










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Ciencias Químico - Farmacéuticas
Print version ISSN 0034-7418On-line version ISSN 1909-6356
Rev. colomb. cienc. quim. farm. vol.39 no.2 Bogotá July/Dec. 2010
Artículo de Investigación
Evaluación del efecto in vitro de anticuerpos inducidos por péptido-miméticos derivados del antígeno de superficie de merozoito-2 de Plasmodium en malaria causada por Plasmodium yoelii y Plasmodium berghei en ratones BALB/c
Assessment of the in vitro effect of antibodies induced by peptido-mimetics derived from the Plasmodium merozoite surface protein-2 in malaria caused by P. berghei and P. yoelii in BALB/c mice
Liliana Patricia Lesmes 1, Gina Marcela Gallego 2,3 , Luisa Fernanda Carreño 1,2 , Johan Hoebeke 4 , José Manuel Lozano 1,2,3 .
1 Universidad Nacional de Colombia, Bogotá, D. C., Colombia.
2 Fundación Instituto de Inmunología de Colombia (FIDIC), carrera 50 No. 26-20, Bogotá, D. C., Colombia.
3 Universidad del Rosario, Bogotá, D. C., Colombia.
4 UPR 9021, IBMC, CNRS, Universidad Louis Pasteur, Estrasburgo, Francia.
Correos electrónicos: jmlozanom@unal.edu.co, jm_lozano@fidic.org.co
Recibido: 15 de septiembre de 2010. Aceptado: 6 de noviembre de 2010.
RESUMEN
Con base en estudios realizados previamente en los cuales se identificaron los residuos críticos para la unión de la secuencia 21 KNESKYSNTFINNAYNMSIR 40 del antígeno MSP-2 del Plasmodium falciparum, se diseñaron y sintetizaron dos secuencias de pseudopéptidos amida reducida en las cuales se sustituyó un enlace peptídico normal por su isóstero ψ[CH2 -NH] entre los residuos fenilalanina-isoleucina y entre los residuos isoleucina-asparagina, para dar lugar a los análogos codificados ψ-128 (forma monomérica), ψ-129 (forma polimérica), ψ-130 (forma monomérica) y ψ-131 (forma polimérica). Con los péptido-miméticos obtenidos en forma de polímero se inmunizaron ratones BALB/c para generar anticuerpos monoclonales que presentaron isotipo IgM. Mediante ensayos controlados de inmunización in vitro se indujo el cambio isotipo de los clones reactivos aislados de los hibridomas obtenidos de manera reproducible. Las inmunoglobulinas aisladas se ensayaron por su capacidad funcional neutralizadora de la infección controlada in vitro de cepas de Plasmodium de roedores a glóbulos rojos. Los resultados obtenidos evidencian el papel que pueden tener los anticuerpos inducidos por péptido-miméticos Plasmodium en ensayos de infección realizados en modelos animales de experimentación.
Palabras clave: péptido-mimético, anticuerpo, inmunización in vitro, actividad antiPlasmodium.
ABSTRACT
Based on previous studies in which those residues being critical for Plasmodium falciparum binding to red blood cells (RBCs) through the antigenic sequence ( 21 KNESKYSNTFINNAYNMSIR 40 ) from the merozoite surface antigen-2 (MSP-2) were identified, we have designed and synthesized two reduced amide pseudopeptide sequences based on the 31 IN 32 binding motif. Synthesized peptido- mimetics, possess each one a modified peptide bond that presented as a ψ[CH 2 -NH] reduced amide isoster bond, to allowing the Phe-Ile modified aminoacid pair allowing pseudopeptides coded ψ-128 (monomer form) and ψ-129 (polymer form) and the Ile-Asn modified aminoacid pair for pseudopeptides coded ψ-130 (monomer form) and ψ-131 (polymer form). By using the polymer forms of both peptido-mimetics as immunogens, monoclonal antibodies were produced in BALB/c mice. These Ig showed an IgM isotype. The isotype antibody switching was lead by in vitro immunization of the original hybridomas. Isolated immunoglobulins were tested for their functional in vitro activity, on a infection controlled experiment of rodent Plasmodium strains infecting red blood cells. Obtained results reveal the role played by antibodies to peptido-mimetic in infection assays performed further on animal experimental models.
Key words: peptido-mimetic, antibody, in vitro immunization, antiPlasmodium activity.
INTRODUCCION
La malaria es la enfermedad parasitaria de mayor importancia para la salud mundial, es causada por una de las cinco especies del género Plasmodium: P. falciparum, P. vivax, P. ovale, P. malariae y recientemente se describió el efecto del P. knowlesi como agente causal de infección por malaria en el humano (1). Históricamente, la malaria ha sido responsable de la muerte de más personas que ninguna otra enfermedad parasitaria y aún cuenta con altos índices de morbilidad y mortalidad (2). Se estima que más de 2,7 millones de personas, principalmente niños, mueren de malaria cada año, y una población de más de 2 billones vive en regiones endémicas para esta enfermedad (3). La malaria es el principal problema de salud pública de las regiones tropicales y subtropicales del mundo; pero debido a la resistencia a los insecticidas que viene presentando el vector transmisor (mosquitos del género Anopheles) y la expansión gradual de cepas del parásito resistentes a las drogas antimaláricas, esta infección se ha convertido en un problema global. Además, el fenómeno del calentamiento global afecta la distribución mundial de la malaria debido a la optimización de la transmisión endémica, incremento de la tasa de propagación en las regiones libres de la enfermedad y aumento en la densidad del mosquito vector (4, 5).
Igualmente, la malaria es un problema socioeconómico y de salud pública en regiones donde la enfermedad es endémica, ya que los países que la padecen presentan una baja tasa de crecimiento económico como consecuencia de numerosos factores, incluyendo efectos de la enfermedad sobre la fertilidad, crecimiento de la población, el ahorro y la inversión, la productividad del trabajador, ausentismo, mortalidad prematura y costos médicos (6).
En Colombia, que es uno de los países de América que presenta alta incidencia en malaria, en la última semana epidemiológica del 2010 se notificaron al sistema de vigilancia en salud pública (sivigila) del Instituto Nacional de Salud, un total de 199.107 casos, de los cuales, 1.411 fueron causados por malaria mixta, 32.664 por P. falciparum, 50 por P. malarie, 164.386 por P. vivax, 596 casos de malaria complicada y 23 muertes; las áreas de mayor riesgo incluyen la rivera baja del río Cauca, la costa pacífica, la rivera alta del río Sinú y el Urabá (7).
