










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Ciencias Químico - Farmacéuticas
Print version ISSN 0034-7418
Rev. colomb. cienc. quim. farm. vol.42 no.1 Bogotá Jan./June 2013
Artículo de investigación científica
Actividad antibacterial de extractos de hormigas de los géneros Crematogaster y Solenopsis
Antibacterial activity extracts of the ant genera Crematogaster and Solenopsis
Germán Matiz Melo1*, María del Rosario Osorio Fortich1
1 Grupo de Investigación en Tecnología Farmacéutica, Cosmética y de Alimentos, GITFCA, Facultad de Ciencias Farmacéuticas, Universidad de Cartagena, Cartagena, Colombia.
*E-mail: gmatizm@unicartagena.edu.co.
Recibido para evaluación: 20 de febrero de 2013.
Aceptado para publicación: 16 de abril de 2013.
RESUMEN
Uno de los principales problemas que enfrenta la comunidad científica en la actualidad, es la aparición cada vez más frecuente de cepas bacterianas patógenas resistentes a los antibióticos y antimicrobianos actuales, porque dichos gérmenes tienen la capacidad, bajo determinadas circunstancias, de hacerse inmunes a tales agentes químicos; por tanto, es necesaria la búsqueda de nuevos agentes antibacterianos. Para este propósito, una buena fuente puede provenir de los insectos sociales, los cuales han desarrollado como estrategia de supervivencia la producción de agentes bactericidas para proteger sus colonias. Este trabajo se centró en determinar si ciertas hormigas que se adaptaron a los ámbitos urbanos, tienen esta actividad. Para ello, se capturaron hormigas de los géneros Crematogaster sp. y Solenopsis sp., que se sometieron a procesos de extracción por maceración, obteniéndose extractos etanólicos totales, muy poco hidrosolubles, los cuales se evaluaron a diferentes concentraciones sobre cepas de bacterias Gram negativas (Klebsiella pneumoniae y Pseudomona aeruginosa) y Gram positivas (Bacillus subtilis spizizenii y Staphylococcus aureus), encontrando actividad inhibitoria al crecimiento en diferentes grados sobre todas ellas, incluyendo una S. aureus resistente a los antibióticos Amoxicilina/Ac. Clavulánico, Ampicilina/Sulbactam y Ceftriazona/Oxacilina; por tanto, estas especies se pueden clasificar como promisorias en la búsqueda de nuevos agentes antimicrobianos.
Palabras clave: Crematogaster, Solenopsis, hormigas, bactericida.
SUMMARY
One of the main problems faced today by the scientific community is the increasingly frequent occurrence of pathogenic bacterial strains resistant to current antibiotics and antimicrobial, because these germs are able, under certain circumstances, to be immune to such chemicals, it is therefore necessary to pursue the search for new agents. For this purpose, a good source can come from social insects, which have developed as a survival strategy, the production of bactericidal agents to protect their colonies. The purpose of this work was to determine if certain ants that have adapted to urban environments, have antibacterial activity and to what extent. For this purpose, ants of the genus Crematogaster sp. and sp. Solenopsis were captured and subjected to extraction process by maceration, yielding total ethanolic extracts, very little soluble, which were evaluated at different concentrations on Gram negative bacteria strains (Klebsiella pneumoniae and Pseudomona aeruginosa) and Gram positive (Bacillus subtilis spizizenii and Staphylococcus aureus), finding growth inhibitory activity at different degrees over all strains, including an S. aureus one resistant to antibiotics Amoxicillin/Clavulanate, Ampicillin/Sulbactam, Ceftriaxone and Oxacillin; therefore, these species can be classified as promising in the search for new antimicrobial agents.
Key words: Crematogaster, Solenopsis, bactericidal activity, ants.
