











 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares em
SciELO
Similares em
SciELO  Similares em Google
Similares em Google 
 Permalink
Permalink1. Introduction
Clavulanic acid (CA) is a potent inhibitor of β-lactamases, enzymes synthesized by some gram-positive and gram-negative bacteria, which provide them a defense mechanism against β-lactam antibiotics. CA is commonly used along with β-lactam antibiotics, resulting in a more efficient antibiotic with a broader spectrum of antimicrobial activity. CA is synthesized by Streptomyces clavuligerus, and is produced at large scale by conventional submerged culture processes. Besides CA, S. clavuligerus also produces secondary metabolites such as cephamycin C and various clavam structure compounds, with antifungal and antitumoral activities [1].
Due to its clinical and industrial importance, attempts to improve CA titers and productivity have focused on assessing the effect of nutrient concentration, environmental conditions and the species metabolic capabilities for CA biosynthesis. The composition of the culture medium is a very important issue; S. clavuligerus is able to metabolize diverse carbon sources, including oils and carbohydrates, but the primary carbon substrate for CA biosynthesis is glycerol. Culture media composed by cereal flours and digested proteins supply the amino acids required in the biosynthesis of clavam-related metabolites [2].
Selection strategies for different S. clavuligerus strains have been designed by the analysis of their metabolic capabilities and their ability to produce CA at high levels, under nutrient stress conditions and/or media supplementation. In some cases, interest has been focused into identifying CA overproducer strains able to metabolize low-cost substrates. However, issues such as genetic variability and overproduction of unwanted metabolites (such as clavams) have suggested the rational manipulation of metabolic pathways as a promising alternative for overproducer strains [3].
In this work, we performed a comparative study of the metabolic capabilities of two standard reference strains of S. clavuligerus to metabolize diverse media components and produce CA; the strains used were S. clavuligerus DSM 41826 (ScDSM) and S. clavuligerus ATCC 27064 (ScATCC). Various synthetic, semi-synthetic and complex culture media were tested for their suitability to induce and/or improve CA biosynthesis. Furthermore, the effect of the nitrogen and phosphorous source on CA biosynthesis was evaluated for the selected strain (ScATCC). The selection procedure was supported by an economic analysis.
2. Materials and methods
2.1. Microorganism and culture conditions
S. clavuligerus ATCC 27064 (ScATCC) and S. clavuligerus DSM 41826 (ScDSM) were used throughout this study. Mycelium was obtained following the standard protocol for Streptomyces [4], and was stored in 1.5 mL Eppendorf tubes with a 20 % glycerol sterile solution at -80 °C. The seed medium consisted of 50 mL TSB medium (30 g·L-1; pH 7.0) in 250 mL baffled Erlenmeyer flasks inoculated with mycelium stock and incubated at 28 ºC for 36h at 220 rpm. The following culture media were used:
Synthetic media (SM): A basal synthetic medium was composed of [g/L]: glycerol, 15; sucrose, 20; NaCl, 5.0; L-proline, 2.5; K2HPO4, 2.0; MgSO4·7H2O, 1.0; CaCl2, 0.4; MnCl2·4H2O, 0.1; FeCl3·6H2O, 0.1; ZnCl2, 0.05; MOPS, 20.9. The N-source was either 1.5 g/L glutamic acid (medium GSPG) or 1.5 g/L L-asparagine (medium GSPA) [5].
Semi-synthetic media (SSM): Two semi-synthetic media were used: SA (Starch-Asparagine) was composed of (g/L): soluble starch, 10.0; asparagine, 2.0; MgSO4, 0.60; K2HPO4, 4.40; MOPS, 20.0; MgSO4·7H2O, 0.75; MnCl2·4H2O, 0.001; FeSO4·7H2O, 0.001 and ZnSO4·7H2O, 0.001, pH was adjusted at 6.8 using NaOH [6]. NZ-Amine medium was composed of (g/L): NZ-Amine (casein hydrolyzate) 15.0; glycerol 15; NaCl 5.0; K2HPO4, 2.0; MgSO4·7H2O; MOPS, 21.0, and 1 ml/L of a trace elements solution containing (g/L): MnCl2·4H2O, 0.1; FeSO4·7H2O, 0.1; ZnCl2, 0.05; CaCl·2H2O, 0.4.