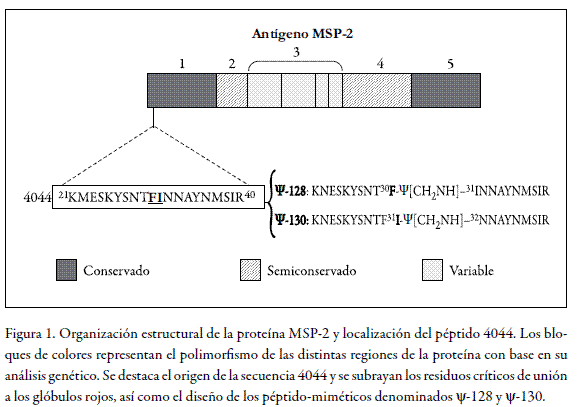
Por estas razones es necesario el desarrollo de una vacuna eficaz y alternativamente el desarrollo de nuevos métodos terapéuticos contra la malaria. En la consecución de estos objetivos, se han estudiado diversas proteínas del Plasmodium falciparum, entre las cuales se encuentra la proteína de superficie del merozoito MSP-2, también conocida como MSA-2 (8). Este antígeno de superficie de aproximadamente 30 kDa (9) a 45 kDa (10) está constituido por dos regiones genéticamente conservadas, una en el extremo carboxi y una en el extremo amino, una región variable y dos regiones semiconservadas en la región central, tal como se observa en la figura 1 (11).
Al realizar el análisis experimental mediante ensayos de unión de los péptidos derivados de la secuencia completa de la proteína MSP-2 a glóbulos rojos, se identificaron péptidos con alta afinidad de unión a los glóbulos rojos (de sus siglas en inglés High activity binding peptides, habp), entre ellos la secuencia de aminoácidos del residuo 21 al 40, denominado 4044 (8). A este péptido se le identificaron los residuos críticos para la unión a glóbulos rojos y, basados en esta información, se sintetizaron dos nuevas secuencias en las cuales se modificaron los enlaces peptídicos con el propósito de evaluar posibles cambios en la estructura con la respuesta biológica frente a las ofrecidas por el péptido nativo 4044. En este trabajo se muestra el empleo de cuatro pseudopéptidos derivados del péptido 4044, a los cuales se les modificó en cada caso un solo enlace de origen natural tipo amida por un enlace modificado químicamente tipo amida reducida, con el objetivo de evaluar su papel frente a una infección por malaria murina in vitro e in vivo. Otro importante componente de este trabajo está constituido por los anticuerpos IgM e IgG de diferentes subclases y su efecto como potencial efecto anti-Plasmodium in vitro. Los péptido-miméticos diseñados de manera sitio-dirigida estimularon anticuerpos de isotipo IgM. Los hibridomas productores de estas inmunoglobulinas se inmunizaron in vitro para así inducir su cambio de isotipo. Estos clones fueron posteriormente caracterizados por su reactividad anti-Plasmodium y su efecto en un ensayo controlado demostró que inmunoglobulinas inducidas por estos péptido-miméticos específicamente inhibieron el desarrollo biológico de este patógeno, por lo que estas propiedades funcionales deberán comprobarse en ensayos in vivo para así poder inferir los posibles mecanismos moleculares involucrados en el efecto inmunoterapéutico de éstos.
MATERIALES Y MÉTODOS
En la modificación de los aminoácido-aldehídos y en la síntesis de los péptidos y pseudopéptidos se usaron L-aminoácidos protegidos en el amino α con el grupo t-Boc provenientes de Bachem (Torrance, CA, EE. UU.); (TBTU) O-(benzotriazol-1)-N,N,N;,N´-tetrafluoroborato de tetrametiluronio, de Bachem (Torrance, CA, EE. UU.); éter etílico (Et2 O); acetato de etilo (EtOAc), n-hexano; trietil amina (TEA); e hidruro de litio y aluminio (Merck, Darmstadt, Alemania); diclorometano (DCM); y N,N´,-dimetilformamida (DMF) (Panreac, Barcelona, España) y O,N-dimetil hidroxilamina-hidrocloruro. Todos los solventes usados fueron grado analítico y el éter usado en la reducción se secó en el laboratorio por destilación con sodio metálico y bezofenona. Los solventes deuterados se obtuvieron de Cambridge Isotope Laboratories (Andover, MA, EE. UU.).
Cromatografía en capa fina
La cromatografía en capa fina (TLC) se usó para monitorear la reacción de formación de la carboxamida, y para comparar por tiempo de retención el aminoácido con la carboxamida y el aminoaldehído; también fue útil para identificar impurezas en la reacción. El sistema de solventes usados para fenilalanina fue acetato de etilo: n-hexano (1:2) y para isoleucina fue acetato de etilo: n-hexano (1:1). El análisis se realizó por fluorescencia a longitud de onda corta Uv (254 nm) o por reacción con ninhidrina.
Infrarrojo - transformada de Fourier (IR-TF)
Los espectros infrarrojos para todos los L-aminoácidos y sus derivados, se tomaron y se registraron en un espectrofotómetro Jasco FT/IR-460 plus (Tokio, Japón); las muestras se analizaron en una celda PIKE MIRacle TM AG ángulo de reflexión horizontal, ATR (Pike Technologies Madison, wi, EE. UU.).
Resonancia magnética nuclear de hidrógeno ( 1 H-RMN)
Los espectros de resonancia magnética nuclear de hidrógeno se obtuvieron y se registraron por disolución de 10 mg de suspensión oleosa o del material liofilizado en dimetilsulfóxido-D6 (D, 99,9%) o cloroformo-D (D, 99,8%), y se realizaron 64 barridos en un espectrofotómetro Bruker DRX 500MHz a 300K (Darmstadt, Alemania).
Síntesis de péptido-miméticos del tipo amida reducida ψ[CH 2 -NH]
La síntesis de pseudopéptidos involucra síntesis orgánicas alternas, como lo es la reducción a aminoaldehído del aminoácido comprometido en el enlace modificado de la secuencia peptídica; la reducción de cada uno de los aminoácidos modificados usados en este trabajo se realizó siguiendo el procedimiento propuesto por Fehrentz y Castro (12).
El procedimiento estándar para la reducción del grupo carbonilo del aminoácido involucra la síntesis de un intermediario (carboxamida), seguido de la reducción con LiAlH4 sobre éter seco. Tal como lo describe la figura 2, esta síntesis tiene un rendimiento global cercano al 80% para la mayoría de los aminoácidos; a pesar de ser una síntesis bastante limpia y la cual presenta un bajo porcentaje de reacciones colaterales, se hace necesaria una caracterización tanto del aminoácido como de la carboxamida y del producto final; se empleó para la caracterización de nuestros compuestos técnicas analíticas como son, infrarrojo, 1 H-RMN, y cromatografía de capa fina (TLC).