INTRODUCCIÓN
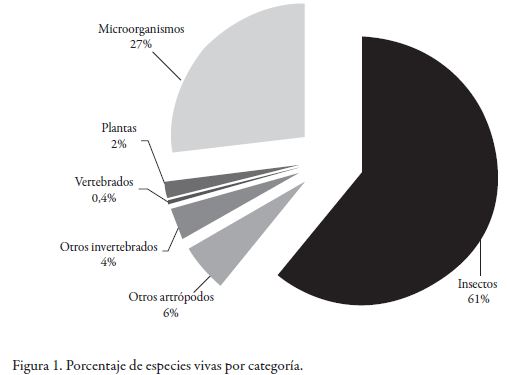
El uso de productos naturales con propiedades terapéuticas, es tan antiguo como el hombre mismo, quien por observación y experimentación empírica (ensayo y error), acumuló durante siglos gran conocimiento para el tratamiento de sus problemas de salud. Hoy día, el impacto de los productos naturales como ingredientes activos en productos farmacéuticos comerciales sigue siendo enorme. De los 252 fármacos considerados como básicos y esenciales por la Organización Mundial de la Salud (OMS), el 11% son exclusivamente originados de plantas [1]; además, los productos naturales y sus derivados representan más del 50% de los medicamentos de uso clínico en el mundo [2]. Igualmente, desde que Alexander Fleming descubrió por accidente la penicilina [3], los microorganismos comenzaron a ser fuente de fármacos, principalmente antibióticos y otros microbicidas [4]. A pesar de lo anterior, existe una fuente relativamente poco explorada en la búsqueda de compuestos activos antimicrobianos: los insectos. Su éxito evolutivo se ve reflejado en su número y hábitats colonizados durante 500 millones de años, tanto, que el número de especies de plantas y microorganismos juntos, son apenas la mitad de aquellos [5] Figura 1. Parte de la estrategia para alcanzar tal logro se encuentra en el desarrollo de poderosos sistemas de defensa contra infecciones [5].
Los insectos sociales hormigas, abejas, avispas y termitas viven en condiciones de hacinamiento en comunidades conformadas por enormes cantidades de individuos, en microambientes húmedos y tibios, que constituyen un ambiente ideal para las enfermedades infecciosas potencialmente letales para las colonias [6], pero estas no desaparecen; de hecho, la longevidad de una colonia está determinada por la vida de la reina, que puede extenderse hasta por quince años [7].
Las hormigas, uno de los cuatro insectos sociales, han creado y adaptado defensas químicas para sostener sus nidos. Las principales fuentes de estas armas químicas de defensa se localizan en las glándulas metapleurales (GMP) y las glándulas mandibulares (GM). Además, algunas especies, como las cortadoras de hojas (Atta spp., Acromyrmex spp.), viven de manera simbiótica con bacterias alojadas en su abdomen que producen agentes antisépticos.
Las GMP, cuyas aperturas se localizan justo por encima del punto donde el tercer par de patas articula con la parte trasera del tórax [8, 9], producen una secreción lipídica con componentes que se han identificado en algunas especies ácido acético, ácidos grasos de cadena corta, media y larga, γ-lactonas, γ-cetoácidos [10, 11]. Veal, Trimble y Beattie [12] evaluaron la actividad antimicrobiana de secreciones de esta glándula de la especie Myrmecia gulosa (hormiga toro australiana), encontrando actividad de amplio espectro sobre microorganismos Gram positivos, Gram negativos y hongos. Una especie en donde esta glándula se considera de especial importancia, es en la cortadora de hojas Acromyrmex octospinosus, ya que tales secreciones deben protegerlas a ellas y al hongo que cultivan en sus nidos y les sirve de alimento [13]. En sus secreciones se encontraron, aparte de los compuestos nombrados anteriormente, ácido indolacético y mymicacina [10, 13]. Tales secreciones mostraron actividad antimicrobiana sobre bacterias (Bacillus subtilis y Pseudomona stutzeri) y sobre hifas y esporas de varios hongos [11]. Sobre el abdomen de esta misma Acromyrmex, se identificó una bacteria simbiótica del género Streptomyces, capaz de producir antibióticos macrólidos tipo candicidina, activos contra el microhongo patógeno Escovopsis sp., que ataca al hongo Leucoagaricus sp., alimento cultivado por dichas hormigas en sus nidos sobre las hojas que cortan en el bosque [14]; igualmente, en la cutícula de estas, se encontró una bacteria del género Pseudonocardiaceae, productora de antibióticos activos contra el mismo patógeno [15].