Complex media (CM): Four different soy-based media were used: Tryptic Soy Broth (TSB) composed of (g/L): TSB, 30.0; glycerol, 15.0; MOPS, 21.0. Soy protein isolate (ISP) composed of (g/L): glycerol, 15.0; soy protein isolate, 10.0; malt extract, 10.0; yeast extract, 1.0; K2HPO4, 2.5; MgSO4·7H2O, 0.75; MnCl2·4H2O, 0.001; FeSO4·7H2O, 0.001; ZnSO4·7H2O, 0.001; MOPS, 21.0 [7].
Soybean flour based medium (SF): composed of: glycerol, 15.0; soybean flour, 10.0; malt extract, 10.0; yeast extract, 1.0; K2HPO4, 2.5; MgSO4·7H2O, 0.75; MnCl2·4H2O, 0.001; FeSO4·7H2O, 0.001; ZnSO4·7H2O, 0.001; MOPS, 21.0.11 Soybean flour with casamino acids (SF-CSS), same composition as SF with addition of 5 g/L casamino acids [8].
All cultures were performed in 250-baffled Erlenmeyer flasks containing 50 mL of medium. Flasks were inoculated with seed medium (10% v/v) as described above, at an average initial biomass concentration of 0.09 ± 0.015 g·L-1. Cultures were incubated for 120 h, at 220 rpm and 28 ºC.
2.2. Analytical techniques
Clavulanic acid was determined by HPLC using an Agilent 1200 Series equipped with a Diode Array Detector, using a reverse phase column ZORBAX Eclipse XDB-C18 (4.6 x 150 mm, 18 μm). The mobile phase was a 96% v/v KH2PO4 (50 mM, pH 3.2) and 6% v/v methanol solution at a flow rate of 1mL/min. Culture samples were centrifuged at 14000 g for 10 min at 4°C and filtered through a 0.22 µm membrane. The filtered sample (300 µL) was mixed with 100 µL of imidazole reagent (99.0%) and allowed to stand for 15 min at 30 °C. After incubation, 20 µL were injected into the HPLC unit. The CA derivative was detected at 311 nm [9].
For amino acid determination, an analytical column ZORBAX Eclipse AAA-C18 (5 μm, 4.6 x 150 mm) was used a mobile phase gradient elution composed by water, methanol, acetonitrile and NaH2PO4 buffer (40mM and pH 7.8) at conditions 2.0 mL·min-1 and 40 °C. Each sample was derivatized online using o-phthalaldehyde (OPA) for primary amino acids and 9-fluorenyl-methyl-chloroformate (FMOC) for secondary amino acids. The primary and secondary AAs were detected at 338 nm and 262 nm respectively [10].
Phosphate was quantified using a colorimetric method, as described elsewhere [11]. Glycerol was quantified spectrophotometrically [12]. Biomass was determined by calculating the dry cell mass of culture samples [13].
2.3. Analysis of clavulanic acid production
The soy-based culture media that were used for strain selection included ISP, SF and TSB, all containing 15 g·L-1 glycerol. Flask cultures, inoculated with 10% seed medium, were carried out at 28 ºC, pH 6.80 + 0.20 and 250 rpm for 120h. Production rates of biomass and CA, and consumption rates of phosphate and glycerol were calculated in TSB medium supplemented with glycerol. For the remaining soy-based medium studies, only CA was measured. Experiments were performed in biological triplicates.