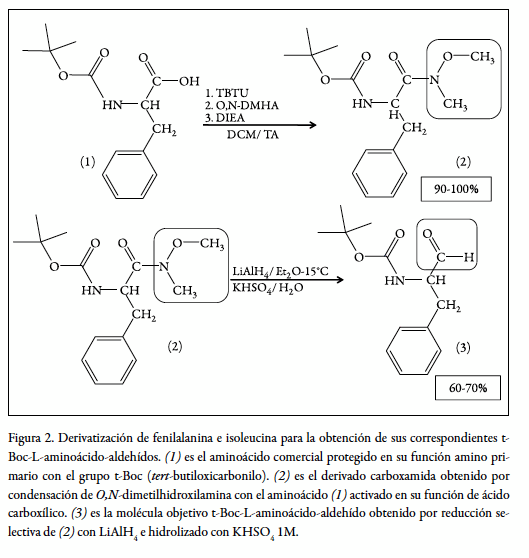
Los resultados obtenidos de la caracterización para los aminoácidos modificados fenilalanina e isoleucina para la síntesis de los pseudopéptidos ψ-128 (F-ψ[CH2 NH]-I), ψ-129 (F-ψ[CH2 NH]-I, en forma polimérica), ψ-130 (I-ψ-[CH2 NH]-N) y ψ-131 (I-ψ-[CH2NH]-N, en forma polimérica) se muestran a continuación.
La síntesis química de los cuatro pseudopéptidos se llevó a cabo empleando aminoácidos protegidos con el grupo t-Boc en el amino alfa, siguiendo el procedimiento de síntesis de péptidos en fase sólida descrito por Merrifield (13). El acople del aminoaldehído a la secuencia requiere unas condiciones de síntesis diferentes a las de un acople normal entre aminoácidos; es necesario mencionar que el tiempo de reacción de este acople es mayor al de un acople normal entre aminoácidos.
La reacción procede al disolver tres equivalentes del aminoaldehído en 2 mL de DMF/ ácido acético al 1% y luego poner en contacto con la resina-péptido manteniendo agitación por un período de tres horas; terminado este tiempo, descartar la solución aminoaldehído/DMF y realizar lavados con DMF, IPA y DCM 3 mL de cada uno por un minuto. Preparar una solución de un equivalente de NaCNBH3 en DMF/ácido acético al 1% y poner a reaccionar con ella la resina-péptido por una hora; finalizado este tiempo, se realizan los mismos lavados del paso anterior y se hace el test de ninhidrina; si el resultado es positivo (solución de color azul) se repite el procedimiento las veces que sea necesario hasta observar que aparentemente no hay grupos amino libres. Como ya se mencionó, esta reacción es un poco lenta, no es de extrañarse que se haga necesario repetir el acople aproximadamente cinco veces hasta obtener una reacción completa; sin embargo, no es recomendado dejar el acople períodos más largos a los sugeridos en este documento. Terminada la reacción de este acople, se desprotege el grupo amino y se continúa la síntesis de la forma tradicional.
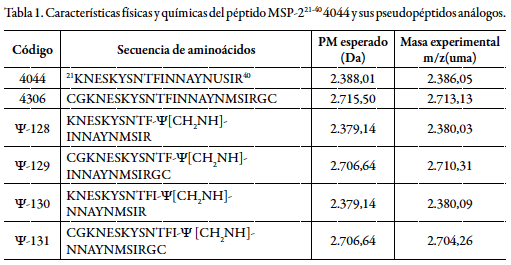
Los pseudopéptidos utilizados para el estudio que se describe en este documento, son análogos a la secuencia del péptido denominado 4044, el cual corresponde -como ya se mencionó- a la región de aminoácidos 21 a 40 de la proteína MSP-2; en la tabla 1 se muestran dichas secuencias, sus respectivos códigos y caracterización.
Evaluación de afinidad relativa por BiaCore (resonancia plasmática de superficie) (11)
Las medidas se realizaron en un equipo Biacore-3000 (Uppsala, Suecia), el cual cuenta con un chip como sensor (CM5); se usaron los siguientes reactivos: EDC, NHS, etanolamina y como buffer de corrida se usó HBS-EP. Los péptidos 4044 y ψ-130 fueron inmovilizados en buffer acetato 10 mM pH 4,9 (100 µg/mL) a un flujo de 5 ïL/min por un período de 7 min; después se realizó la activación de CM5 vía grupos carbono carboxílicos con EDS y vía grupos amino con NHS por el mismo tiempo; luego de inmovilizados los péptidos, los canales se neutralizaron con etanolamina 1 M, pH 8, por 4 min con el mismo flujo. Los canales control son producidos por activación-desactivación en ausencia de péptido.
Los estudios de inhibición se realizaron manteniendo un flujo de 20 µL/min, en donde hubo una fase de asociación por 90 segundos, seguido de una fase de disociación por 60 segundos y una fase de regeneración por 30 segundos con 50 mM de HCl.
La concentración de anticuerpo monoclonal usada para la inmovilización del péptido 4044 fue de 5 µg/mL y 50 µg/mL en la inmovilización de r130; las soluciones de IgM contenían concentraciones de péptido entre 42 y 0,164 µM y sin péptido para el control; los análisis se realizaron con ayuda del software sigma-plot.
Obtención de anticuerpos monoclonales anti-ψ-128 (129) y ψ-130 (131)
Las células del bazo de ratones BALB/c previamente inmunizados con los pseudopéptidos ψ-129 y ψ-131, se fusionaron físicamente con células de la línea celular de mieloma murino X63Ag8, usando polietilenglicol 3000-3700 (Sigma Chemical Co., St. Louis, mo, Estados Unidos) de acuerdo con la metodología descrita anteriormente (11, 14), para obtener así hibridomas reactivos. Estos hibridomas se clonaron dos veces por dilución limitante. Después de una evaluación específica por ELISA y Western blot, los clones reactivos se expandieron en un volumen de cultivo mayor para la producción de estos anticuerpos a una escala mayor.
Cambio de isotipo por inmunización in vitro de hibridomas
Las células reactivas de los clones derivados de los pseudopéptidos ψ-129 y ψ-131 se distribuyeron de 100.000 células/pozo en una caja de 96 pozos de fondo plano, en medio de cultivo RPMI suplementado con suero fetal bovino (SFB) al 15%. Estas células se pulsaron cinco veces con 200 ng de su respetivo pseudopéptido inductor monómero (ψ-128 y ψ-130, respectivamente), diluido en RPMI 15% SFB a intervalos de ocho días por pulso. El cambio de isotipo se verificó a través del kit monoclonal antibody isotyping (HRP/ABTS) de Thermo Scientifi c (Rockford, il, EE. UU.). Los sobrenadantes de los hibridomas productores de anticuerpos se recogieron periódicamente. Los anticuerpos presentes en los sobrenadantes provenientes del clon inducido por el ψ-128 (ψ-129) se codificaron como SN 6 y SN 7, y los anticuerpos de los sobrenadantes provenientes del clon inducido por el ψ-130 (ψ-131) se codificaron como SN: G, M, O y Q. Seguidamente se evaluó la reactividad de los anticuerpos presentes en cada uno de los sobrenadantes de los clones frente a lisado de P. falciparum cepa FCB2 por Western Blot.
