










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Ciencias Químico - Farmacéuticas
Print version ISSN 0034-7418On-line version ISSN 1909-6356
Rev. colomb. cienc. quim. farm. vol.36 no.1 Bogotá Jan./June 2007
Artículo de revisión
Modelos en desarrollo de terapias derivadas de biotecnología
Models on the development of therapies derived from biotechnology
Gustavo Jaimes Monroy1
1 Grupo de investigación, Facultad de Medicina, Fundación Universitaria San Martín, Bogotá, D.C., Colombia. gjaimesm@unal.edu.co
Recibido para evaluación: 19 de febrero de 2007 Aceptado para publicación: 28 de mayo de 2007
RESUMEN
El objetivo de la terapia génica para el tratamiento del cáncer es eliminar las células tumorales preservando el tejido normal. Diversas estrategias para llevar acabo este objetivo han demostrado resultados promisorios en el laboratorio, incluyendo moléculas que pueden tener blancos específicamente predeterminados en secuencias en el ADN y el ARNm; estas secuencias proporcionan un medio para modular la función de genes endógenos. Diferentes tipos de moléculas: inductores de hélices triples, oligodeoxinucleótidos antisentido, ribozimas, ARN de interferencia, aptámeros y señuelos son revisados en este documento.
Palabras clave: ARN de interferencia, terapia antisentido, ribozimas, aptámeros, oligonucleótidos formadores de hélices triples, señuelos.
SUMMARY
The objective of gene therapy for the treatment of cancer is to kill tumor cells but preserve normal tissue. A variety of strategies to achieve these aims have demonstrated promising results in the laboratory, including molecules that can specifically target pre-determined sequences in the DNA or RNAm; that sequences provide a means to modulate endogenous gene function. The different types of molecules: triplex forming molecules, antisense oligodeoxynucleotides, ribozymes, RNA interference, aptamers and decoys, are reviewed in this paper.
Key words: Interference RNA, antisense therapy, ribozymes, aptamers, oligonucleotides triplex helix formers, decoys.
INTRODUCCIÓN
Al analizar el potencial de la biotecnología en el desarrollo de terapias mediante los procesos de la ciencia, se le puede considerar a ésta como un campo a través del cual se producen nuevos beneficios para el tratamiento de múltiples patologías. En las últimas décadas se han apreciado avances muy importantes en el conocimiento de las estructuras y de los procesos que regulan la vida de las células, lo que ha conducido a diseñar y desarrollar mejores y más eficaces fármacos, mejores tratamientos, además de avances importantes en la detección y la prevención de múltiples enfermedades. Todo esto es producto de la investigación jalonada por los avances en clonación, análisis genéticos (geonómica y proteómica) y las técnicas modernas de terapia genética (1). La biotecnología centrada en los procesos de mejoramiento de la salud humana, se constituye en uno de los pilares más importantes, ya que sus productos repercuten directamente en el bienestar social, económico, legislativo y ético (2-4).
Uno de los problemas al abordar la revisión de un tema como éste, es poder considerar la gran cantidad de avances realizados en el desarrollo de herramientas para la terapéutica moderna, sin dejar de lado algunos temas que podrían parecer importantes para algunos; sin embargo, corriendo el riesgo de omitir aspectos que pueden parecer muy importantes, esta revisión está centrada en los aspectos derivados de la biotecnología involucrados en el tratamiento del cáncer, que tienen que ver principalmente con ARN de interferencia, oligonucleótidos antisentido, ribozimas, señuelos y aptámeros.
EL CÁNCER
El cáncer es en realidad un grupo grande de enfermedades, que comparten características como el daño en la regulación del ciclo celular y la interrupción o bloqueo de las vías que llevan a la célula a morir. Estos hechos son producto de la acumulación de alteraciones genéticas y epigenéticas, que terminan afectando proteínas fundamentales en estos procesos generando el fenotipo inmortal (5-8). Desde esta perspectiva no puede verse el cáncer como un proceso estático, sino más bien como un evento dinámico compuesto de múltiples sistemas de señalización, tendientes a mantener el statu quo. Encontrar o diseñar modelos que permitan evaluar estas vías útiles para la célula tumoral, y evaluar la posibilidad de intervenirlas, son dos tareas fundamentales de la biotecnología y de la investigación en ciencias farmacéuticas, con el ánimo de ampliar las alternativas de tratamiento farmacológico.
Las estadísticas de cáncer en el mundo para el año 2000, revelaron que más de 10 millones de personas sufrieron de algún tipo de tumor y se cree que para el año 2020 esta cifra será superior a los 18 millones de personas y que al menos el 60% de estos casos se presentarán en los países en vías de desarrollo (9). En Colombia en el año 2001, por ejemplo, al Instituto Nacional de Cancerología, centro hospitalario de referencia, llegaron 4.389 casos nuevos de cáncer, 333 más que en el año anterior. La incidencia de cáncer en Colombia, calculada en 2004 para todos los tipos de cáncer, fue de 223,2/100.000 en hombres y de 212,9/100.000 en mujeres. Cerca de 28.000 hombres y mujeres mueren anualmente por cáncer y la mayor parte de los casos se diagnostican cuando la enfermedad está muy avanzada (10-14). En 1960, de cada cien muertes que ocurrían en el país, aproximadamente cuatro eran ocasionadas por el cáncer; para el año 2000 este número se incrementó a cerca de 15. Las primeras cuatro localizaciones de cáncer en hombres fueron, en orden decreciente: la próstata, el estómago, el pulmón y el colon/recto, y en las mujeres, las principales localizaciones fueron: el cuello uterino, la mama, el estómago y el colon/recto (13-15).
El aumento de personas afectadas en el mundo por estos procesos, sumados al avance en el conocimiento de los mecanismos de la célula tumoral para mantener su fenotipo, han impulsado la búsqueda de estrategias terapéuticas que conduzcan al encuentro de nuevos fármacos para el tratamiento. La biotecnología ha desarrollado desde hace varias décadas, estrategias altamente específicas, algunas de las cuales se encuentran en fases clínicas de desarrollo y serán la base de este documento en los siguientes puntos.