2.4. Experimental design
A central composite experimental design was carried out with two variables at two levels each (soy protein isolate: 10 and 20 g·L-1; K2HPO4: 0.50 and 2 g·L-1), using CA production (mg·L-1) as the response variable. Assays were run in triplicates. Experimental results were statistically analyzed by using Statgraphics® software (version 6.0).
2.5. Cost analysis
The Net Income associated with the maximum yield of clavulanic acid was calculated based on culture media cost using eq. (1) as follows:

3. Results and discussion
3.1. Analysis of clavulanic acid production
The dynamics of biomass, substrate and product biosynthesis for the strains ScATCC and ScDSM grown in TSB-glycerol are shown in Fig. 1a and 1b. In contrast to the biosynthesis of most secondary metabolites, CA was detectable at very early stages of growth, rather than at the onset of the stationary growth phase. On average, the glycerol uptake rate was 1.80.30 mmol·gDW-1·h-1 and 0.6±0.20 mmol·gDW-1·h-1, for the ScATCC and ScDSM strains, respectively. ScATCC seems to be catabolically more active and due to the lower biomass concentration, it might be more appropriate for CA production. For the ScATCC culture conditions, carbon flux distribution seems to favor product biosynthesis rather than biomass growth.
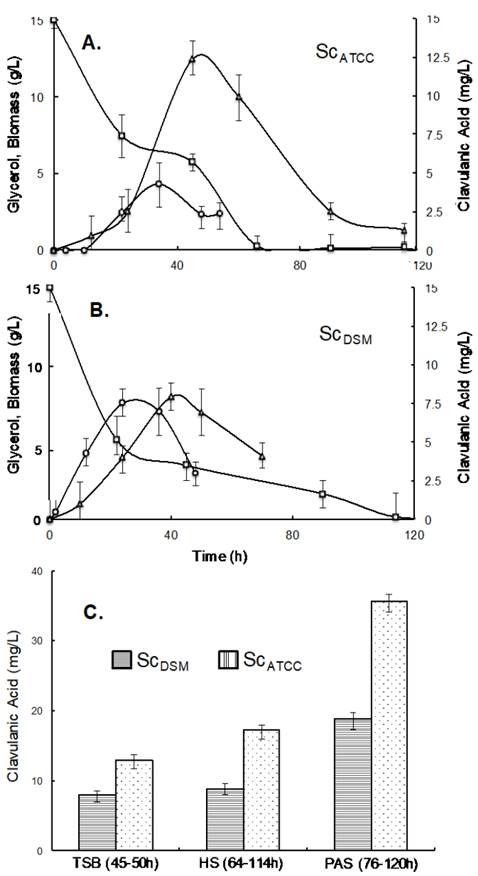
Source: The authors
Figure 1 Clavulanic acid production during Streptomyces clavuligerus batch cultures a. Strain ScATCC grown in TSB supplemented with glycerol. b. Strain ScDSM grown in TSB supplemented with glycerol, (Δ) clavulanic acid, (☐) glycerol, (o) biomass. c. Clavulanic acid production using the strains ScATCC and ScDSM growing in different culture media.
The strains showed different substrate uptake rate profiles. In ScATCC (Fig. 1a), substrate concentration decreased fast up to 30 h of cultivation, when the biomass production slowed down and CA production accelerated. A peak in biomass concentration was observed at approximately 35h, while maximum CA concentration was observed at 42h. After this, the substrate uptake rate increased and glycerol was completely depleted at 70h. For ScDSM (Fig. 1b), maximum substrate uptake rate was sustained up to 30h, decreasing thereafter until complete depletion of glycerol at 120h. While the highest CA concentrations in both strains were detected when substrate uptake rate decreased, product degradation was observed in both strains (see Fig. 1a). ScATCC presented a higher CA production per gram of biomass than ScDSM (2.90 and 1.10 mg·g-1, respectively); this might suggest that the strain ScATCC is more efficient than ScDSM for redirecting metabolic fluxes towards secondary metabolism rather than growth, an important feature in the selection of a CA-producer microorganism.