Producción in vivo de anticuerpos monoclonales anti-ψ-129 y ψ-131
Con el objetivo de producir anticuerpos específicos en mayor cantidad, se empleó la estrategia de producción in vivo por inoculación vía intraperitoneal (IP) de los clones en ratones BALB/c para generar fluidos ascíticos. Para obtener este fluido ascítico enriquecido en anticuerpos monoclonales, la cavidad peritoneal de los ratones se irritó por inyección vía ip con 500 µL de pristano (Sigma Chemical Co., Saint Louis, mo, EE. UU.). Ocho días después, a cada animal se le administró por la misma vía, 500 µL de los clones celulares derivados de ψ-129 y ψ-131 (1.000.000 células) suspendidos en medio RPMI incompleto. Luego de ochos días, se recogió el fluido ascítico por punción abdominal; este fluido se centrifugó a 800 rpm durante 5 min, se inactivó y deslipidificó (11) para continuar con la purificación de los anticuerpos.
Purificación de inmunoglobulinas (Igs)
Los sobrenadantes provenientes de cultivo celular o de los fluidos ascíticos se sometieron a tratamiento con una solución saturada de sulfato de amonio al 80% para sedimentar la proteína presente en ellos. Posteriormente al proceso de centrifugación, los sobrenadantes se separaron cuidadosamente y el precipitado se reconstituyó en buffer Tris-HCl 10 mM pH 8,0; luego se dializó contra este mismo buffer empleando una membrana Spectra-pore® dialysis de 3500 MWCO (Houston, Texas, EE. UU.) a 4 °C. Después, los precipitados enriquecidos con anticuerpos y dializados se sometieron a cromatografía de intercambio aniónico débil usando DEAE-Sephadex A-25 resina (Pharmacia, Uppsala, Suecia) previamente equilibrada con buffer Tris-HCl 10 mM, pH 8,0 empacada en una columna de vidrio de 50 mL. Sobre esta columna se sirvió la solución enriquecida en anticuerpo para permitir así su interacción con la resina. Posteriormente, las inmunoglobulinas se eluyeron de la columna por incremento progresivo en la fuerza iónica del buffer desde 50 mM, 100 mM y 500 mM de NaCl. Un total de cuatro volúmenes de columna (250 mL) se adicionó por paso, colectando fracciones de 10 mL a un flujo constante de 0,5 mL/min. La resina se reequilibró con buffer 10 mM Tris-HCl, pH 8,0. Todas las fracciones se caracterizaron por ensayos de reactividad Dot Blot directo para detección de inmunoglobulinas. Las fracciones reactivas se mezclaron en grupos o pooles con base en tres criterios: su semejante perfil electroforético, su reactividad específica en ensayos de Dot Blot y en su contenido comparable en proteína total de acuerdo con el ensayo colorimétrico del ácido bicinconínico (BCA). Estos grupos de fracciones se dializaron contra PBS pH 7,2 a 4 °C. La pureza de cada pool de fracciones se determinó por electroforesis en geles de poliacrilamida dodecil sulfato de sodio (SDS-PAGE) y Western blot (15).
Estandarización de modelos de infección con malaria por P. yoelii y P. berghei en ratones BALB/c
Con el objetivo de establecer infección de malaria in vivo en ratones BALB/c (obteni- dos del bioterio de la Facultad de Medicina veterinaria de la Universidad Nacional de Colombia), se descongelaron y activaron cepas de P. yoelii 17XL y P. berghei ANKA crio-preservadas en solución de Krebs (NaCl 0,85%, glucosa 5%, sorbitol 4,2% p/v). Para ello se realizaron tres lavados con RPMI incompleto a 2.500 rpm durante 10 min y finalmente se administraron por vía intraperitoneal a tres ratones hembras de la cepa BALB/c de cinco semanas de edad, una cantidad definida de parásito resuspendido en 500 µL de RPMI incompleto. A partir del sexto día de la posinfección, se evaluó la parasitemia diariamente con tinción de Wright en extendidos de sangre periférica. Una vez comprobados los niveles de parasitemia entre el 20% y 50%, se realizó un pase in vivo por administración ip de 1 x 10 5 glóbulos rojos parasitados para mantener así la viabilidad de la cepa del Plasmodium en estudio antes de ser empleada para los ensayos de infección controlada in vitro.
Para realizar el cultivo in vitro de estas cepas de malaria murina, se permitió que los ratones infectados alcanzaran niveles de parasitemia del 10%. Para la identificación del parásito, se realizó el recuento de glóbulos rojos parasitados en una cámara de Neubauer con tinción de naranja de acridina.
Actividad in vitro anti-Plasmodium de inmunoglobulinas inducidas por péptidomiméticos
Dada la importancia de tener modelos de infección in vitro de cepas de Plasmodium tanto humanas como de otras especies (roedores), el poder mantener en cultivo por períodos superiores a los diez días este tipo de patógenos parasitarios, motivó este trabajo el abordaje de esta problemática y, en consecuencia, se logró establecer eficientemente dos cultivos de cepas de Plasmodium berghei y Plasmodium yoelii.
Para ello, se efectuó una adaptación de condiciones estándar para el crecimiento in vitro P. yoelii y P. berghei (16-19). Por tanto, se tomaron 500 µL de sangre de ratón infectado con parasitemia entre 7-10% en PBS - 0,1% citrato de sodio. Luego se centrifugó a 800 rpm durante 10 min. Se hicieron dos lavados con RPMI incompleto 0,1% citrato de sodio. Asépticamente, se adicionó al sedimento 2,5 mL de medio RPMI completo (20% de SFB, gentamicina 5 mg/mL, antibiótico antimicótico 1%). En cajas de cultivo celular de 24 pozos se adicionó a cada pozo 400 µL de medio completo, 500 µL de anticuerpos o controles y, finalmente, 100 µL de glóbulos rojos parasitados o glóbulos rojos sanos, para un volumen final de 1.000 µL/pozo. Se emplearon como controles de ensayo glóbulos rojos sanos, glóbulos rojos parasitados, cloroquina 2,83 x 10 -3 mM y EGTA 55,55 mM. Cada muestra se evaluó por duplicado. Este ensayo se incubó por 15 horas a 37 °C en cámara de gases suplementada con 90% N2, 5% CO2, 5% O2 . En los días 1, 4, 7, 10 y 14, se preparó una mezcla de 500 µL de GR sanos y 7 mL de medio completo con el fin de adicionar 300 µL de esta mezcla a cada uno de los pozos. Se evaluaron los porcentajes de parasitemia a diario a partir del segundo día por recuento en cámara de Neubauer con naranja de acridina, en una dilución 1/500. Por no existir evidencia previa a este trabajo, en este tipo de ensayos no se emplea otro tipo de controles, como los anticuerpos monoclonales inducidos por otros antígenos con capacidad inhibitoria de la infección.