TERAPIAS DERIVADAS DE LA BIOTECNOLOGÍA
El ARN de interferencia (ARNi)
Fire y colaboradores en 1998, reportaron el silenciamiento postranscripcional de genes específicos luego de la introducción de los ARN de cadenas dobles largas con secuencias idénticas a los genes blanco en el gusano Caenorhabditis elegans (16). Estudios posteriores los señalaron como los ARN de interferencia, que corresponden a secuencias de los ARN de menos de 30 nucleótidos de longitud, que suprimen la expresión de genes a través de complementariedad en sus secuencias. Pertenecen a un mecanismo secuencia específico de control postranscripcional, evolutivamente conservado en plantas y animales. Además de ser un sistema de regulación de la expresión de genes críticos para la vida celular, son un dispositivo de defensa contra ácidos nucleicos foráneos u oportunistas, como los de los virus y transposones (17).
En las células, el ARNdc (largo de doble cadena) se corta inicialmente en un fenómeno dependiente de ATP, en pequeños segmentos de aproximadamente 20 nucleótidos (nt), denominados ARNip (interferencia pequeño), por acción de Dicer, una enzima de la familia RNasa III. Estos ARNip se incorporan a un complejo denominado de "silenciamiento inducido por ARN (RISC); en este complejo, el ARNip se despliega (fenómeno ATP dependiente) y genera un ARN activo que efectúa finalmente el silenciamiento, fragmentando los ARNm diana (Figura 1). RISC es una endonucleasa que contiene proteasas argonautas y cadenas de ARN guía de una sola hebra. En una secuencia de 21 nucleótidos dúplex sobre el ARNm diana, el corte se realiza entre los nucleótidos 10 y 11 contando desde el extremo 5' de la cadena antisentido, corriente arriba. La especificidad del proceso se da por el grado de apareamiento entre el ARNip y la secuencia del ARNm (17-19).
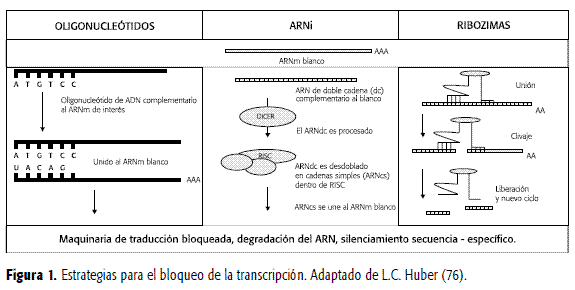
Cada cadena de ARNip, es un dúplex de aproximadamente 20 nucleótidos de longitud, diseñado con dos nucleótidos que sobresalen en el extremo 3' de cada cadena, tomando como modelo los sistemas celulares. Las limitaciones de las secuencias derivan de los métodos de síntesis que regularmente se realizan por dos vías, química y de vectores de expresión, que utilizan polimerasa III. Las observaciones para los diseños están basadas en los sistemas naturales de las células y que implican, tamaño, contenido de G/C de alrededor de 50% y evitan la presencia de tres o más purinas en línea. Los sitios activos de los dúplex no son predecibles, existen estudios que han intentado correlacionar la forma con la función de estos ARN, pero aún quedan muchas consideraciones por indagar; parece que la actividad no sólo es dependiente de la estructura, sino además de factores como competencia por la unión a factores proteicos; estabilidad termodinámica, principalmente en el extremo 5'; dependencia de la secuencia con el complejo RISC, entre otras (18).
Todas las células poseen la maquinaria del ARNi y prácticamente cualquier gen puede ser una diana de este sistema de regulación postranscripcional. El mecanismo ha sido entendido como una herramienta de gran poder en el silenciamiento de genes desde el punto de vista terapéutico. Es una terapia que se considera incluso más efectiva que otras como la antisentido y la de ribozimas. Se han realizado estudios que han establecido que el ARNi puede ser 100 a 1.000 veces más eficiente que la técnica antisentido. La razón, aunque aún en estudio, parece estar relacionada con la actividad del ARNi, puesto que una sola molécula es capaz de clivar múltiples moléculas del ARNm (17).
Con las consideraciones anteriores, es cuestión de tiempo la exploración del potencial terapéutico de los ARNip y sus precursores en el tratamiento de enfermedades; se han implementado varias estrategias para utilizar estas moléculas como fármacos administrados en complejos diseñados para la liberación intracelular y utilizando la expresión de precursores del ARNi a partir de vectores virales. Algunas de estas estrategias se encuentran actualmente en estudios de clínica fase 1 (17, 18).
El empleo del ARNip se convierte en una excelente herramienta para el tratamiento del cáncer. Esta técnica permite actuar sobre secuencias blanco del ARNm de oncogenes alterados en los procesos neoplásicos, realizando el papel de supresores tumorales reguladores (20, 21). Otra potencial utilidad se evidencia en el bloqueo de la expresión de genes que regulan el transporte de fármacos al interior y exterior de las células y que tienen que ver con fenotipos de resistencia a quimio y radioterapia, como la familia de resistencia a múltiples fármacos (22), in vivo ha sido probado en leucemia con alteraciones BCR-ABL (20), en cáncer de ovario contra CD147 que promueve invasión y metástasis en ratones atímicos desnudos (23), en cáncer de seno contra ErbB2 (24), en cáncer de páncreas contra K-Ras (21). Estos trabajos permiten evidenciar que una buena parte de la esperanza para encontrar nuevas alternativas de tratamiento, están siendo estructuradas sobre esta terapia y que el camino a la aplicación de la estrategia del ARNi como un fármaco efectivo contra algún tipo de tumor está aún por explorar en muchos aspectos.
Tecnología antisentido
Esta tecnología, desarrollada en la década de los años ochenta, se consideró como alternativa molecular para el bloqueo de la traducción del ARNm, evitando de esta manera la expresión del producto génico. Se realiza utilizando a manera de fármaco, una secuencia pequeña de nucleótidos que hibrida de manera irreversible con cadenas del ARNm, que son su blanco en el citoplasma; la base de su actividad es la complementariedad de bases entre el antisentido y el ARNm diana (Figura 1) (25). Este concepto, con más de veinte años de desarrollo, llega al año 2000 con productos en estudios de fase clínica 1; tal es el caso de G3139, un antisentido útil en linfomas no Hodgkin, por hibridación con Bcl-2, señalado como el responsable de la resistencia refractaria de estos tumores a la quimioterapia y la radioterapia convencional (26).
