











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares em
SciELO
Similares em
SciELO  Similares em Google
Similares em Google 
 Permalink
Permalink1. Introducción
Los avances producidos en el estudio del sistema nervioso en los últimos años han proporcionado una ingente cantidad de nuevos datos relativos a los mecanismos de codificación, transformación, coordinación, creación, ejecución y aprendizaje de información en distintos tipos de redes neuronales biológicas [1]. Uno de estos nuevos descubrimientos es un tipo de código temporal particular y discriminatorio denominado firma neuronal [2,3]. Las firmas neuronales así definidas son estructuras temporales robustas y específicas que se localizan en los intervalos entre potenciales de acción de las señales generadas por una neurona con comportamiento en ráfagas. Aunque todavía no se ha demostrado experimentalmente si tienen algún significado funcional, trabajos computacionales sugieren que estas estructuras temporales precisas pueden jugar un papel crítico en la codificación de la información neuronal ya que permiten identificar la fuente de la señal. El tratamiento de la información basada en la identificación del emisor del mensaje (señal neuronal) podría ser una potente estrategia de procesamiento y codificación para los sistemas neuronales, que incrementaría en gran medida sus propiedades computacionales. Simulaciones de modelos de circuitos generadores de patrones indican que estas firmas neuronales pueden desempeñar un rol funcional clave en los circuitos donde están presentes [4,5,6]. Por otra parte, las firmas neuronales que identifican cada célula podrían ser parte de una estrategia multicódigo según la cual una neurona podría multiplexar en una misma señal de salida distinto tipo de información codificada simultáneamente con distintos códigos [5,6]. Sin embargo, desde un punto de vista teórico, se han desarrollado muy pocos trabajos sobre mecanismos de procesamiento de información basado en la emisión y el reconocimiento de firmas neuronales específicas o, dicho de otro modo, basados en la identificación del origen de la señal. En este escenario, se requiere la realización de estudios teóricos y computacionales detallados.
En esta línea, en los últimos años, además de los modelos generadores centrales de patrones, se han construido distintos modelos bio-inspirados a gran escala para estudiar la habilidad de las redes neuronales para procesar la información basada en la emisión y reconocimiento de firmas neuronales [7,8]. En estos trabajos, por ejemplo, se han analizado las dinámicas colectivas de diferentes modelos de red en respuesta a estímulos introducidos en unas pocas neuronas dentro de la red o se discuten los efectos de los patrones de conectividad en las propiedades auto-organizativas de la red.
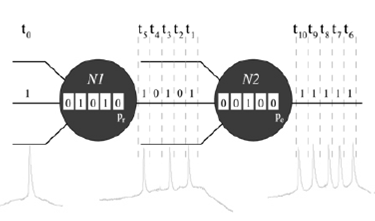
Fuente: Carrillo-Medina JL and Latorre R., 2015
Figura 1 Representación esquemática del proceso de reconocimiento de firmas en el modelo de neurona de acuerdo a [8], Para simplificar, en este ejemplo consideramos un canal de entrada y su correspondiente contexto de información local (secuencia de 5 bits representados al interior de cada neurona). En el ejemplo, las neuronas reconocen una única firma neuronal de 5 bits, F = (1, 0, 1, 0, 1), y el patrón binario espontáneo (1, 1, 1, 1, 1). En el paso de tiempo t0, la entrada de la neurona N1 es 1. El contexto de información local de esta unidad indica que, en las 5 iteraciones previas, este ha recibido la secuencia (0, 1, 0, 1, 0). Entonces, cuando en t0 la entrada es procesada, N1 reconoce F - es decir, el nuevo contexto de información local contiene el patrón (1, 0, 1, 0, 1). Por lo tanto, en los siguientes 5 pasos de tiempo (t1 − t5), esta neurona emite el patrón binario serial (1, 0, 1, 0, 1) con la probabilidad pr. Entonces, asumiendo que N1 emite F, la entrada de N2 en el paso de tiempo t1 es 1. En este caso, si asumimos que el contexto de información local de N2 en este paso de tiempo es la secuencia (0, 0, 1, 0, 0), la neurona no reconoce ninguna firma y emite la actividad espontánea con probabilidad pe.