RESULTADOS Y DISCUSIÓN
Diseño, síntesis y caracterización estructural del péptido de alta unión 4044 del antígeno MSP-2 y de sus péptido-miméticos de tipo amida reducida
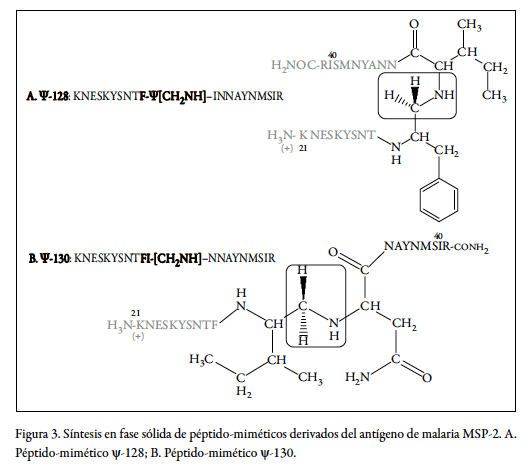
Como se observa en la figura 1, el péptido codificado como 4044 se ubica en el bloque 1 conservado genéticamente de la región amino terminal de la secuencia del antígeno MSP-2. El motivo de unión a glóbulos rojos definido por el par de residuos 30 I-N 31 sirvió como base para el diseño para la sustitución sitio-dirigida de un solo enlace peptídico a la vez por el isóstero ψ[CH2-NH], para dar lugar a los pseudopéptidos ψ-128 (F-I) y ψ-130 (I-N) y a sus correspondientes formas poliméricas ψ-129 y ψ-131, respectivamente.
En la figura 2 se esquematiza el proceso sintético de transformación del aminoácido comercial fenilalanina Boc-L-Phe-OH hasta su forma aminoácido aldehído Boc-L-Phe-H. Esta estrategia de dos etapas incluye la derivatización de compuesto (1) a su forma carboxamida (2) para proteger al carbono carbonílico del aminoácido el cual luego se trata con un agente reductor fuerte, el hidruro de litio y aluminio para dar lugar a la formación de un anillo parcial de cinco miembros que incluye al litio, el cual es posteriormente hidrolizado para generar la forma aldehído del aminoácido (3). Este proceso tiene un rendimiento global del 60-70% cuando se realiza con cualquier aminoácido, exceptuando los aminoácidos asparagina y glutamina (Asn y Gln), los cuales deben tratarse de manera diferente.
Caracterización espectroscópica de aminoácido-aldehídos
Derivados de Boc-L-Phe
Boc-L-Phe-OH, Rf = 0,40 acetato de etilo: n-hexano (1:2).
1H-NMR [dimetilsulfóxido-D6 (D, 99,9%)]: δ = 1,42 ppm (9H, C(CH3)); δ = 2,66 ppm (2H, Hβ); δ = 5,76 ppm (1H, Hα); δ = 7,17 ppm (2H, H2-6-anillo); δ = 7,22 ppm (2H, H3-5-anillo); δ = 7,23 ppm (2H, H4-anillo); δ = 7,28 ppm (1H, NHα); δ = 2,5 ppm (m, 6H, dimetilsulfóxido).
FT-IR (sólido amorfo): ν(cm -1 ) = 3.299,14, 3.246,12, 2.974,23, 1.707,43, 1.644,67, 1.496,54, 1.405,60, 1.359,34, 1.272,23, 1.156,45, 774.09.
Boc-L-Phe-N(CH 3 )OCH 3, Rf = 0,50 acetato de etilo: n-hexano (1:2).
1H-NMR [dimetilsulfóxido-D6 (D, 99,9%)]: δ = 1,39 ppm (9H, C(CH3)); δ = 2,81 ppm (2H, Hβ); δ = 3,18 ppm (3H, N-(CH3)); δ = 3,65 ppm (3H, N-O-(CH3)); δ = 5,25 ppm (1H, Hα); δ = 7,19 ppm (2H, H2-6-anillo); δ = 7,24 ppm (2H, H3-5- anillo); δ = 7,25 ppm (2H, H4-anillo); δ = 7,28 ppm (1H, NHα); δ = 2,5 ppm (m, 6H, dimetilsulfóxido).
FT-IR (película oleosa): ν(cm -1 ) = 3.299,14, 3.246,12, 2.974,23, 1.894,98, 1.657,14, 1.644,67, 1.496,54, 1.405,60, 1.359,34, 1.272,23, 1.156,45, 774,09.
Boc-L-Phe-H, Rf = 0,69 acetato de etilo: n-hexano (1:2).
H-NMR [dimetilsulfóxido-D6 (D, 99,9%)]: δ = 1,48 ppm (9H, C(CH3)); δ = 2,81 ppm (2H, Hβ); δ = 5,01 ppm (1H, Hα); δ = 7,17 ppm (2H, H2-6-anillo); δ = 7,23 ppm (2H, H3-5-anillo); δ = 7,25 ppm (2H, H4-anillo); δ = 7,32 ppm (1H, NHα); δ = 9,63 ppm (1H, formilo); δ = 2,5 ppm (m, 6H, dimetilsulfóxido).
FT-IR (película oleosa): ν(cm -1 ) = 3.360,43, 3.246,12, 2.974,23, 1.677,05, 1.644,67, 1.496,54, 1.405,60, 1.359,34, 1.272,23, 1.156,45, 774,09.
Derivados de Boc-L-Ile
Boc-L-Ile-OH, Rf = 0,66 acetato de etilo: n-hexano (1:1).
1H-NMR [dimetilsulfóxido-D6 (D, 99,9%)]: δ = 0,94 ppm (3H, Hγ); δ = 0,97 ppm (3H, Hδ); δ = 1,20 ppm (2H, Hγ); δ = 1,45 ppm (9H, C(CH3)); δ = 1,92 ppm (1H, Hβ); δ = 4,29 ppm (1H, Hα); δ = 7,25 ppm (1H, NHα); δ = 2,5 ppm (m, 6H, dime- tilsulfóxido).
FT-IR (sólido amorfo): ν(cm -1 ): 3.352,65, 3.299,24, 2.963,19, 1.726,67, 1.699,54, 1.673,08, 1.530,43, 1.451,60, 1.412,79, 1.299,21, 1.160,34, 1.016,01, 776,44.
Boc-L-Ile-N(CH 3 )OCH 3, Rf = 0,54 acetato de etilo: n-hexano (1:1).
