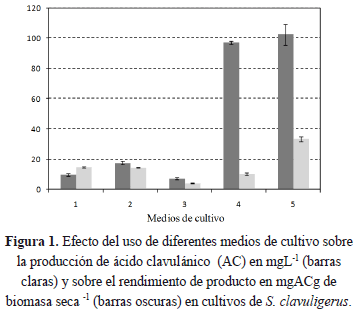
De acuerdo con los resultados de los medios de cultivo, la evaluación de la concentración de glicerol se realizó con el medio 5 y los resultados se presentan en la Figura 2. Al realizar un análisis estadístico empleando el test de rangos múltiples de Duncan [25, 26] para comparar las velocidades específicas de crecimiento y concentración de producto, se observa que hay diferencia significativa entre todos los medios para la velocidad específica de crecimiento; no hay diferencia significativa para la concentración del producto entre los medios con 2 y 10% de glicerol. Se observó un máximo en la producción de AC con 5% de glicerol alcanzando 994 mg.L-1, mientras que a 2% y 10% de glicerol la producción fue menor. El rendimiento de producto no presenta este máximo y se observa que a mayor concentración de glicerol disminuye el rendimiento, debido a que a mayor concentración de glicerol, se incrementa la formación de biomasa.
Un comportamiento tipo campana observado en la producción de AC en función de la concentración de glicerol, también ha sido reportado por otros autores, quienes indican que altas concentraciones de glicerol inhiben la producción de ácido clavulánico y reprime la producción de otros antibióticos en Sc[6, 32,33].
3.3. Fermentación en biorreactor de laboratorio
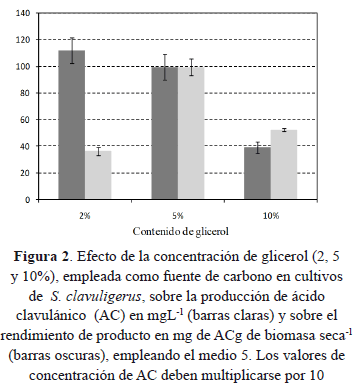
Una vez evaluado el medio apropiado para la producción de ácido clavulánico y definida la concentración de glicerol para este medio, se cultivó S. clavuligerus en un fermentador con el fin de determinar la cinética de crecimiento, consumo de sustrato y producción de ácido clavulánico y evaluar el modelo matemático propuesto. Los resultados se presentan en la Figura 3. Se observa que el AC se detectó a partir de las 24 h de cultivo pero fue a partir de las 48h que comenzó su fase de producción exponencial. Durante este periodo y hasta las 81 h se observó crecimiento de la biomasa lo que demuestra que la producción de AC está, en parte, asociada a la producción de biomasa. Debido a que la concentración de AC es muy baja hasta las 48 h, se asumió en el modelo que hasta este tiempo la producción era nula, sin embargo los resultados experimentales se muestran sin eliminar estos puntos. La velocidad de producción de biomasa cae a cero cuando la concentración de glicerol está por debajo de 4,6 gL-1, sin embargo el glicerol se continúa consumiendo hasta no detectarse a partir de las 107 h. Esto podría significar que velocidad de producción de biomasa es en realidad el balance entre formación y disrupción de material celular. A partir de las 81 h la velocidad de producción de AC experimentó una fuerte reducción correlacionándose con la presencia de una baja concentración de glicerol. Esta menor velocidad de producción se mantuvo constante hasta las 126 h. Durante este periodo, la producción de AC, no estuvo asociada a la velocidad de crecimiento sino a la concentración de biomasa presente.
De acuerdo con el comportamiento del modelo, se puede observar que hay un ajuste significativo con los datos de consumo de glicerol y de formación de AC y un mejor ajuste con los datos de producción de biomasa, cuyos valores predichos son menores que los observados durante la fase de crecimiento. Un mejor ajuste de los valores de biomasa se observa en la fase estacionaria. Los valores de los parámetros del modelo se presentan en la Tabla 1.