Los resultados previos señalan que modelos de red muy simples son capaces de mostrar una dinámica colectiva compleja basada en la emisión y reconocimiento de firmas neuronales específicas. En este artículo, discutimos el efecto de variar los parámetros que afectan las propiedades auto-organizativas de estas redes, de forma que existan poblaciones de neuronas dentro de la red que se especialicen en el reconocimiento de emisión de determinadas firmas. Para lo cual hemos variado la probabilidad de reconocimiento que tiene cada neurona para distintas firmas en la red.
2. Material y Métodos
2.1. Modelos de red y neuronas
Para modelar la dinámica individual de la red hemos utilizado el modelo de red binaria con la capacidad de reconocer y emitir firmas propuesto en [7,8], ver Fig. 1.
El modelo considera que una firma es una secuencia de unos y ceros que puede ser reconocida por cada una de las neuronas individuales de la red con una cierta probabilidad. Cada neurona tiene 8 canales sinápticos de entrada en función de las conexiones que se establecen con otras unidades, según el patrón de conectividad utilizado en la simulación (como se indica más abajo). Cada neurona emite una firma a sus vecinas con probabilidad pe. Esta firma define la actividad espontánea de la red. En ausencia de un estímulo la red evoluciona a un estado estacionario, relacionada con la emisión de la actividad espontánea. Cada unidad puede responder al reconocimiento de un conjunto de firmas específicas recibidas a través de sus canales de entrada. El reconocimiento de una firma en cada intervalo de tiempo se chequea usando un contexto de información local [9], que es una memoria transitoria que mantiene un registro de las entradas recibidas en los instantes de tiempo anteriores. Cuando una firma es reconocida, la neurona emite la misma firma a través de sus canales de salida con probabilidad pr (pr >>> pe). Si no se reconoce ninguna firma, se emite la firma espontánea con una pe. Además de los canales de conexión (canales sinápticos) que tiene cada neurona, está tiene un canal adicional para inyectar estímulos externos (firmas) en la red. Si una neurona reconoce una firma entonces la emite a sus vecinas con una probabilidad pr, lo que puede producir cambios en la dinámica espontánea de la red debido a la propagación del estímulo externo.
La dinámica colectiva de la red se basa en calcular su evolución temporal según las reglas de reconocimiento y emisión de firmas descritas anteriormente. De esta forma, pe controla el nivel de actividad espontánea y pr la intensidad de la actividad asociada con el estímulo externo. Mientras mayor es la probabilidad de pe y de pr mayor será la actividad de la firma espontánea y de la estimulación respectivamente. Después de emitir una firma la neurona tiene un período de refracción donde ni reconoce ni emite ninguna firma. En [12] se ha estudiado cómo afecta a la dinámica de la red la duración del periodo de refracción.
Como se indicaba anteriormente, cada neurona está conectada a otras ocho neuronas de la red. Dado que nuestro objetivo es comparar la dinámica de redes heterogéneas frente a la observada en redes homogéneas, en las simulaciones que vamos a discutir en este trabajo hemos construido redes análogas a las discutidas en [8]. En todos los casos hemos simulado redes de 2500 neuronas que pueden estar conectadas con tres posibles topologías de red: regular, small-world o aleatoria. Para construir las redes small-world partimos de una red bidimensional de tamaño 50x50 con condiciones periódicas de contorno y rompemos conexiones con cierta probabilidad p (p es el grado de regularidad de la red) para establecer una nueva conexión con otra neurona de manera aleatoria [10,11]. De esta forma, las redes regulares (p=0) y aleatorias (p=1) son las condiciones extremas para las topologías small-world. Con el fin de comparar nuestros resultados con los obtenidos en redes homogéneas [8], además de las topologías regulares y aleatorias, en este trabajo vamos a simular redes con dos posibles topologías small-world (p=0.10 y p=0.25).