1H-NMR [dimetilsulfóxido-D6 (D, 99,9%)]: δ = 0,89 ppm (3H, Hγ); δ = 0,93 ppm (3H, Hδ); δ = 1,12 ppm (2H, Hγ); δ = 1,43 ppm (9H, C(CH3)); δ = 2,80 ppm (1H, Hβ); δ = 3,21 ppm (3H, N-(CH3)); δ = 3,78 ppm (3H, N-O-(CH3)); δ = 4,61 ppm (1H, Hα); δ = 7,25 ppm (1H, NHα); δ = 2,5 ppm (m, 6H, dimetilsulfóxido).
FT-IR (polvo): ν(cm -1 ): 3.352,65, 3.299,24, 2.963,19, 1.699,54, 1.688,14, 1.654,77 1.530,43, 1.451,60, 1.412,79, 1.299,21, 1.160,34, 1.016,01, 776,44.
Boc-L-Ile-H, Rf = 0,58 acetato de etilo: n-hexano (1:1).
1H-NMR [dimetilsulfóxido-D6 (D, 99,9%)]: δ = 0,91 ppm (3H, Hγ); δ = 0,94 ppm (3H, Hδ); δ = 0,96 ppm (2H, Hγ); δ = 1,45 ppm (9H, C(CH3)); δ = 2,03 ppm (1H, Hβ); δ = 4,37 ppm (1H, Hα); δ = 7,25 ppm (1H, NHα); δ = 9,66 ppm (1H, formilo); δ = 2,5 ppm (m, 6H, dimetilsulfóxido).
FT-IR (película oleosa): ν(cm -1 ): 2.963,21, 1.693,09, 1.530,43, 1.451,60, 1.412,79, 1.299,21, 1.160,34, 1.016,01, 776,44.
En un proceso sintético realizado en fase sólida, el sintón (3) se incorpora a la cadena peptídica creciente in situ, para luego someterse a una segunda reducción en condiciones más suaves con cianoborohidruro de sodio (NaCNBH3), para dar así lugar a los péptido-miméticos objetivo, tal como se ilustra en la figura 3. En A se esquematiza la secuencia del péptido-mimético ψ-128 y en B la del ψ-129 obtenidas mediante los procedimientos en fase sólida descritos. En la tabla 1 se pueden observar las características fisicoquímicas de las moléculas producidas para este estudio; es decir, el péptido nativo 4044 y sus análogos pseudopeptídicos en sus formas de monómero y polímero.
Todas ellas reunieron las cualidades moleculares físicas y bioquímicas adecuadas para la realización de los experimentos inmunológicos.
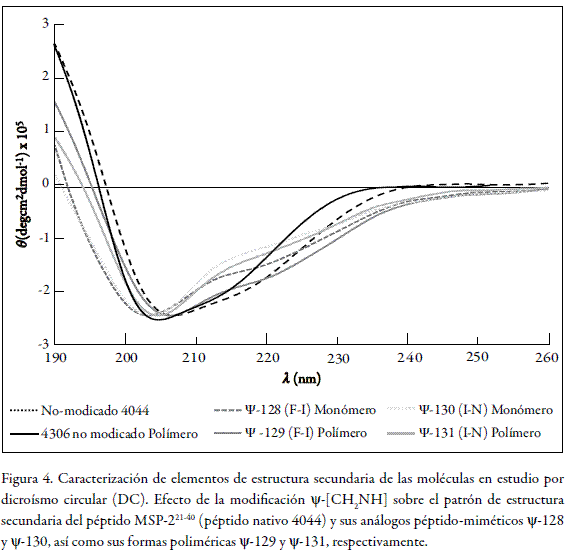
Seguidamente, este nuevo grupo de moléculas se caracterizó en cuanto a la presencia de elementos de estructura secundaria, mediante la técnica de dicroísmo circular (DC). El resultado de este análisis se observa en la figura 4, en la cual se muestran los perfiles obtenidos tanto para el péptido nativo no modificado 4044, como para sus análogos pseudopeptídicos ψ-128 y ψ-130 junto con sus correspondientes formas poliméricas ψ-129 y ψ-131, respectivamente. Como se puede observar, el péptido nativo 4044 y sus análogos presentan algunos rasgos estructurales propios de las hélices α. Esto se evidencia por la presencia de dos valores mínimos de elipticidad molar en 208 nm y 222 nm, respectivamente; pero a la vez presentan rasgos de elementos estructurales más extendidos tipo hebra-β, evidenciando la presencia de un solo mínimo en 218 nm, además de regiones carentes de estructura. Por tanto, la preferencia estructural de estas moléculas es intermedia entre una hélice distorsionada y una hebra beta no estabilizada.
Producción de anticuerpos monoclonales contra los péptido-miméticos
Seguidamente se procedió a la producción de anticuerpos monoclonales inducidos por los péptido-miméticos ψ-128 y ψ-130. Se generó un grupo de hibridomas que en principio presentaron el isotipo pentavalente IgM. Estos hibridomas productores de inmunoglobulinas IgM sirvieron de base para realizar experimentos de inmunización in vitro para inducir el cambio de isotipos a formas de anticuerpos más simples pero con propiedades funcionales comparables o superiores a la inicial. Estas células inducidas se clonaron por dilución limitante.
En la tabla 2 se muestran los principales resultados obtenidos mediante este procedimiento y su versatilidad al inducir el cambio drástico de isotipos de IgM a IgG1, IgG2a, IgG2b e IgG3, cuya actividad neutralizante de la infección por malaria fue evidente.
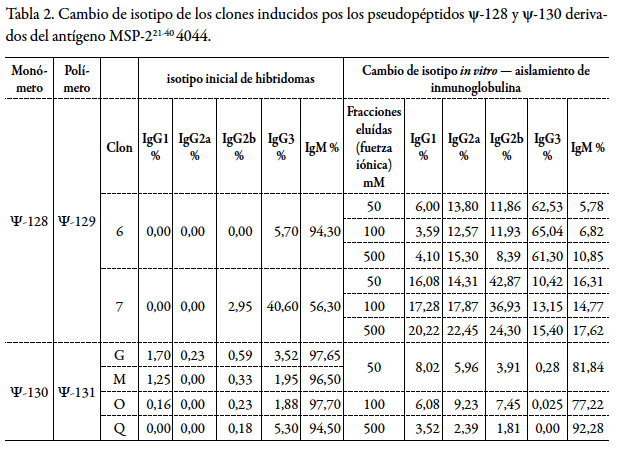
Por otra parte, la versatilidad de este procedimiento de inducción del cambio del isotipo IgM a otros isotipos como IgG1, IgG2a, IgG2b e IgG3, permitió obtener clones celulares definidos hacia formas reactivas de inmunoglobulinas con propiedades neutralizadoras de la infección controlada de cepas murinas de malaria en los ensayos en cultivo celular. Como se observa en la tabla 2, los porcentajes de conversión fueron superiores al 60% para el clon inducido por el pseudopéptido ψ-128 (ψ-129) e inferiores al 10% para los clones generados por el pseudopéptido ψ-130 (ψ-131). Aunque en el segundo de los casos los porcentajes de conversión fueron relativamente bajos, inferiores al 10%, es evidente que el procedimiento de inducción de cambio es eficiente y potencialmente aplicable y susceptible de ser escalonado a niveles biotecnológicos.