Strains were also assessed in three soy-based culture media, Tryptic Soy Broth (TSB), Soybean Flour based medium (SF) and Soy Protein Isolate (ISP), to identify the best strain for CA production (Fig. 1c). The highest CA concentration (45.70 mg·L-1) was observed in the strain ScATCC grown in ISP medium. This titer was almost double than that observed for the ScDSM strain in the same medium.
3.2. Culture media for clavulanic acid production by the strain Sc ATCC
Culture media formulations included complex nitrogen sources that provide significant concentrations of amino acids (AAs). Fig. 2 shows the characterization of the amino acid content of complex and semi-synthetic NZ-Amine media, compared with the widely used casamino acids (CSS). As expected, the total AA content was higher in the complex media, particularly in hydrolysed products i.e., ISP (4.20 g·L-1) and NZ-Amine (3.38 g·L-1). In contrast to SF (2.34 g·L-1), containing no hydrolyzed component, CSS provided 4.04 g·L-1 of total amino acids. TSB does not contribute significant amounts of amino acids.

When a complex nitrogen source is employed, the combination of AAs results in a positive effect on CA synthesis, a fact not fully elucidated [14,15]. In the presence of glycerol, AAs derived from aspartate (asparagine and threonine) and alpha-ketoglutarate (glutamine and glutamate), which are present in high concentrations in the ISP, SF and CSS media, have proved to favor CA production [16,17]. The ISP, SF and CSS media are good sources of lysine, which is known to stimulate the production of Cephamycin C rather than CA in Streptomyces clavuligerus.
The production of CA by strain ScATCC was studied in different media (Fig. 3). Overall, complex media showed low CA titers, with the highest production and lowest cost observed in ISP (45.70 mg·L-1). This is a nutrient-rich medium, with a high protein content, which, after hydrolysis, provides the medium with important anabolic precursors for CA biosynthesis, such as amino acids from the aspartate and glutamate families.
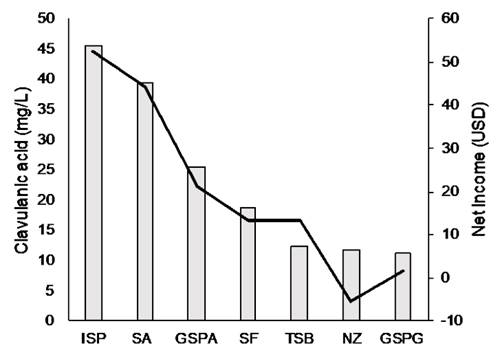
Source: The authors
Figure 3 Culture media and economic analysis for clavulanic acid production in Streptomyces clavuligerus ScATCC.
A low CA concentration (18.80 mg·L-1) was observed in the SF medium, despite its nutrient-rich composition (carbohydrates, lipids, proteins and free amino acids such as lysine, valine, tyrosine, leucine and threonine) [18]. SF is composed of non-hydrolyzed soy flour, which does not provide any additional easy-to-uptake substrates.
The TSB medium rendered a lower CA concentration (12.50 mg·L-1), probably due to its low amino acid content (Fig. 2). TSB has a weak buffering capacity, and the accumulation of CA might have caused the decrease of the pH outside of the favorable range (6.80 - 7.20), affecting S. clavuligerus metabolism and physiology.
In the semi-synthetic medium SA, with starch as the main carbon source, S. clavuligerus cultures showed the second highest CA concentration of all media tested (39.50 mg CA·L-1). Streptomyces species slowly hydrolyze soluble starch releasing maltotriose as the primary product of hydrolysis. This eventually prevents repression and/ or inhibition in β-lactam antibiotic biosynthesis, which is normally triggered by C-sources easily metabolized [19]. Using the NZ-Amine medium a CA production of 11.80 mg·L-1 was achieved. Likewise, the NZ- amine has lysine as the main amino acid source with a high concentration (430 mg·L-1), which favors cephamycin C biosynthesis [20].