3. Resultados
En trabajos previos con el modelo descrito en la Sección 2, los parámetros que afectan a la dinámica de la red siempre están fijos (ver [8] para una descripción detallada). En donde la probabilidad de reconocimiento pr se fijó al 80%, la probabilidad de emisión de la firma espontánea pe al 5%, el tamaño de la firma (Tf) es 5 bits, el período de refracción Rp se fijó en 10 pasos de tiempo y los estímulos externos los recibe una única neurona. Las simulaciones aquí presentadas tienen los mismos parámetros, con la diferencia de que la probabilidad de reconocimiento es distinta para cada una de las firmas A y B. En particular, nos vamos a centrar en el caso en donde la firma A tiene la probabilidad de reconocimiento del 85% mientras que la firma B tiene un 80%.
3.1. Actividad espontánea
La actividad espontánea es la dinámica intrínseca de la red incluso en ausencia de estímulos externos. En los trabajos previos, concluimos que el nivel de la actividad espontánea en la red depende principalmente de la probabilidad pe. Para este tipo de simulaciones si variamos la probabilidad de reconocimiento varía el número promedio de neuronas que emiten la firma espontánea por unidad de tiempo, ver [8]. Pero es equivalente en cada una de las topologías de red, ver [8]. La única diferencia es el nivel constante alcanzado por medio de la actividad espontánea con las diferentes probabilidades de reconocimiento.
3.2. Codificación con estímulos externos
Como en los estudios previos los resultados más interesantes son las simulaciones relacionadas con la codificación de estímulos externos. Para la codificación de estímulos hemos realizado dos tipos de simulaciones, una con poblaciones homogéneas al igual que en [7,8] y otra con poblaciones heterogéneas. Las simulaciones con poblaciones homogéneas se las realizó fijando los parámetros que afectan la red: pe = 0.05, pr = 0.80, Nns = 10, Rp = 10, Tf = 5 bits, Numero de firmas (Nf) = 1 o 2. En el caso de las simulaciones con poblaciones heterogéneas se fijó los parámetros pe = 0.05, Rp = 10, Tf = 5 bits y Nf = 1 o 2. En donde la firma A tiene una probabilidad pr de 0.85 y la firma B con pr igual a 0.80. En ambos casos cuando se estimula la red con estas pr obtenemos una red con memoria a largo plazo o permanente. En las simulaciones que se presentan aquí, inicialmente dejamos evolucionar a la red libremente hasta el intervalo de tiempo 5000, es decir ningún estímulo se introduce en la red. En el intervalo de tiempo 5000, una firma externa se introduce dentro de n neuronas escogidas aleatoriamente durante el intervalo 10000. Entonces, la red evoluciona libremente sin ninguna estimulación de nuevo, evolucionando a un nivel constante de neuronas que reconocen la una o la otra firma, mismas que se mantiene en el tiempo, pero con diferente nivel durante y después de la estimulación. La finalidad de este tipo de simulaciones es obtener redes equivalentes (ver sección 3.5 de [8]) después de finalizado la estimulación, es decir, cuando pr = 0.85, queremos obtener una red equivalente cuando pr = 0.80, partiendo para ambos casos con Nns = 10, Para ello calculamos Nns de acuerdo a la topología de red, hasta que ambas redes puedan ser comparadas bajo las mismas condiciones.
3.3. Codificación con estímulos múltiples
Desde la perspectiva de codificación de información, una característica interesante a ser evaluada es la habilidad de la red para procesar y codificar múltiples estímulos, simultáneamente. Tomando en cuenta los resultados descritos en la sección anterior, la competencia entre la actividad espacio-temporal generada por la red como respuesta a diferentes estímulos y la influencia de poblaciones homogéneas y/o heterogéneas en esta competición, nosotros hemos realizado simulaciones donde diferentes estímulos se introducen en la red.