Los antisentidos son diseñados de acuerdo con la complementariedad con los ARNm dianas; corresponden a un fragmento de una sola hebra, compuestos usualmente por unidades de ADN, a diferencia de la tecnología del ARNi, y no activan la maquinaria propia de este sistema. El resultado es el bloqueo de la traducción por impedimento estérico de la unión al ribosoma o la interrupción del proceso de elongación; sin embargo, el efecto más potente y esperado para fines terapéuticos, sucede cuando se forma un heterodúplex ADN/ARN con el oligonucleótido. Esta situación, activa las endonucleasas ARNasa H o ARNasa L que, finalmente, rompen el complejo formado. Las ARNasas H y L son ubicuas y se desconocen las secuencias que reconocen; sin embargo, parece que oligos pequeños de ADN, son capaces de activarlas (27).
Para el tratamiento antitumoral se han presentado algunos inconvenientes a pesar de lo particularmente buena que puede ser la tecnología antisentido, una es la insuficiente liberación de fragmentos activos en el tejido tumoral. Se han utilizado liposomas para evadir este inconveniente y otras técnicas similares. Otro es que las células tumorales tienen múltiples vías, tendientes a mantener el fenotipo, las respuestas son de hecho redundantes, lo que deja corta la terapia en muchos casos. Sin embargo, es posible actuar sobre genes específicos claves para la célula tumoral, a través de los antisentidos, como se ha demostrado con Bcl2, para inducir apoptosis en varios tipos tumorales (27).
Las modificaciones que se han realizado a los antisentidos, para mejorar su farmacocinética, la estabilidad frente al ataque de endonucleasas y su actividad, permiten clasificarlas de acuerdo con los cambios efectuados en diferentes etapas del desarrollo. La primera generación correspondió a los oligonucleótidos fosforotioatos, moléculas modificadas en el esqueleto carbonado en donde el oxígeno es cambiado por un azufre; esto les confirió mayor estabilidad en suero, resistencia a la actividad de nucleasas, aumento de la hidrofobicidad, mejorando el paso a través de membranas, manteniendo una alta afinidad de unión al ARNm blanco e induciendo la activación de ARNasa H. La segunda generación introdujo cambios en las piezas de la estructura en las que se incluyó ARN -2'- O - alquil y los grupos O–metil (OMe) y O–(2'- metoxi) etil (MOE), que resultaron con mayor eficiencia. En la tercera generación se introdujo un puente entre 2'- O, 4' - C–metileno produciendo una estructura con la ribosa más o menos encerrada (LNA del inglés locked nucleic acids), en una configuración C3'- endo que potencia de una manera importante la afinidad de unión al ARNm; los copolímeros formados DNA/LNA no se degradan fácilmente ni en suero ni con extractos celulares, mantienen la activación de ARNasa H y se discute si resulta menos tóxico que los fosforotioatos (28-30). Se han estudiado otras modificaciones estructurales, como la de cintas con giros en los extremos con múltiples secuencias antisentido conectadas por una cadena corta entre los dos giros, son llamados oligos RiA (del inglés ribbons antisense); son estables aún después de 24 horas en cultivos in vitro en presencia de exonucleasa III y con una eficiencia en el silenciamiento que aparentemente no es muy afectada (31).
Por último, es importante observar cómo esta técnica ha recibido un nuevo impulso, en la actualidad; debido a que pueden ser elaborados de una manera relativamente fácil y rápida, se pueden tener buenas cantidades de oligos para utilizarlos en investigación para validación de blancos en cáncer o para la búsqueda de un potencial fármaco. El talón de Aquiles sigue siendo cómo llevar eficientemente los oligos a los tejidos tumorales y en la concentración precisa en experiencias in vivo (30, 32, 33).
En cáncer, su utilidad ha sido importante cuando se ha probado en conjunto con la terapia corriente. Los blancos habitualmente han sido genes que codifican proteínas para sobrevida, diferenciación, angiogénesis y apoptosis (34). Tienen dos ventajas importantes: pueden fácilmente hacer blanco en genes intracelulares que sería imposible de alcanzar con otras estrategias como las de macromoléculas de anticuerpos, y la baja toxicidad por corresponder a mecanismos que las células utilizan para regular sus genes o bloquear ácidos nucleicos foráneos. Los estudios clínicos tienen limitaciones debido a los aspectos ya expuestos; sin embargo, el potencial es amplio y aún contin&uac ute;an realizándose ensayos de esta naturaleza para probar su potencial antitumoral en el control de algunos genes implicados en el fenotipo tumoral (27).
Ribozimas
Las ribozimas son moléculas de ARN con actividad catalítica. En 1981 un grupo de investigadores las descubrió mientras trabajaban con reacciones de splicing en Tetrahymena thermophyla, y descubrieron que ciertas reacciones de éstas eran catalizadas por el ARN. La estrategia ofrece algunas ventajas sobre otras como la de antisentidos, ya que su actividad catalítica, además de específica, puede ejercerla sobre múltiples moléculas blanco de ARNm. Se han descrito alrededor de seis tipos de ribozimas; sin embargo, para efectos terapéuticos, la más importante es la cabeza de martillo (35).
Las ribozimas incluyen los intrones del grupo I y del grupo II, la subunidad ARN de la ARNasa P, ribozimas de horquilla del virus de la hepatitis delta, ribozimas ribosomales y las ribozimas cabeza de martillo (35). La ribozima cabeza de martillo fue descubierta en viroides y está involucrada en el procesamiento de mensajes policistrónicos, y tiene aproximadamente 40 nucleótidos de longitud. De forma natural actúa en cis durante la replicación viral, pero se puede producir una ribozima cabeza de martillo que actúe en forma trans sobre otras moléculas de ARNm (30). La estructura secundaria de la ribozima cabeza de martillo consiste de dos secciones en hélice antisentido, denominadas hélices I y III, y un dominio catalítico (dependiente de cationes divalentes) adyacente, así como una sección en asa (36). La ribozima y su sustrato hibridan a través de los tallos I y III. Corta eficientemente sustratos con secuencias tripletes AUC, GUC, GUA, GUU, UUC y CUC, es decir, secuencias NUH en donde H es cualquier nucleótido diferente de guanosina; la cinética enzimática muestra mejor eficiencia sobre los tripletes AUC y GUC que sobre los demás, así como preferencia en sustratos cortos de 25 bases y baja eficiencia sobre sustratos de 200 a 1.000 bases. En teoría, la ribozima cabeza de martillo puede ser rediseñada para cortar cualquier ARNm. La especificidad del corte y su eficiencia vienen determinada por los brazos de unión I y III. El corte es una reacción de trans esterificación, en la cual reacciona el extremo 5'-hidroxilo y el extremo 2', 3'-fosfato cíclico en el sustrato nucleotídico. Ésta es una de las enzimas DEL ARN más pequeñas conocidas; en estudios in vitro, ribozimas anti-ras se han utilizado para discriminar secuencias mutadas en el oncogen H-ras en el codón 12 y en otro estudio utilizando ribozimas se logró actuar sobre genes defectuosos sin afectar el gen normal, en líneas celulares de vejiga (30, 35). Los blancos potenciales para la degradación mediada por ribozimas incluyen proteínas virales, reguladoras del ciclo celular, factores de transcripción y proteínas de fusión aberrantes. Existen algunos productos en la industria farmacéutica que utilizan ribozimas: en fase clínica I Herzyme®, cuyo blanco es HER-2 que se encuentra alterado en varios procesos tumorales, y Angiozyme®, en fase clínica II, con blanco en VEGF-R 1 para cáncer colorrectal metastásico (30).