S. clavuligerus grown in the chemically defined medium GSPA produced 25.50 mg CA·L-1; this medium contains proline and asparagine which facilitate microbial anabolic activity conducing to increased CA biosynthesis. Besides, maintenance and growth are well supported by two carbon sources, saccharose and glycerol, hence ensuring availability of the C3 precursor, GAP. Conversely, despite having a similar composition to GSPA, the medium GSPG produced the lowest CA concentration of all media tested (11.30 mg·L-1). This concentration represents half of that observed in media containing asparagine, which confirms, once again, the importance of this amino acid, rather than the direct CA precursor arginine, in CA biosynthesis.
In order to test whether CA production is affected by the availability of free amino acids, S. clavuligerus was grown in a medium containing a complex nitrogen source, SF-CSS (non-hydrolysed soybean flour), supplemented with free amino acids (casamino acids-casein hydrolysate). The maximum CA concentration reached was 195 mg·L-1 at 120 h, and the maximum yield was YP/X=18.60 mg·g-1. The difference between yields achieved using the medium non-supplemented and supplemented with free amino acid might be attributed to the presence of extracellular proteases. Streptomyces are heterotrophic microorganisms that can use both complex and simple molecules as nutrients by means of extracellular enzymes induced by complex sources, also related with aerial mycelium formation and sporulation [21].
Table 1 shows the yield coefficients and productivities attained for ScATCC growing in different media. ISP was the most favorable medium for CA biosynthesis (Qp = 0.44 mg·L-1·h-1; YP/X = 5.20 mg·g-1; YP/S = 3,08 mg·g-1), while medium GSPG showed the lowest yield and productivity (Qp = 0.09 mg·L-1·h-1; YP/X = 1.14 mg·g-1; YP/S = 0.76 mg·g-1). These values confirm the different response observed between complex and synthetic media [8,22]. The synthetic media GSPG and GSPA showed production levels similar to published results [5]. In these media, glycerol is the main carbon source and the amino acids glutamate or asparagine and proline are used as nitrogen source. Nonetheless, some reports have shown that the most important amino acids for CA production are ornithine or arginine, rather than glutamate or proline [5].
Table 1 Yield coefficient and productivities attained for clavulanic acid biosynthesis in cultures of Streptomyces clavuligerus, ScATCC
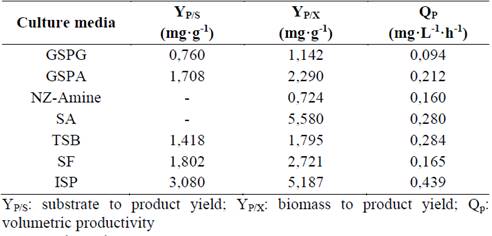
Source: The authors
The production of CA or cephamycin C may be enhanced not only by nutrients in the culture medium but also by intermediates produced along the biosynthetic route [23,24]. Cephamycin C and CA are synthesized by different pathways, which are controlled by intra and extracellular factors through complex regulatory mechanisms. These factors include components of the culture medium and/or intermediate compounds of primary metabolism, which in turn participate directly or indirectly in product biosynthesis [25]. Glycerol, for example, is preferentially used as carbon source forCAproduction [5,26], whereas starch is more suitable for the production of cephamycin C [2,25,27]. The regulation of cephamycin C and CA production are interconnected by the pathway specific regulatory gene Ccar [28].
3.3. Economic analysis
In large-scale production of antibiotics, the composition of the culture medium has the highest impact on process costs [29]. Therefore, any detailed analysis should include an economic evaluation to establish the feasibility of the process. In this work, the net income associated with the maximum yield of CA was calculated based on culture media costs. Fig. 3 shows the net income calculated for all of the media tested. Among the complex media, the use of ISP results in an economically feasible process, with a total gain of 50.5 USD·L-1. SF and TSB exhibited an average total gain of 12 USD·L-1 for the production of CA.