3.3.1. Simulación en serie
En simulaciones con poblaciones homogéneas, alternamos episodios de simulación de dos diferentes firmas (A y B), cuando son inyectadas en el mismo grupo de células. Inicialmente, la red evoluciona libremente. En el intervalo de tiempo 5000, la estimulación inicia. En cada episodio de simulación, una firma es inyectada en la red para 10000 unidades de tiempo. Entonces, hay un intervalo de inter-estimulaciones de 5000 intervalos de tiempo donde ningún estímulo se introduce dentro de la red. Esta secuencia se repite ocho veces alternando dos diferentes firmas neuronales. Finalmente, en el intervalo de tiempo 120000 no se introduce ningún estímulo más.
La Fig. 2. muestra gráficos que permiten comparar la evolución del número de neuronas que emiten la firma espontánea (trazos azules), la firma A (trazos rojos) y la firma B (trazos verdes) en redes con memoria a largo plazo con diferentes topologías de red. Observe los diferentes intervalos durante los cuales las dos firmas externas compiten. La Fig. 2. indica además que la competencia entre dos firmas externas es una competencia con ganador (winner-take-all) donde la firma activa prevalece sobre las otras. No obstante, en redes regulares de neuronas con un período refractario pequeño, existen competencias transitorias sin ganador (winnerless) [12-14], que pueden emerger en la red, las firmas A y B pueden coexistir en la red (ver paneles A, B en la Fig. 2). Durante estos episodios transitorios, el estímulo activo no gana la competencia como en una dinámica con ganador (winner-take-all) y, por lo tanto, los patrones correspondientes que codifican este estímulo no reemplazan completamente los patrones relacionados a la firma recibida anteriormente. Este es un interesante fenómeno desde el punto de vista de codificación y procesamiento de información porque la competencia sin ganador (winnerless) permite a la red codificar dos firmas diferentes a través de una de ellas que no está sostenida por la estimulación.
En redes más regulares (paneles A y B de la Fig. 2), los patrones espacio-temporales en evolución tienen una estructura espacial bien definida y coherente. La clusterización y la coherencia de estos patrones transitorios producen regiones de frontera bien delimitados entre los estímulos A y B, que potencia la competencia. Esto mejora la capacidad de la red para sostener múltiples estímulos simultáneamente. De la misma manera, la competencia con ganador observada en las redes más aleatorias en la Fig. 2 (paneles C y D de la Fig. 2) se observa una propagación no estructurada de estímulos que rápidamente reemplazan los estímulos inducidos por el estímulo anterior dentro de la red.
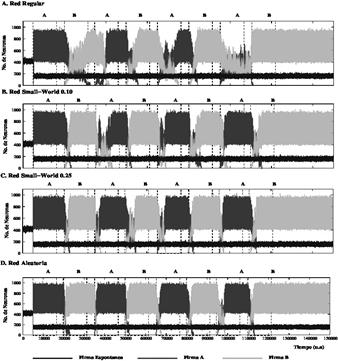
Fuente: Carrillo-Medina JL and Latorre R., 2015
Figura 2 Comparación del número promedio de neuronas que reconocen y emiten las firmas A y B en cuatro redes de memoria de largo plazo con diferentes topologías para poblaciones de neuronas homogéneas. Dada la topología de red, durante los períodos de estimulación, siempre surge una competencia entre la actividad inducida por firmas A y B. En redes con un alto grado de conexiones regulares (paneles A y B) la competencia entre firmas A y B puede ser una competencia con o sin ganador (winner-take-all, winnerless). En la mayoría de redes aleatorias (panel C y D), existe siempre una competencia con ganador (winner-take-all) en la cual la actividad sostenida por el estímulo activo siempre prevalece.
En simulaciones con poblaciones heterogéneas, alternamos episodios de simulación de dos diferentes firmas (A y B), cuando son inyectadas en diferentes grupos de células y con diferentes pr, de acuerdo a los parámetros indicados en la sección 3.2.