El valor de velocidad específica de crecimiento celular es del mismo orden que algunos de los valores reportados en la bibliografía [31, 33], aunque se aleja del máximo valor de 0,2 h-1 [29]. Los valores de rendimiento de producto Ypx obtenidos con el ajuste del modelo fueron similares a los que se presentan en las Figuras 1 y 2 para el medio 5.
El modelo propuesto ajusta bien los resultados experimentales y permite explicar el comportamiento de la fermentación con base en la teoría de las cinéticas de biomasa, sustrato y producto. Esto es, la biomasa y el sustrato limitan la velocidad específica de crecimiento, el AC es producido con componentes asociados y no asociados a la velocidad de crecimiento.
A pesar de que los modelos matemáticos son una herramienta importante para desarrollo de técnicas de optimización y control de procesos [34], existen pocos estudios al respecto para Streptomyces. Ozergin-Ulgen en 1993, propuso el primer trabajo orientado en obtener los paramétros cinéticos que describen la actividad de S. coelicolor para producir actinorrodina en cultivo en lote. El modelo de formación de producto del tipo Luedeking-Piret, permitió plantear que la producción era asociada al crecimiento [35]. Continuando con esta hipótesis, Baptista y colaboradores en el 2000, utilizaron un modelo cinético basado en la ecuación de Monod para describir los datos experimentales de Sc y emplearon un modelo cinético para el producto de efecto mixto, que considera dos términos asociado y no asociado al crecimiento [29,36]. El modelo propuesto en este trabajo incluye las consideraciones de estos autores para la formación de producto observándose un buen ajuste con los resultados experimentales.
CONCLUSIONES
Un medio de cultivo complejo con fuentes de aminoácidos libres o hidrolizados de proteínas favorece la síntesis de ácido clavulánico. Un medio como este, es el medio 5, evaluado en el presente trabajo. El glicerol empleado como fuente de carbono, mostró un valor óptimo de concentración en 50 gL-1, dentro de los valores de concentración evaluados, confirmándose que a concentraciones elevadas, inhibe la síntesis de AC. Con base en los resultados obtenidos de medio seleccionado y concentración de glicerol, se ajustaron los resultados de una fermentación en biorreactor de laboratorio y el modelo propuesto simuló con alto grado de ajuste los resultados experimentales, confirmando la bondad del modelo propuesto que incluye una cinética tipo Contois para la biomasa y una cinética de formación de producto con componentes de formación asociado y no asociado al crecimiento.
AGRADECIMIENTOS
Al Departamento Nacional de Ciencia Tecnología e Innovación, por el financiamiento del proyecto No. COL08-1-06.
A los profesores Juan Francisco Martín y Paloma Liras por el apoyo para el desarrollo de la parte experimental en el Instituto de Biotecnología INBIOTEC-León, España.
REFERENCIAS
[1] So, A.D., Gupta, N., Brahmachars, K., Chopra, I., Munos, B., Nathan, C., et al., Towards new business models for Rand D for novel antibiotics, Drug Resistance Updates. 14 (2), pp. 88-94, 2011. [ Links ]
[2] Van Den Berg, M., Gidijala, L., Kiela, J., Bovenberg, R. and Vander Keli, I., Biosynthesis of active pharmaceuticals: ß-lactam biosynthesis in filamentous fungi, Biotechnol Genet Eng Rev, 27, pp. 1-32, 2010. [ Links ]
[3] Chroma, M. and Kolar, M., Genetic methods for detection of antibiotic resistance: focus on extended-spectrum ß-lactamases, Biomedical Paper of the Medical Faculty of the University Palacký, Olomouc, Czechoslovakia. 154(4), pp. 289-96, 2010. [ Links ]