Regarding the semi-synthetic media, SA (Starch-Asparagine medium) proved to be economically feasible, with a total gain of 41 USD·L-1, while NZ-amine showed a net loss of 5 USD·L-1. The chemically defined media GSPA showed an acceptable product concentration, with a very low total gain (18.1 USD·L-1), mainly due to the high cost of its components. Medium GSPG showed a very low gain.
Clavulanic acid production and the net profit were highly favored when complex nitrogen sources were used. In contrast, in the second-best CA production medium, glycerol, the conventional carbon source, was replaced by starch. For the other media, the high cost of individual components such as free amino acids and casein hydrolysate showed to be expensive and unpromising alternatives.
3.4. Effect of the P-source and N-source on clavulanic acid production
The effect of phosphate and nitrogen sources on CA biosynthesis was tested using ISP, the medium showing the highest CA production; for this, a surface response analysis was used, (see Table 2).
Table 2 Effect of the phosphate and nitrogen source on clavulanic acid biosynthesis.

Source: The authors
The results show that the highest concentration of CA (57 mg·L-1) is obtained when the levels of isolated soy protein and phosphate (KH2PO4) were 15 g·L-1 and 2.30 g·L-1, respectively. Decreasing concentrations of P- and N-sources showed a decrease in CA titters, which suggests the role of these components in driving the cell metabolic activities towards CA biosynthesis. The analysis of the effect of phosphate on secondary metabolism shows that the activity of the enzymes, CA synthetase and cephamycin C synthetase, is higher at low concentrations of phosphate, as previously reported [30,31].
The analysis of variance indicates the existence of three statistically significant effects (p<0.05) on CA biosynthesis: the effect of phosphate is highly significant, followed by the interaction ISP-ISP, and the phosphate-phosphate interaction, with an R-squared coefficient of 0.978412. The complete variance analysis is presented in Table 3.
Table 3 Variance Analysis for clavulanic acid biosynthesis in Streptomyces clavuligerus cultures

Source: The authors
In the Pareto chart (Fig. 4a), the left vertical axis represents the significant effect with a 95% confidence interval (phosphate, ISP-ISP and phosphate-phosphate interaction), as indicated by the anova analysis. Phosphate positively affects CA production while the ISP-ISP interaction presents a negative effect. However, high phosphate concentrations (>100 mM) induce biomass growth but somehow inhibit cephamycin C and CA biosynthesis [23,31].
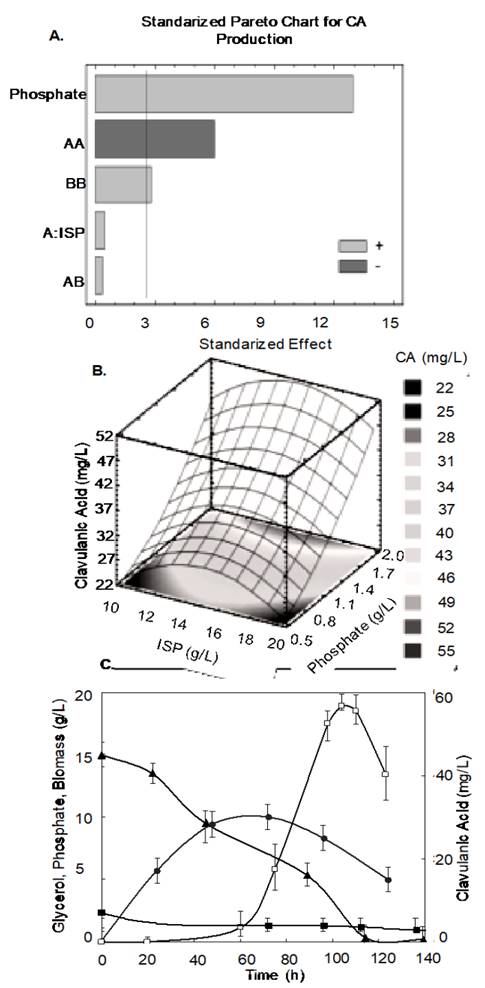
Source: The authors
Figure 4 Effect of phosphate- and nitrogen- sources on CA biosynthesis using the ISP medium by a surface response analysis. a. Standardized Pareto chart for clavulanic acid biosynthesis. b. Response surface diagram for the experimental design aim at studying the effect of isolated soy protein and phosphate concentration on clavulanic acid biosynthesis. c. Experimental validation of the predicted values by the surface methodology approach ([A: ISP] [B: Phosphate]), and their effect on clavulanic acid biosynthesis by ScATCC: (☐) Clavulanic acid, (▲) glycerol; phosphate (◼) and (•) biomass, YX/S (g·g-1) =0.74, YP/S (mg·g-1)=4.32, YP/X(g·g-1)=5.82, QP(mg·L-1·h-1)=0.54, qp(h-1) =0.06, µ (h-1) =0.097.