Al igual que en las redes con poblaciones homogéneas, inicialmente, la red evoluciona libremente. En el intervalo de tiempo 5000, la estimulación inicia. En cada episodio de simulación, una firma es inyectada en la red para 10000 unidades de tiempo. Entonces, hay un intervalo de inter-estimulaciones de 5000 intervalos de tiempo donde ningún estímulo se introduce dentro de la red. Esta secuencia se repite ocho veces alternando dos diferentes firmas neuronales. Finalmente, en el intervalo de tiempo 120000 no se introduce ningún estímulo más. Estas simulaciones se las realizó por separado, una simulación para la firma A y otra simulación para la firma B. Para poder comparar los dos tipos de simulaciones se hizo necesario obtener redes equivalentes (con poblaciones heterogéneas) en las diferentes topologías de red, determinando que es inevitable aumentar el Nns, en este caso específico cuando la pr de una de firma A es mayor que la firma B. Obteniéndose los valores que se indican en la Tabla 1. El número promedio de neuronas que reconocen y emiten una firma (Nnref), durante la estimulación se incrementa debido al Nns. Después de la estimulación el Nnref decrece. Esto se debe a que el número de neuronas seleccionadas en las cuales se inyecta el estímulo, deja de recibirlo. La competencia entre firmas se la realiza una vez terminado el episodio de estimulación de la red con una firma dada. De las simulaciones podemos indicar que cuando más regular es la red mayor es el número de neuronas seleccionadas para obtener una red equivalente.
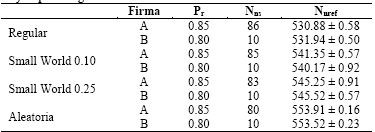
Tabla 1 Número promedio de neuronas que reconocen y emiten una firma una vez finalizado la inyección del estímulo, cuando el primer estímulo tiene una pr mayor que el segundo.
Fuente: Elaboración propia

Fuente: Elaboración propia
Figura 3 Evolución de la actividad de una de red SW10 con memoria a largo plazo que recibe dos estímulos en serie. La red consta de 2500 neuronas con los parámetros intra-unidad mostrados en la Tabla 1. El estímulo A es (1,0,0,0,1) con una pr de 0.85 y el estímulo B (1, 1, 0, 1, 1) con una pr de 0.80. Las etiquetas en la parte superior indican el estímulo inyectado en cada episodio de la estimulación. La figura representa gráficamente la evolución de la actividad espontánea intrínseca (trazo azul) y el nivel de emisión de las firmas asociadas a los estímulos A (Trazo rojo) y B (trazo verde). En redes con memoria a largo plazo, existen pequeñas diferencias resultantes de la topología de red en las dinámicas colectivas de la red. En estos casos el estímulo previo de un episodio siempre sobrevive, cuando se inicia un nuevo episodio de la estimulación, el estímulo previo gana instantáneamente la competencia (con ganador) y el estímulo nuevo desaparece completamente de la red.
La Fig. 3. muestra un gráfico representativo de las cuatro redes el cual permite comparar la evolución del número de neuronas que emiten la firma A (trazos rojos) y la firma B (trazos verdes) en redes con memoria a largo plazo con diferentes topologías de red, pr (Firma A, pr = 0.85; Firma B, pr = 0.80) y Nns (Firma A, Nns = 86; Firma B, Nns = 10). En este tipo de simulaciones hemos alternado episodios de simulación de dos diferentes firmas (A y B) con los valores obtenidos en la Tabla 1. Nosotros podemos observar en los diferentes intervalos durante los cuales las dos firmas externas se estimulan que siempre existe una competencia con ganador (winner-take-all) donde la firma A prevalece sobre la firma B, debido a su mayor pr, es decir siempre gana la firma que tiene mayor probabilidad, aunque se aumente el Nns de la firma que tiene menor probabilidad, no obstante, estén en las mismas condiciones para competir, ver Tabla 1. Esto sucede independientemente de la topología de red. Cabe notar que en estas simulaciones no existe una competencia global en la red, existe una competencia local, misma que esta sostenida solo cuando se estimula la red con la firma B.