Establecimiento de modelos de infección por malaria in vitro
La morfología propia de eritrocitos de ratones BALB/c infectados con las cepas de malaria de roedores Plasmodium yoelii 17XL y Plasmodium berghei ANKA, permitió controlar y caracterizar el ciclo eritrocítico de estas dos entidades por la presencia de los estadios sanguíneos del parásito, como la morfología de anillo, el trofozoito y el esquizonte maduro del parasito de la malaria de roedores. En condiciones controladas se logró mantener cada cultivo celular en etapas temporales hasta de un mes en ambos casos. Para realizar el ensayo de infección in vitro, se tomaron muestras de sangre de un ratón efectivamente infectado con la cepa del Plasmodium correspondiente y se establecieron ensayos de infección por duplicado tanto para los grupos control como para las muestras en estudio.
Una vez se logró establecer los modelos de infección en cultivo controlado in vitro y en ratones hembras de la cepa BALB/c in vivo, se procedió a realizar la evaluación de la actividad funcional de anticuerpos monoclonales inducidos por los péptido-miméticos derivados de la secuencia del péptido nativo 4044.
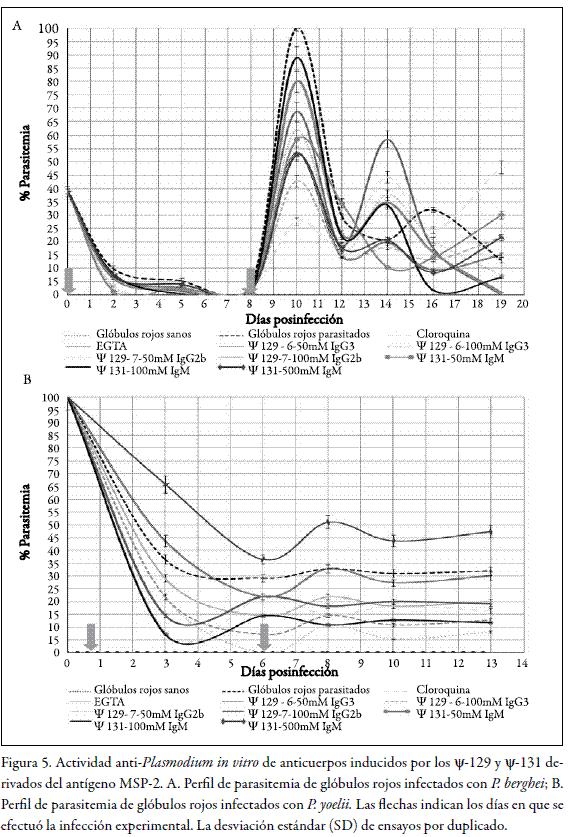
Los estudios para evaluar la actividad antiplasmodio in vitro realizados en cultivo controlado, demostraron un efecto neutralizador de la infección por malaria en ensayos por duplicado. En todos los casos, los porcentajes de parasitemia fueron inferiores a los manejados por el experimento control de infección normal en los glóbulos rojos parasitados con la cepa de P. berghei, tal como se observa en la figura 5A, luego de dos desafíos experimentales. El pico de parasitemia observado hacia el día 10, se debe al efecto del segundo desafío in vitro (véase figura 5).
Dadas las evidencias funcionales de las inmunoglobulinas inducidas por los péptido-miméticos, se decidió realizar estudios fisicoquímicos para evaluar la afinida d del anticuerpo original IgM inducido por el pseudopéptido ψ-128 (ψ-129), gracias a la colaboración del laboratorio de inmunoquímica de péptidos de la Universidad Louis Pasteur en Estrasburgo, Francia.
Afinidad del anticuerpo monoclonal primario IgM inducido por el péptido-mimético ψ-130
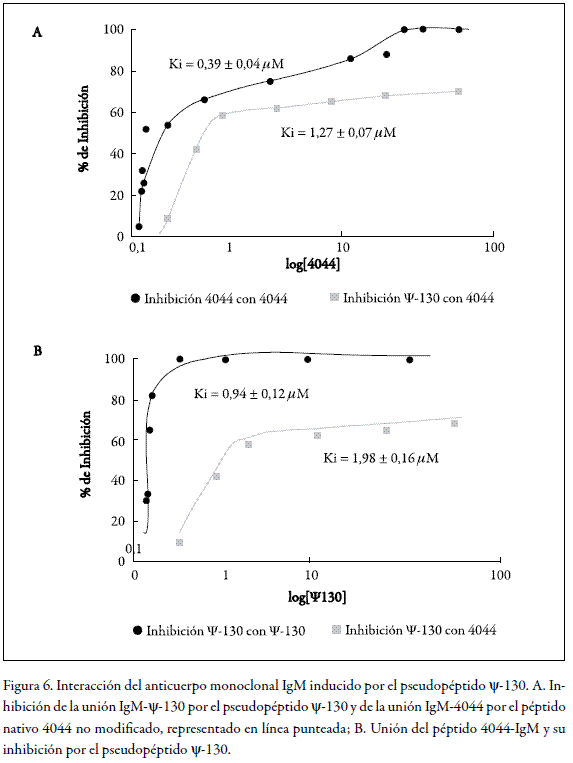
Debido a la multivalencia del anticuerpo monoclonal de isotipo IgM, se decidió estudiar la afinidad de éste, tanto por el péptido nativo 4044 como por uno de su péptido-mimético inductor ψ-130, mediante un ensayo de inhibición de la unión del anticuerpo por cada uno de ellos. Así, tanto el péptido como el péptido-mimético se inmovilizaron sobre las placas de los chips del equipo BiaCore a una densidad alta de 10,3 ng/mm 2 para el péptido 4044 y 3,1 ng/mm 2 para el péptido-mimético ψ-130. En estas condiciones, la velocidad de asociación se define básicamente por transferencia de masa y, en consecuencia, por la cantidad de sitios libres del analito que quedan en la solución. Esto permitió una estimación precisa de la constante de inhibición Ki del péptido. Como se muestra en la figura 6, el anticuerpo monoclonal IgM tuvo aproximadamente la misma afinidad tanto para el péptido nativo 4044 (0,39 ± 0,04 µM) como para el pseudopéptido ψ-130 (0,94 ± 0,12 µM). Las constantes de inhibición heterólogas fueron 1,98 ± 0,16 µM para el pseudopéptido y 1,27 ± 0,04 µM para el péptido natural. Es importante señalar que la afinidad de la IgM parece ser menor por su inductor ψ-130 ( 31 I-N 32 ) que por el péptido nativo no modificado 4044, lo cual demostró que este anticuerpo reconoce y se adapta con facilidad a la estructura 3D del péptido original.