[4] Song, J.Y., Jensen, S.E. and Lee, K.J., Clavulanic acid biosynthesis and genetic manipulation for its overproduction, Appl Microbiol Biotechnol, 88(3), pp. 659-69, 2010. [ Links ]
[5] Jensen, S.E., Wong, A., Griffin, A. and Barton, B., Streptomyces clavuligerus has a second copy of the proclavaminate amidinohydrolase gene, Antimicrob Agents Chemother, 48(2), pp. 514-20, 2004. [ Links ]
[6] Teodoro, J.C., Baptista-Neto, A., Araujo, M., Hokka, C. and Badino, A., Influence of glycerol and ornithine feeding on clavulanic acid production by Streptomyces clavuligerus, Brazilian Journal of Chemical Engineering, 27(4), pp. 499 - 506, 2010. [ Links ]
[7] Viana, D.A., Carneiro-Cunha, M.N., Araújo, J.M., Barros-Neto, B., Lima-Filho, J.L., Converti, A., Pessoa-Júnior, A. and Porto, A.L., Screening of variables influencing the clavulanic acid production by Streptomyces DAUFPE 3060 strain., Appl Biochem Biotechnol, 160(6), pp. 1797-807, 2010. [ Links ]
[8] Jnawali, H.N., Yoo, J.C. and Sohng, J.K., Improvement of clavulanic acid production in Streptomyces clavuligerus by genetic manipulation of structural biosynthesis genes, Biotechnol Lett, 33(6), pp. 1221-6, 2011. [ Links ]
[9] Viana, D.A., Carneiro, M.N., Araújo, J.M., Lima-Filho, J.L., Converti, A., Pessoa-Jr A. and Figueiredo, A.L., Optimization of clavulanic acid production by Streptomyces daufpe 3060 by response surface methodology, Brazilian Journal of Microbiology, 42, pp. 658-667, 2011. [ Links ]
[10] British patent Specification No.1,571,888. Clavulanic Acid Production. Greenford, U.K., 1980. [ Links ]
[11] Patente Española No. 537,158. Un Procedimiento para Mejorar la Extracción de ácido clavulánico. Madrid, España, 1984. [ Links ]
[12] Ortiz, S.C., Hokka, C.O. and Badino A.C., Utilization of soybean derivatives on clavulanic acid production by Streptomyces clavuligerus, Enzyme and Microbial Technology, 40, pp. 1071-1077, 2007. [ Links ]
[13] Li, R. and Townsend, C.A., Rational strain improvement for enhanced clavulanic acid production by genetic engineering of the glycolytic pathway in Streptomyces clavuligerus., Metab. Eng., 8(3), pp. 240-52, 2006. [ Links ]
[14] Teodoro, J.C., Baptista-Neto, A., Cruz-Hernández, I.L., Hokka, C.O. and Badino, A.C., Influence of feeding conditions on clavulanic acid production in fed-batch cultivation with medium containing glycerol., Appl Microbiol Biotechnol, 72(3), 450-5, 2006. [ Links ]
[15] Baños, S, Perez-Redondo, R., Koerman, B. and Liras, P., Glycerol Utilization Gene Cluster in Streptomyces clavuligerus, Appl Environ Microbiol, 75(9), pp. 2991-2995, 2009. [ Links ]
[16] Saudagar, P.S. and Singhal, R.S., Optimization of nutritional requirements and feeding strategies for clavulanic acid production by Streptomyces clavuligerus., Bioresour Technol, 98(10), pp. 2010-7, 2007. [ Links ]
[17] Silva C. D., Cuel M.F., Barreto V.O., Kwong W.H., Hokka C. O. and Barboza M. 29(3), pp 428-31, 2012 [ Links ]
[18] Kirk, S., Avignone-Rossa, C. A. and Bushell, M. E., Growth limiting substrate affects antibiotic production and associated metabolic fluxes in Streptomyces clavuligerus, Biotechnology Letters., 22, pp. 1803-1809, 2000. [ Links ]
[19] Saudagar, P. S., Survase, S. A., and Singhal, R. S., Clavulanic acid: A review, Biotechnology Advances, 26(4), pp. 335-351, 2008. [ Links ]
[20] Pérez, R., SantaMarta, I., Bovenberg, R. and Martín, J.F., The enigmatic lack of glucose utilization in Streptomyces clavuligerus is due to inefficient expression of the glucose permease gene, Microbiology, 156, pp. 1527-1537, 2010. [ Links ]