En base a las simulaciones anteriores hemos aumentado el Nns para la firma B (cuando pr de la firma B es menor que la firma A), lo cual permite iniciar una competencia local en una competencia global dentro de la red. En función del Nns de la firma A (cuando Nns = 10) encontrándose valores de Nns para la Firma B, como se indica en la Tabla 2. De acuerdo a esta tabla podemos indicar que mientras más conexiones regulares tengan una red, mayor debe ser el número de neuronas seleccionadas en las cuales se inyecte el estímulo.
La Fig. 4. muestra el comportamiento de las cuatro redes de acuerdo a los valores de la Tabla 2, lo que permite obtener competencias globales durante los episodios transitorios. El estímulo activo de la firma B no gana la competencia como en una dinámica con ganador (winner-take-all) y, por lo tanto, los patrones correspondientes que codifican este estímulo no reemplazan completamente los patrones relacionados a la firma recibida anteriormente. Este es un interesante fenómeno desde el punto de vista de codificación y procesamiento de información porque la competencia sin ganador (winnerless) permite a la red codificar dos firmas diferentes a través de una de ellas que no está sostenida por la estimulación. En este tipo de simulaciones siempre existe una competencia con ganador y es el estímulo que tiene la mayor pr, independiente de la topología de red. Donde se representa la firma A (trazos rojos), la firma B (trazos verdes) y la actividad espontánea (trazos azules).
Tabla 2 Número de neuronas seleccionadas en las cuales se inyectan las firmas A y B para que empiece existir competición global.

Fuente: Elaboración propia
Fuente: Elaboración propia
Figura 4 Comparación del número promedio de neuronas que reconocen y emiten las firmas A y B en cuatro redes de memoria de largo plazo con diferentes topologías para poblaciones de neuronas heterogéneas. Dada la topología de red, durante los períodos de estimulación, siempre surge una competencia entre la actividad inducida por las firmas A y B de manera local. En este tipo de redes existe siempre una competencia con ganador (winner-take-all) en la cual la actividad sostenida por el estímulo activo siempre prevalece.
Tabla 3 Número de neuronas seleccionadas para estimular la red y número promedio de neuronas que reconocen y emiten una firma, para obtener cuatro redes equivalentes.

Fuente: Elaboración propia
Con las simulaciones realizadas no se pudo obtener redes equivalentes para ello equiparamos el número de neuronas que reconocen y/o emiten una de las firmas para que en estas condiciones observar la competencia entre las dos firmas y determinar si existe competencia con ganador o sin ganador. Para lo cual hemos variado el Nns en cada una de las topologías de red, ver Tabla 3.
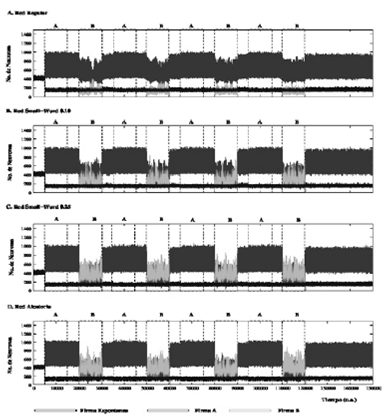
La Fig. 5 muestra un gráfico representativo de las cuatro redes el cual permite comparar la evolución del número de neuronas que emiten la firma A (trazos rojos) y la firma B (trazos verdes) en redes con memoria a largo plazo con diferentes topologías de red, pr (Firma A, pr = 0.85; Firma B, pr = 0.80) y Nns (Firma A, Nns = 10; Firma B, Nns = 550) de acuerdo a la Tabla 3.