El mayor reto para el uso de esta técnica in vivo, es mantener la eficiencia catalítica de las ribozimas en un ambiente celular donde ocurren complejas interacciones moleculares. Se han empleado para su transporte al interior celular, plásmidos, con el promotor de la polimerasa III y la estructura tallo-giro, que estabiliza la ribozima. Debido a la actividad de las nucleasas de los sistemas biológicos, las ribozimas presintetizadas han tenido que ser modificadas para su protección al ataque nucleofílico luego de ser usadas in vitro o in vivo. Se han utilizado varias estrategias de modificación, pues las más efectivas son la incorporación de una timidina invertida en el extremo 3' y la incorporación de una uridina 2'- C – alil; algunos de estos cambios han logrado aumentar el tiempo de vida media de unos pocos minutos hasta diez días en ensayos in vitro (30, 37).
Oligonucleótidos formadores de triples hélices (tríplex ADN)
Son secuencias que hibridizan con el ADN de doble hebra, lo que resulta en la formación de secuencias de triple hélice que terminan bloqueando la transcripción del gen diana, básicamente por el bloqueo de las secuencias de unión de las proteínas activadoras de la transcripción.
Los oligonucleótidos formadores de triples hélices (OFT) se pueden formar in vivo por unión a oligonucleótidos a continuación de purinas cercanas o pirimidinas o como una parte de H-ADN, el cual contiene regiones plegadas de hélices triples. Esta estrategia aprovecha la frecuencia importante de sitios de polipurinas o polipirimidinas en el genoma humano. Para la estabilización de estas estructuras, las células utilizan cationes divalentes; in vitro se han utilizado moléculas como espermina e incluso intercaladores del ADN como hidralazina unidos a las estructuras (38).
Se reconocen principalmente dos motivos estables de los OFT; difieren, básicamente, en la orientación y en la composición de las bases de la tercera cadena. En el motivo paralelo de pirimidinas, la formación triple de oligonucleótido homopirimidina está orientada en paralelo a la cadena dúplex de purinas formando un puente H de Hoogsteen con las bases C+-GC y T-AT en tripletes. El modelo antiparalelo o motivo de purina está caracterizado por tripletes G-GC, A-AT y T-AT Hoogsteen inversos entre una cadena OFT rica en purinas y el ADN dúplex. Para ambos motivos la tercera cadena se une a la hendidura mayor del ADN y requiere un segmento de secuencia blanco homopurina - homopirimidina, idealmente de 15 a 30 nt. Para el motivo paralelo la restricción es la sensibilidad al pH del triplete C+-GC, el cual es inestable bajo condiciones fisiológicas debido al bajo pKa de la tercera cadena de citosinas. La estabilidad del OFT en el motivo antiparalelo es fuertemente dependiente de la secuencia (39, 40).
Los OFT pueden unirse selectivamente a una secuencia del genoma, de doble cadena interfiriendo con la transcripción y la maquinaria de procesamiento del ADN por vía de formación de una hélice triple sin necesidad de abrir la molécula. Esta estrategia tiene varias ventajas sobre otras como la de antisentido, ya que se necesitan sólo dos copias por cada alelo para realizar un bloqueo específico, mientras que con antisentidos se requieren varios cientos de copias, para causar sus efectos biológicos; además, su utilidad adicional de permitir knockout de genes y también mutagénesis dirigida contra blancos (gracias a la posibilidad de asociarlos a moléculas que cortan de manera específica secuencias), recombinación dirigida de blancos y manipulación genómica de genes diana, hacen que esta herramienta biotecnológica se tenga en cuenta actualmente como estrategia antitumoral (41-43).
Se ha demostrado que oligonucleótidos formadores de triples hélices pueden unirse a p53 de una manera sitio-específica por homología, inhibiendo la proliferación de células en cáncer de colon in vitro y en ratones atímicos desnudos inyectados por vía subcutánea con células tumorales humanas de carcinoma de colon HCT 116 (44). En otro ensayo, realizado sobre el gen Ets2, un factor de transcripción, que tiene un papel importante en la promoción crecimiento y sobrevida en cáncer de próstata y seno, se utilizó una secuencia OFT rica en purinas sobre células de cáncer de próstata con un alto grado de especificidad y de selectividad, que resultaron en la regulación del ciclo celular e inducción de apoptosis (45).
En la última década se han incrementado los esfuerzos para desarrollar nuevas formas de OFT que tengan mejor estabilidad y reconozcan más secuencias de las habituales; por ejemplo, se han desarrollado nuevos OFT, llamados BNA (del inglés bridged nucleic acids) y se ha encontrado que las modificaciones introducidas promueven fuertemente la forma paralela bajo condiciones fisiológicas (46, 47).
Oligonucleótidos de doble cadena con elementos cis (señuelos)
Corresponden a pequeños fragmentos de ADN de doble cadena que contienen secuencias cis con alta afinidad para un factor de transcripción. Estos fragmentos de ADN generalmente no superan 25 pb; pueden ser introducidos en una célula blanco como señuelos capaces de "atrapar un factor de transcripción, antes de unirse a la secuencia blanco en una región promotora del ADN celular con la consecuente activación de la transcripción de genes, lo que corresponde a una verdadera interacción cis - trans. El factor de transcripción es retenido en el citoplasma regulando de esta manera la expresión de genes (48, 49).