CONCLUSIONES
Las evidencias obtenidas en este estudio acerca de la funcionalidad in vitro de anti- cuerpos inducidos por pseudopéptidos diseñados de manera sitio-dirigida, permiten proponer a estos péptido-miméticos del tipo amida reducida como potenciales componentes candidatos a una formulación de una vacuna de subunidades contra la malaria. Este tipo de acercamiento experimental, tomando como base éste y otros antígenos de superficie del Plasmodium, permitirá evaluar más extensamente los datos reportados en este trabajo así como también sus propiedades in vivo. En consecuencia, la aplicación e implementación de modelos de infección in vitro de malaria de especies como roedores, aves y primates no humanos, constituye una valiosa herramienta para el estudio sistemático de nuevos medicamentos y candidatos a vacuna contra esta enfermedad.
AGRADECIMIENTOS
Los autores de este trabajo agradecemos al posgrado Interfacultades de Microbiología y a la maestría en Ciencias Farmacéuticas, Departamento de Farmacia de la Universidad Nacional de Colombia. A los diferentes grupos funcionales de investigación de la Fundación Instituto de Inmunología de Colombia (FIDIC). A la doctora Lilian Spencer-valero del Departamento de Biología Celular de la Universidad Simón Bolívar (Caracas, venezuela) por la donación de la cepa de Plasmodium yoelii 17XL, y a la doctora Silvia Blair del Laboratorio de Malaria, Facultad de Medicina de la Universidad de Antioquia (Medellín, Colombia) por la donación de la cepa de Plasmodium berghei ANKA. Especial agradecimiento al profesor Manuel Elkin Patarroyo, inspirador de las ideas desarrolladas en este trabajo.
BIBLIOGRAFÍA
1. B. Singh, L.K. Sung, A. Matusop, A. Radhakrishnan, S.G. Shamsul, J. Cox- Singh, A. Thomas, D.J. Conway, A large focus of naturally acquired Plasmodium knowlesi infections in human beings, Lancet, 363(9414), 1017-1024 (2004). [ Links ]
2. R. Tuteja, Malaria the global disease, FEBS Journal, 274(18), 4669 (2007). [ Links ]
3. S.I. Hay, C.A. Guerra, A.J. Tatem, A.M. Noor, R.W. Snow, The global distribution and population at risk of malaria: Past, present, and future, Lancet Infect. Dis., 4, 328 (2004). [ Links ]
4. P.E. Parham, E. Michael, Modeling the effects of weather and climate change on malaria transmission, Environ. Health Persp., 118(5), 620-626 (2010). [ Links ]
5. K.P. Paaijmans, S.S. Imbahale, M.B. Thomas, W. Takken, Relevant microclimate for determining thedevelopment rate of malaria mosquitoes and possible implications of climate change, Malaria J., 9(196), 1-8 (2010). [ Links ]
6. D. D´Alessandro, U. Olaleve, W. Mcguire, P. Langerock, S. Bennett, M.K. Aikins, Mortality and morbidity from malaria in Gambian children after introduction of an impregnated bednet programme, Lancet, 345(8948), 479 (1995). [ Links ]
7. Instituto Nacional de Salud - sivigila, Reporte semana 52 del 2010 [en línea]. Bogotá: sivigila, consultado en noviembre de 2010. url: http://www.ins.gov.co [ Links ]
8. M. Ocampo, M. Urquiza, F. Guzmán, L.E. Rodríguez, J. Suárez, H. Curtidor et al., Two MSA-2 peptides that bind to human red blood cells are relevant to Plasmodium falciparum merozoite invasion, J. Peptide Res., 55(3), 216-223 (2000). [ Links ]
9. C.G. Adda, v.J. Murphy, M. Sunde, L.J. Waddington, J. Schloegel, G.H. Talbo, K. vingas, v. Kienzle, R. Masciantonio, G.J. Howlett, A.N. Hodder, M. Foley, R.F. Anders, Plasmodium falciparum merozoite surface protein 2 is unstructured and forms amyloid-like fibrils, Mol. Biochem. Parasit., 166(2), 159-171 (2009). [ Links ]
10. J.A. Smythe, M.G. Peterson, R.L. Coppel, A.J. Saul, D.J. Kemp, R.F. Anders, Structural diversity in the 45-kilodalton merozoite surface antigen of Plasmodium falciparum, Mol. Biochem. Parasitol., 39(2), 227-234 (1990). [ Links ]
11. J.M. Lozano, F.J. Montoya, J. Hoebeke, G.H. Cifuentes, M. Forero, M.E. Patarroyo, Antibodies induced by Plasmodium falciparum merozoite surface antigen- 2-designed pseudopeptides possess neutralizing properties of the in vitro malarial infection, Peptides, 28(10), 1954-1965 (2007). [ Links ]
12. J.A. Fherentz, B. Castro, An efficien t synthesi s o f opticall y active α-(t-butoxycarbonylamino)-aldehydes from α-amino acids, Synthesis, 8, 676-679 (1983). [ Links ]
13. R.B. Merrifield, Solid phase peptide synthesis. I. The synthesis of a tetrapeptide, J. Am. Chem. Soc., 85, 2149 (1963). [ Links ]
14. G. Kohler, C. Milstein, Derivation of specific antibody producing tissue culture and tumor lines by cell fusion, Eur. J. Immunol., 6(7), 511-519 (1976). [ Links ]
15. J.M. Lozano, L.P. Lesmes, G.M. Gallego, M.E. Patarroyo, Protection against malaria is conferred by passive transferring rabbit F(ab)2´ antibody fragments, induced by Plasmodium falciparum MSP-1 site-directed designed pseudopeptide-BSA conjugates assessed in a rodent model, Mol. Immunol., aceptado para publicación. [ Links ]
16. M.E. Smalley, G.A. Butcher, The in vitro culture of the blood stages of Plasmodium berghei, Int. J. Parasitol., 5, 131-132 (1975). [ Links ]
17. B. Mons, C.J. Janse, J.J.A.B. Croon, H.J. van der Kayy, In vitro culture of Plasmodium berghei using a new suspension system, Int. J. Parasitol., 13(2), 213-217 (1983). [ Links ]
18. P.H. Lewis-Hughes, M.J. Howell, In vitro culture of Plasmodium yoelii blood stages, Int. J. Parasitol., 14(5), 447-451 (1984). [ Links ]
19. M. Ramaiya, Kamath, R. vijayalaxmi, D.M. Renapurkar, Long-term in vitro cultivation of Plasmodium berghei, Int. J. Parasitol., 17(7), 1329-1331 (1987). [ Links ]