Fuente: Elaboración propia
Figura 5 Evolución de la actividad representativa de cuatro redes equivalentes; Red Regular con memoria a largo plazo que recibe dos estímulos en serie. La red consta de 2500 neuronas con los parámetros intra-unidad mostrados en la Tabla 3. El estímulo A es (1, 0, 0, 0,1) con una pr de 0.85 y el estímulo B (1, 1, 0, 1, 1) con una pr de 0.80. Las etiquetas en la parte superior indican el estímulo inyectado en cada episodio de la estimulación. La figura representa gráficamente la evolución de la actividad espontánea intrínseca (trazo azul) y el nivel de emisión de las firmas asociadas a los estímulos A (trazo rojo) y B (trazo verde). En redes con memoria a largo plazo, con diferentes pr y Nns pero en las mismas condiciones, su comportamiento es el mismo que con redes aleatorias con poblaciones de neuronas homogéneas. En estas redes no existe competencia ya que casi instantáneamente el estímulo activo prevalece ante el estímulo previo.
En este tipo de simulaciones hemos alternado episodios de simulación de dos diferentes firmas (A y B). Nosotros podemos observar, en los diferentes intervalos durante los cuales las dos firmas externas se estimulan, que siempre existe una competencia con ganador (winner-take-all) donde la firma activa prevalece sobre la firma previa en forma instantánea, independientemente de que exista en los episodios transitorios una disminución del número de neuronas que reconocen o emiten la firma que tiene menor pr. En estas redes siempre gana la firma que se está inyectando, existiendo una competencia global en la red. Esto sucede independientemente de la topología de red.
En todas las topologías de red se puede observar que, en los episodios transitorios de una firma a otra, existe una disminución del número promedio de neuronas que reconocen o emiten una firma dada, esto se produce debido a que se deja de estimular grandes grupos de neuronas.
En todas las redes, independientemente de la topología de red, en los intervalos de estimulación podemos indicar que siempre existe una competencia con ganador de forma inmediata y esta es la firma con la cual se estimula la red.
4. Discusión
Los resultados experimentales hacen hincapié en que el sistema nervioso utiliza diferentes estrategias para la codificación de la información [15,16], lo que mejora en gran medida su capacidad de procesamiento [17]. Una hipótesis interesante desde el punto de vista del procesamiento de información es que el sistema nervioso puede utilizar diferentes códigos al mismo tiempo, lo que resulta en una multiplexación de información [5,6,18,19]. En este contexto, el procesamiento de la información basada en el reconocimiento de firmas neuronales específicas emerge como una estrategia de codificación de gran alcance para tenerse en cuenta en los circuitos neuronales [20]. Este tipo de codificación permitiría que los receptores de señales discriminen la información neuronal en función de la fuente de señal, procesándose o rechazándose directamente las entradas de ciertos emisores. De confirmarse la existencia de mecanismos celulares que permitan la utilización de este tipo de estrategia, implicaría que las firmas neuronales son un mecanismo general para procesar y codificar información en el sistema nervioso.
Descubrimientos recientes demuestran que modelos simples de redes neuronales pueden implementar una estrategia de codificación y procesamiento basado en firmas [8]. Estos modelos muestran propiedades auto-organizativas complejas que una red de neuronas que emite y procesan firmas puede desplegar.
En este artículo, hemos analizado el efecto que una característica adicional puede tener en la red propuesta, es decir, neuronas con diferentes probabilidades de reconocimiento de firmas, las cuales generan diferentes dinámicas colectivas e incrementan las propiedades dinámicas de la red. Además, muestran que la red presenta capacidades de codificación adicional y mecanismos para adaptar la estrategia de procesamiento a situaciones diferentes. Los resultados reportados sugieren que la mayoría de modelos requieren determinar los beneficios que el procesamiento de información, basado en el reconocimiento de firmas neuronales específicas, puede proporcionar a una red neuronal, por ejemplo, para ponderar la salida de una neurona, basada en el reconocimiento de firmas específicas y/o en el contenido del resto de la señal (mecanismo de contextualización), que en un momento dado emiten firmas funcionales diferentes en la salida y estas pueden realizar tareas específicas por cada neurona.