Los señuelos se han utilizado in vitro e in vivo como herramienta biotecnológica de interrupción de la expresión de genes tanto en la transcripción como en la traducción. Esta interacción puede significar un decrecimiento de la tasa de transcripción al interactuar con la ARN polimerasa; de hecho, un solo factor puede llegar a afectar la transcripción de varios genes (50).
En 1996, la Food and Drug Administration (FDA) aprobó la aplicación clínica de señuelos contra E2F para el tratamiento de hiperplasia de la neoíntima en desviaciones coronarias venosas (48). En cáncer, su uso terapéutico ha sido bien acogido gracias a la facilidad para identificar los factores de transcripción blanco en múltiples tumores, la síntesis relativamente simple y rápida; además, no requiere que se conozcan las estructuras exactas de los factores de transcripción blanco de la terapia. Algunos sugieren que puede llegar a ser más efectivo que los antisentidos. Ha demostrado ser eficaz en varios tumores contra E2F, NFkß, Stat3 y CRE en tumores de seno, próstata, melanoma y otros (51-55).
La utilización in vitro de terapias conjugadas moleculares con las terapias convencionales está siendo explorada y es una buena aproximación para combinar la potencia de estas herramientas biotecnológicas con la quimioterapia actual. En 2004, Liu y colaboradores trataron con un oligonucleótido señuelo contra CRE líneas celulares de carcinoma colorrectal en conjunción con terapia convencional obteniendo resultados promisorios (56).Los señuelos tienen una actividad importante sobre factores que se activan en citoplasma; sin embargo, aquellos que lo hacen en núcleo o que son rápidamente transportados al núcleo no son atrapados, lo que se constituye en una limitante de su actividad (57). Las modificaciones para mejorar su actividad terapéutica incluyen, básicamente, las mismas que han sido empleadas para otro tipo de terapias con oligonucleótidos, e incluyen cambios en el esqueleto carbonado o en las bases o incorporación de otras moléculas que mejoren sus características (58). Actualmente se realizan modificaciones a los señuelos para mejorar su potencial y utilización como fármacos; una de las más promisorias incluye los "señuelos quimera que contienen ADN y estructuras PNAs (del inglés peptide nucleic acids), en donde los esqueletos carbonados son reemplazados por unidades de glicina N-(2-amionoetil). Son estructuras PNA-ADN-PNA quimeras. Se han utilizado contra NFkß (59), Myc (60), y más reciente fue construido y utilizado un señuelo quimera contra p53, en donde se ha adicionado una secuencia de localización celular para su transporte al núcleo, una secuencia para facilitar el paso por las membranas y una secuencia para facilitar su purificación (61).
Aptámeros
En los años ochenta se realizaron observaciones de algunos procesos virales, en donde se encontraba que pequeñas moléculas de ARN se unían a algunas proteínas virales como mecanismo de regulación o para inhibir las proteínas de respuesta antiviral en la célula huésped. Posteriormente, aparecieron estudios que informaban que un ARN aptámero podía ser utilizado para inhibir la actividad de una proteína en un proceso patológico. La proteína blanco correspondía a una proteína de replicación del virus del VIH (62). Demostraron que a partir de librerías de los ARN era posible seleccionar estos pequeños ARN de alta afinidad de unión en un proceso denominado SELEX (del inglés systematic evolution of ligands by exponential enrichment) (62-64). El proceso se inicia por la generación de una librería de secuencias al azar de los ARN, usualmente estas librerías contienen entre 1014 y 1015 especies diferentes de ARN, que se pliegan en dos estructuras diferentes dependiendo de su secuencia particular. La librería es incubada con la proteína de interés y los ARN presentes en la librería que se unen a la proteína son separados de los que no lo están. Luego estos ARN son amplificados por RT-PCR y transcritos in vitro para generar un grupo de ARN enriquecidos con las proteínas de interés. Esta selección y amplificación se repite de 8 a 12 veces. Los aptámeros seleccionados son clonados y secuenciados (62, 65). Tienen grandes ventajas sobre otro tipo de terapias que actúan sobre proteínas, por ejemplo, los aptámeros pueden unirse de manera específica y fuertemente (Kd del orden de nanomolar y picomolar) a biopolímeros blanco; no se ha observado desarrollo de resistencia contra ellos; su pequeño tamaño los hace fáciles de sintetizar o modificar para mejorar sus propiedades farmacológicas y no han demostrado inmunogenicidad (66).En su mayoría no presentan conformaciones definidas en solución, pero en presencia del ligando se producen una serie de plegamientos alrededor del mismo, que pasa a ser parte de la estructura del ácido nucleico (67).
Los aptámeros han sido sometidos a modificaciones estructurales que mejoran su actividad y los hacen más estables, por ejemplo, al diseñar formas circulares protegidas estos aptámeros pueden durar más de 10 horas en suero, sin sufrir ninguna alteración (68, 69). Su uso en cáncer no ha sido suficientemente explorado, probablemente por la dificultad para construirlos y seleccionarlos comparado con otras técnicas como las de señuelos o la antisentido. Actualmente se encuentran en evaluación aptámeros para la inhibición de la replicación del VIH (natural), angiogénesis (natural) e hiperplasia de la íntima (generado in vitro). La FDA ha aprobado el uso de Macugen®, un aptámero terapéutico dirigido contra el factor de crecimiento epitelial vascular en enfermedad macular (66).
Por otro lado, se encuentran también en desarrollo terapias basadas en vacunas en donde se utilizan células dendríticas con objeto de estimular la respuesta humoral y la respuesta inmune citotóxica; este tipo de estrategias se ha utilizado con éxito en el tratamiento de melanoma y cáncer renal, leucemias, próstata y otros (70, 71). En general, se puede asumir que todos estos esfuerzos en investigación y desarrollo abordan los mecanismos alterados en las transformaciones tumorales, los cuales son más o menos generales y conciernen los procesos de control del ciclo celular, la reparación, la evasión de la muerte celular, la expresión de la maquinaria necesaria para invadir y migrar, y las señales necesarias para escapar a los controles del organismo, además de otros no menos importantes, como la producción de proteínas para evadir terapias e intervenciones. Visto de esta manera, todo este conjunto complejo de estrategias que mantienen el fenotipo de la enfermedad, puede ser ubicado en proteínas receptoras, proteínas intermediarias de señalización y factores de transcripción que inducen la expresión de genes necesarios para el mantenimiento del mismo fenotipo. Por supuesto, el mecanismo real no es tan simple, cada tumor tiene su propio repertorio de proteínas y cascadas de señalización que hacen de este conjunto de enfermedades un proceso dinámico, múltiple y complejo. Los avances en patología molecular continuamente develan los mecanismos y recuperan los modelos, las proteínas claves para cada tipo tumoral (72-74). El entendimiento de estos procesos es indispensable para el adelanto de nuevas estrategias de tratamiento derivadas de la biotecnología, así como el perfeccionamiento de ensayos que permitan llegar de una manera efectiva, al tratamiento de pacientes afectados que son la verdadera razón de su desarrollo.