[21] Liras, P., Gómez-Escribano, J. and SantaMarta, I., Regulatory mechanisms controlling antibiotic production in Streptomyces clavuligerus, 35, pp. 667-676, 2008. [ Links ]
[22] Liras, P. and Martín, J.F., Assay Methods for Detection and Quantification of Antimicrobial Metabolites Produced by Streptomyces clavuligerus, en: Methods in Biotechnol. 18: Microbial Processes and Product, New Jersey, pp. 149-163, 2005 [ Links ]
[23] Malmberg, L.H., Hu, W.S. and Sherman, D.H., Precursor flux control through targeted chromosomal insertion of the lysine epsilon-aminotransferase (lat) gene in cephamycin C biosynthesis, J Bacteriol, 175(21), pp. 6916-24, 1993. [ Links ]
[24] Botas, Alma. Comunicación personal con investigadora del Laboratorio de Microbiología, Universidad de León y ensayado en el Instituto INBIOTEC (España). [ Links ]
[25] Duncan, D., Multiple range and multiple F-test, Biometrics, 11(1), pp. 1-42, 1955. [ Links ]
[26] Kramer, C., Extension of multiple range test to group mean with unequal numbers of replications. Biometrics, 12(3), pp. 307-310, 1956. [ Links ]
[27] Burton, K., A study of the conditions and mechanism of the diphenylamine reaction for the colorimetric estimation of deoxyribonucleic acid., Biochem J, . 62(2), pp. 315-23, 1956. [ Links ]
[28] Foulstone, M. and Reading, C., Assay of amoxicillin and clavulanic acid, the components of Augmentin, in biological fluids with high-performance liquid chromatography, Antimicrob Agents Chemother, 22(5), pp. 753-62, 1982. [ Links ]
[29] Neto, A., Gouveia, E., Badino-Jr, A., and Hokka, C., Phenomenological model of the clavulanic acid production process utilizing Streptomyces clavuligerus, Brazilian Journal of Chemical Engineering, 17(4-7), pp. 1-12, 2000. [ Links ]
[30] Neto, A.B., Hirata, D.B., Cassiano Filho, L.C., Bellão, C., Badino Júnior, A.C., and Hokka, C.O., A study on clavulanic acid production by Streptomyces clavuligerus in batch, fed batch and continuous processes, Brazilian Journal of Chemical Engineering, 22(4), pp. 557 - 563, 2005. [ Links ]
[31] Ives, P.R. and Bushell, M.E., Manipulation of the physiology of clavulanic acid production in Streptomyces clavuligerus. Microbiology, 143, 3573-9, 1997. [ Links ]
[32] Chen, K.C., Lin, Y.H., Tsai, C.M., Hsieh, C.H. and Houng, J.Y., Optimization of glycerol feeding for clavulanic acid production by Streptomyces clavuligerus with glycerol feeding, Biotechnology Letters, 24(6), pp. 455-458, 2002. [ Links ]
[33] Efthimiou, G., Thumser, A.E. and Avignone-Rossa, C.A., A novel finding that Streptomyces clavuligerus can produce the antibiotic clavulanic acid using olive oil as a sole carbon source, Journal of Applied Microbiology, 105(6), pp. 1365-2672, 2008. [ Links ]
[34] Trejos, V.M., Fontalvo, A. J. and Gómez, G. M. A., Descripción matemática y análisis de estabilidad de procesos fermentativos, Dyna, 76(158), pp. 111-121, 2009. [ Links ]
[35] Ozergin-Ulgen, K. and Mavituna, F., Actinorhodin production by Streptomyces coelicolor A3(2): Kinetic parameters related to growth, substrate uptake and production, App. Microbiol. Biotechno., 40, pp. 457-462, 1993. [ Links ]
[36] Neto, Á., Teodoro, J., Macedo, F. C., Battaglia, H. D., Badino-Jr, A. and Hokka, C., Kinetic studies on clavulanic acid production by Streptomyces clavuligerus, Memorias II Mercosur Congress on Chemical Eng. And IV Mercosur Congress on Process System Eng. Village Rio das Pedras, Club Med, Rio de Janeiro, pp. 1-10, Agosto de 2005. [ Links ]