The combination of variables and their levels, predicted by the surface methodology analysis, to yield the highest CA biosynthesis (phosphate 2.30 g·L-1 and ISP 15.40 g·L-1) is presented in Fig. 4b. These values were experimentally tested, obtaining 56 mg·L-1 of CA at 118h with no residual glycerol (Fig. 4c). Concerning biomass growth, the stationary phase in this culture was achieved at the same time as CA starts to be detectable (75h), following the typical pattern of secondary metabolite biosynthesis. Fig. 4c also shows the kinetic parameters obtained after exploring the predicted values from the surface methodology approach. Both performance and productivity increased after the adjustment of the culture medium, though the final CA concentration is not comparable with values from literature. Besides, it was observed that the nitrogen source greatly influences the final product concentration, while phosphate is limiting for secondary metabolism in S. clavuligerus [3].
Overall, it is clear that the soybean-based media favors CA biosynthesis due to the high content of amino acids known to be precursors of CA biosynthesis [22]. Our results also suggest that the ISP culture medium components induce the formation of extracellular hydrolases and proteases in S. clavuligerus, forcing the bacterium to further degrade the protein during growth, thus ensuring a constant supply of nutrients, essential for primary and secondary metabolism. The use of a non-hydrolyzed nitrogen complex source (soybean flour), supplemented with free amino acids, suggests that proteolytic activity might have been insufficient to release the required nitrogen precursors from the components of flour, resulting in CA titers lower than those achieved in media containing free amino acids.
4. Conclusions
Our results clearly show that the strain exhibiting the best metabolic capabilities was ScATCC. This strain has been widely used in medium optimization studies. Medium optimization is an essential part of process development for microbial fermentation products, as it enables cost reduction by developing culture media with increased productivity and confers ability to use inexpensive raw materials.
Our results showed that the complex culture media providing the most favorable conditions for CA production was ISP, a soybean protein-based media, possibly due to its high free amino acid content. In that medium, the maximum concentration observed was 45.70 mg/L. Medium SF was not effective for CA production (18.8 mg/L). The evaluation of chemically defined media showed that the carbon source, along with appropriate amino acid sources, favors the production of CA. In the case of media GSPG and GSPA, replacing glutamate by asparagine enhanced product concentration by 2.1-fold, suggesting that amino acids of the aspartate family such as asparagine, improve the availability of the C5 CA precursor. The semisynthetic medium SA, using starch as the carbon source, favored product biosynthesis; the production of CA is limited mainly by the availability of the C5 precursor rather than the C3. It seems that starch might have favored the accumulation of GAP.
On the other hand, the use of a non-hydrolyzed nitrogen complex source (soybean flour), supplemented with free amino acids, suggests that the protease activity might have been insufficient to degrade the components of flour, thus rendering lower CA titers compared to those achieved when free amino acids were present.
In the optimization studies, ISP medium showed an increase of CA production by 11 mg/L. The presence of phosphate, in the end, remained in the margins of restrictive concentration thus favoring antibiotic biosynthesis.