BIBLIOGRAFÍA
1. D.D.Y. Ryu y D.H. Nam, Biomolecular engineering: A new frontier in biotechnology, J. Mol. Catalysis B: Enzymatic, 10, 23 (2000). [ Links ]
2. H. Herrera, Actividades de la Organización de los Estados Americanos para el desarrollo de indicadores en biotecnología, ed. Washington: Oficina de Ciencia y Tecnología, comunicación personal, 2004. [ Links ]
3. I. Casal, L. García. J.M. Guisán y J.M. Martínez Zapater, "Biotecnología y salud", ed. SEBIOT, S.E. d. B. España, 2000, p. 56. [ Links ]
4. Comunicación de la comisión de las comunidades europeas al Parlamento Europeo, al comité económico y social y al comité de las regiones. Ciencias de la vida y biotecnología: una estrategia para Europa, Luxemburgo: Comisión Europea, 2002, p. 56. [ Links ]
5. A.G. Knudson, Genetics of human cancer, Genetics, 79, suppl, 305 (1975). [ Links ]
6. C. Garnis, T.P.H Buys y W.L. Lam, Genetic alteration and gene expression modulation during cancer progression, Molecular Cancer, 3, 9 (2004). [ Links ]
7. J. Gray y C. Collins, Genome changes and gene expression in human solid tumors, Carcinogenesis, 21, 443 (2000). [ Links ]
8. S. Knuutila, Y. Aalto, K. Autio, A. Bjorkqvist, W. El-Rifai, S. Hemmer, T. Huhta, E. Kettunen, S. Kiuru-Kuhlefelt, M. Larramendy, T. Lushnikova, O. Monni, H. Pere, J. Tapper, M. Tarkkanen, A. Varis, V. Wasenius, M. Wolf y Y. Zhu, DNA copy number losses in human neoplasms, Am. J. Pathol., 155, 683 (1999). [ Links ]
9. M. Rodríguez, J. del S. García, J. Rosales, J. E. Duque, V. Abello, M. Grajales, M. Gómez, C. Sossa, C. Casas, J.F. Combarías, G. Quintero, A.F. Cardona, M. Daza, L. Enciso y A. Camacho, Consenso colombiano sobre el tratamiento del linfoma folicular no Hodgkin, Revista Colombiana de Cancerología, 9, 149 (2005). [ Links ]
10. M. Piñeros, C. Pardo, L.F. Cantor, G. Hernández, T.M. Martínez, N.P. Almanza, A. Serrano y H. Posso, Registro institucional de cáncer del Instituto Nacional de Cancerología, empresa social del Estado. Resultados principales, año 2001, Revista Colombiana de Cancerología, 6, 4 (2002). [ Links ]
11. C. Pardo, R. Murillo, M. Piñeros y M. A. Castro, Casos nuevos de cáncer en el Instituto Nacional de Cancerología, Colombia, Revista Colombiana de Cancerología, 7, 4 (2003). [ Links ]
12. A. Jemal, C. Tiwari, T. Murray, A. Ghafoor, A. Samuels, E. Ward, E.J. Feuer y M.J. Thun, Cancer statistics, Cancer J. Clin., 54, 8 (2004). [ Links ]
13. M. P. Petersen y R. H. Murillo, Incidencia de cáncer en Colombia: importancia de las fuentes de información en la obtención de cifras estimativas, Revista Colombiana de Cancerología, 8, 5 (2004). [ Links ]
14. Ministerio de la Protección Social, Instituto Nacional de Cancerología, E.S.E., "Anuario estadístico", MediLegis ed., Bogotá, 2005. [ Links ]
15. Ministerio de la Protección Social, Instituto Nacional de Cancerología, E.S.E., "El cáncer. Aspectos básicos sobre su biología, clínica, prevención, diagnóstico y tratamiento", Bogotá, 2004. [ Links ]
16. A. Fire, S. Xu y M.K. Montgomery, Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans, Nature, 391, 806 (1998). [ Links ]
17. D.M. Dykxhoorn, D. Palliser y J. Lieberman, The silent treatment: siRNAs as small molecule drugs, Gene Ther., 13, 541 (2006). [ Links ]
18. M. Manoharan, RNA interference and chemically modified small interfering RNAs, Current Opinion in Chemical Biology, 8,570 (2004). [ Links ]
19. R.L. Juliano, V.R. Dixit, H. Kang, T.Y. Kim, Y. Miyamoto y D. Xu, Epigenetic manipulation of gene expression: A toolkit for cell biologists, J. Cell Biol.,169, 847 (2005). [ Links ]
20. A.L. Gartel y E.S. Kandel, RNA interference in cancer, Biomolecular Engineering, 23, 17 (2006). [ Links ]
21. R.K.M. Leung, y P.A. Whittaker, RNA interference: From gene silencing to gene-specific therapeutics, Pharmacology & Therapeutics, 107, 222 (2005). [ Links ]
22. Y. Huang, y W. Sadee, Membrane transporters and channels in chemoresistance and -sensitivity of tumor cells, Cancer Letters, 239, 168 (2006). [ Links ]
23. W. Zou, H. Yang, X. Hou, W. Zhang, B. Chen y X. Xin, Inhibition of CD147 gene expression via RNA interference reduces tumor cell invasion, tumorigenicity and increases chemosensitivity to paclitaxel in HO-8910pm cells, Cancer Letters, 248, 211 (2007). [ Links ]
24. X. Hu, F. Su, L. Qin, W. Jia, C. Gong, F. Yu, J. Guo y E. Song, Stable RNA interference of ErbB-2 gene synergistic with epirubicin suppresses breast cancer growth in vitro and in vivo, Biochemical and Biophysical Research Communications, 346, 778 (2006). [ Links ]
25. T. C. Stanley y C.F. Bennet, Progress in antisense oligonucleotide therapeutics, Ann. Rev. Pharmacol. Toxicol., 36, 107 (1996). [ Links ]
26. K. Senior, Why antisense technology makes good sense for cancer treatment, Pharmaceutical Science & Technology Today, 3, 217 (2000). [ Links ]
27. V. Wacheck y U. Zangemeister-Wittke, Antisense molecules for targeted cancer therapy, Critical Reviews in Oncology/Hematology, 1, 65 (2006). [ Links ]
28. C. Wahlestedt, P. Salmi, L. Good, J. Kela, T. Johnsson, T. Hôkfelt, C. Broberger, F. Porreca, J. Lai, K. Ren, M. Ossipov, A. Kohkin, N. Jakobsen, J. Skouv, H. Oerum, M.H. Jacobsen y J. Wengel, Potent and nontoxic antisense oligonucleotides containing locked nucleic acids, PNAS, 97, 5633 (2000). [ Links ]
29. M. Matsukura, K. Shinozuka, G. Zon, H. Mitsuya, M. Reitz, J. Cohen y S. Broker, Phosphorothioate analogs of oligodeoxinucleotides: Inhibitors of replication and cytopathic effects of human immunodeficiency virus, Proc. Natl. Acad. Sci. USA, 84, 7706 (1987). [ Links ]
30. J. Kurreck, Antisense technologies: Improvement through novel chemical modifications, Eur. J. Biochem., 270, 1628 (2003). [ Links ]
31. I.J. Moon, K. Choi, Y.K. Choi, J.E. Kim, Y. Lee, A.D. Schreiber y J.G. Park, Potent growth inhibition of leukemic cells by novel ribbon-type antisense oligonucleotides to c-myb1, The Journal of Biological Chemistry, 275, 4647 (2000). [ Links ]
32. T.L.H. Jason, J. Koropatnick y R.W. Berg, Toxicology of antisense therapeutics, Toxicology and Applied Pharmacology, 201, 66 (2004). [ Links ]
33. W. Pawlak, J. Zolnierek, T. Sarosiek y C. Szczylik, Antisense therapy in cancer, Cancer Treatment Reviews, 26, 333 (2000). [ Links ]
34. C. A. Stein, The experimental use of antisense oligonucleotides: a guide for the perplexed, The Journal of Clinical Investigation, 108, 641 (2001). [ Links ]
35. L.D. Curcio, D.Y. Bouffard y K.J. Scanlon, Oligonucleotides as modulators of cancer gene expression, Pharmacology & Therapeutics, 74, 317 (1997). [ Links ]
36. Y. Fichou y C. Ferec, The potential of oligonucleotides for therapeutic applications, Trends in Biotechnology, 24, 563 (2006). [ Links ]
37. S. Schubert, J.P. Furste, D. Werk, H.P. Grunert, H. Zeichhardt, V.A. Erdmann y J. Kurreck, Gaining Target Access for Deoxyribozymes, Journal of Molecular Biology, 339, 355 (2004). [ Links ]
38. T.J. Thomas, J.R. Seibol, L.E. Adams y E.V. Hess, Triplex DNA stabilization by hydralazine and the presence of anti(triplex DNA) antibodies in patients treated with hydralazine, Biochem., 311, 183 (1995). [ Links ]
39. S. Buchini y C.J. Leumann, Dual recognition of a C-G pyrimidine-purine inversion site: Synthesis and binding properties of triplex forming oligonucleotides containing 2´aminoethoxy 5 methyl 1H pyrimidin 2 one ribonucleosides, Tetrahedron Letters, 44, 5065 (2003). [ Links ]
40. S. Buchini y C.J. Leumann, Recent improvements in antigene technology, Current Opinion in Chemical Biology, 7, 717 (2003). [ Links ]
41. D. Praseuth, A.L. Guieysse y C. Helene, Triple helix formation and the antigene strategy for sequence-specific control of gene expression, Biochimica et Biophysica Acta (BBA) - Gene Structure and Expression, 1489, 181 (1999). [ Links ]
42. R.V. Guntaka, B.R. Varma y K.T. Weber, Triplex-forming oligonucleotides as modulators of gene expression, The International Journal of Biochemistry & Cell Biology, 35, 22 (2003). [ Links ]
43. M. M. Seidman y P.M. Glazer, The potencial for gene repair via triple helix formation, J. Clin. Invest., 112, 487 (2003). [ Links ]
44. N.R. Richard, J.L. Cook y J.F. Giardina, The inhibition of tumor growth by triplex-forming oligonucleotides, Cancer Letters, 209, 51 (2004). [ Links ]
45. G.M. Carbone, S. Napoli, A. Valentini, F. Cavalli, D.K. Watson y C.V. Catapano, Triplex DNA-mediated downregulation of Ets2 expression results in growth inhibition and apoptosis in human prostate cancer cells, Nucl. Acids Res., 32, 4358 (2004). [ Links ]
46. S. Obika, T. Uneda, T. Sugimoto, D. Nanbu, T. Minami y T. Doi, 2´-O,4´-C-methylene bridged nucleic acid (2´;, 4´-BNA): Synthesis and triplex-forming properties, Bioorganic & Medicinal Chemistry, 9, 1001 (2001). [ Links ]
47. S. Obika, Development of bridged nucleic acid analogues for antigene technology, Chem. Pharm. Bull., 52, 1399 (2004). [ Links ]
48. R. Morishita, J. Higaki, N. Tomita y T. Ogihara, Application of transcription factor "Decoy" strategy as means of gene therapy and study of gene expression in cardiovascular disease, Circ. Res., 82, 1023 (1998). [ Links ]
49. M.J. Mann y V.J. Dzau, Therapeutic applications of transcription factor decoy oligonucleotides, J. Clin. Invest., 106, 1071 (2000). [ Links ]
50. L. Penolazzi, E. Lambertini, M. Borgatti, R. Piva, M. Cozzani, I. Giovanni, R. Naccari, G. Siciliani y R. Gambari, Decoy oligodeoxynucleotides targeting NF-kappab transcription factors: Induction of apoptosis in human primary osteoclasts, Biochemical Pharmacology, 66, 1189 (2003). [ Links ]
51. J.D. Ahn, C.H. Kim, J. Magae, Y.H. Kim, H.J. Kim, K.K. Park, S. Hong, K.G. Park, I.K. Lee y Y.C. Chang, E2F decoy oligodeoxynucleotides effectively inhibit growth of human tumor cells, Biochemical and Biophysical Research Communications, 310, 1048 (2003). [ Links ]
52. H. Gao, J. Xiao, Q. Sun, H. Lin, Y. Bai, L. Yang, B. Yang, H. Wang y Z. Wang, A single decoy oligodeoxynucleotides targeting multiple oncoproteins produces strong anticancer effects, Mol. Pharmacol., 70, 1621 (2006). [ Links ]
53. R. Morishita, N. Tomita, Y. Kaneda y T. Ogihara, Molecular therapy to inhibit NF[kappa]b activation by transcription factor decoy oligonucleotides, Current Opinion in Pharmacology, 4, 139 (2004). [ Links ]
54. Y.G. Park, M. Nesterova, S. Agrawal y Y.S. Cho-Chung, Dual blockade of cyclic AMP response element- (CRE) and AP-1 directed transcription by CRE-transcription factor decoy oligonucleotide, The Journal of Biological Chemistry, 274, 1572 (1999). [ Links ]
55. B.E. Barton, T.F. Murphy, P. Shu, H.F. Huang, M. Meyenhofen y A. Barton, Novel single-stranded oligonucleotides that inhibit signal transducer and activator of transcription 3 induce apoptosis in vitro and in vivo in prostate cancer cell lines, Mol. Cancer Ther., 3, 1183 (2004). [ Links ]
56. W.M. Liu, K.A. Scott, S. Shahin y D.J. Propper, The in vitro effects of CRE-decoy oligonucleotides in combination with conventional chemotherapy in colorectal cancer cell lines, Eur. J. Biochem., 271, 2773 (2004). [ Links ]
57. A. Bene, R.C. Kurten y T.C. Chambers, Subcellular localization as a limiting factor for utilization of decoy oligonucleotides, Nucl. Acids Res., 32, 142 (2004). [ Links ]
58. S. Agrawal y R.P. Iyer, Modified oligonucleotides as therapeutic and diagnostic agents, Current Opinion in Biotechnology, 6, 12 (1995). [ Links ]
59. R. Gambari, New trends for the development of novel decoy molecules against nuclear factor kappa-b (NF-kb) transcription factor, Minerva Biotec., 16, 145 (2004). [ Links ]
60. S. EL-Andaloussi, H. Johansson, A. Magnusdottir, P. Jarver, P. Lundberg y U. Langel, TP10, a delivery vector for decoy oligonucleotides targeting the Myc protein, Journal of Controlled Release, 110, 189 (2005). [ Links ]
61. M. Sakaguchi, T. Nukui, H. Sonegawa, H. Murata, J. Futami, H. Yamada y N.H. Huh, Targeted disruption of transcriptional regulatory function of p53 by a novel efficient method for introducing a decoy oligonucleotide into nuclei, Nucl. Acids Res., 33, e88 (2005). [ Links ]
62. P.S. Pendergrast, H.N. Marsh, D. Grate, J.M. Healy y M. Stanton, Nucleic acid aptamers for target validation and therapeutic applications, J. Biomol. Tech., 16, 224 (2005). [ Links ]
63. S.M. Nimjee, C.P. Rusconi y B.A. Sullenger, Aptamers: An emerging class of therapeutics, Ann. Rev. Med., 56, 555 (2005). [ Links ]
64. J. Tang, T. Yu, L. Guo, J. Xie, N. Shao y Z. He, In vitro selection of DNA aptamer against abrin toxin and aptamer-based abrin direct detection, Biosensors and Bioelectronics, Artículo en prensa (2006). [ Links ]
65. S.M. Nimjee, C.P. Rusconi, R.A. Harrington y B.A. Sullenger, The potential of aptamers as anticoagulants, Trends in Cardiovascular Medicine, 15, 41 (2005). [ Links ]
66. J.F. Lee, G.M. Stovall y A.D. Ellington, Aptamer therapeutics advance, Current Opinion in Chemical Biology, 10, 282 (2006). [ Links ]
67. N. Piganeau y R. Schroeder, Aptamer structures: A preview into regulatory pathways?, Chemistry & Biology, 10, 103 (2003). [ Links ]
68. D.A. di Giusto y G.C. King, Construction, stability, and activity of multivalent circular anticoagulant aptamers, J. Biol. Chem., 279, 46483 (2004). [ Links ]
69. E.N. Brody y L.Golden, Aptamers as the therapeutic and diagnostic agents, Reviews in Molecular Biotechnology, 74, 5 (2000). [ Links ]
70. J. Bancheeau, F. Briere, C. Caux, J. Davoust, S. Lebeque, Y. Liu, B. Palendrand y K. Palucka, Immunobiology of dendritic cells, Annu. Rev. Immunol., 18, 767 (2000). [ Links ]
71. K.A. Berlyn, S. Ponniah, S.A. Stass, J.G. Malone, G. Hamlin-Green, J.K. Lim, M. Cottler-Fox, G. Tricot, R.B. Alexander, D.L. Mann y R.W. Malone, Development dendritic cell polynucleotide vaccination for prostate cancer immunotherapy, Journal of Biotechnology, 73, 155 (1999). [ Links ]
72. B. Jansen y U. Zangemeister-Wittke, Antisense therapy for cancer--the time of truth, The Lancet Oncology, 3, 672 (2002). [ Links ]
73. F.A. Fleischer, A. Ghadiri, F. Dessauge, M. Duhamel, M.P. Rebollo, F. Álvarez-Franco y A. Rebollo, Modulating apoptosis as a target for effective therapy, Molecular Immunology, 43, 1065 (2006). [ Links ]
74. I. Petak, J.A. Houghton y L. Kopper, Molecular targeting of cell death signal transduction pathways in cancer, Current Signal Transduction Therapy, 1, 113 (2006). [ Links ]
75. L.C. Huber, O. Distler, R.E. Gay y S. Gay, Antisense strategies in degenerative joint diseases: sense or nonsense? Advanced Drug Delivery Reviews, 58, 285 (2006). [ Links ]