La GM, que es constitutiva de las glándulas salivales [16], en muchas hormigas también produce agentes antisépticos. Brough (1983) [17] describió actividad tanto inhibitoria como bactericida de secreciones mandibulares de la especie Calomyrmex sp., secreciones de la GM de Atta sexdens rubropilosa, mostraron actividad inhibitoria sobre la germinación de conidios del hongo Botrytis cinerea Pers. Fr., hongo fitopatogénico de múltiples plantas de importancia económica [18]. En esta misma glándula, pero en la especie Pachycondyla goeldii, se identificaron una serie de 15 péptidos denominados ponericinas, con actividad antibacterial e insecticida [19].
El género Crematogaster está conformado por más de 780 especies y 85 subespecies que pertenecen a 15 subgéneros que se distribuyen por todo el mundo [20, 21]. La especie evaluada en este trabajo es de hábitat fundamentalmente arborícola; cabeza y tórax de color pardo oscuro y abdomen negro, con la característica terminación de este en punta, que se estrecha en el ápice tomando la apariencia de un corazón. Forman filas que siguen caminos invisibles durante días, por lo que la estrategia de captura consistió en aspirarlas de estos caminos. Las hormigas del género Solenopsis, llamadas comúnmente candelillas u hormigas de fuego, se distribuyen ampliamente por todo el mundo con más de 285 especies [21, 22], y han llegado a considerarse una verdadera plaga en muchas partes [23]. Aunque se alimentan principalmente de plantas tiernas, semillas e insectos muertos, pueden incluso atacar animales pequeños hasta matarlos, para lo cual emplean una ponzoña ubicada en el abdomen que inyecta un veneno típico del género, rico en alcaloides piperidínicos [24-26], responsables del característico dolor quemante de su picadura, y de múltiples reacciones anafilácticas en humanos [27, 28]. No es difícil encontrarlas en las basuras o alimentándose de animales muertos en descomposición. Este hecho en particular llamó la atención hacia esta investigación, ya que pueden caminar y alimentarse en lugares y con elementos muy contaminados microbiológicamente, sin que aparentemente esto afecte a la colonia. Pueden responden por esta realidad las secreciones de las GMP y GM, junto con la de su ponzoña de la que se han reportado actividades antibacterianas [29].
En el presente trabajo, se evaluó la actividad antibacteriana de extractos de hormigas de estos dos géneros, capturados en el casco urbano de la ciudad de Cartagena, con el propósito de evaluar su actividad antibacterial, y establecer si serían candidatas a estudios posteriores de separación e identificación de compuestos activos antimicrobianos.
METODOLOGÍA
La captura de la hormiga Crematogaster sp., conocida como "acróbata", se realizó en el casco urbano de la ciudad de Cartagena (10°21´06.42´´ N; 75°25´48.44´´ O. Altitud: 20 msnm), en un árbol de Tabebuia rosea (guayacán rosado) en el cual tienen su nido disperso en todo el tronco debajo de trozos de corteza seca, y por el que deambulan permanentemente. La Solenopsis sp., se capturó igualmente en la ciudad, en una unidad residencial (10°24´55.60´´ N; 75°31´56.00´´ O. Altitud: 5 msnm). Se instalaron trampas con cebos cárnicos, los cuales, al cabo de un par de horas, estaban cubiertos por una capa de hormigas. Capturadas, ambas especies se sacrificaron y los frascos y trampas colectoras se introdujeron en cajas de poliestireno expandido (Icopor) provistas de hielo seco. El gas carbónico y el frío desprendido de este, las anestesia y congela en una sola operación, preservando los componentes de interés [30, 31].
En el laboratorio las masas de hormigas se limpiaron de polvo, tierra, hojas pequeñas y todo elemento extraño, para proceder a secarlas en estufa a 40 °C (horno Memmert, UL 50) por 12 horas. Algunas unidades individuales fueron capturadas e introducidas en tubos plásticos pequeños con alcohol al 96%, y enviadas al Instituto de Ciencias Naturales de la Universidad Nacional de Colombia para su identificación (Voucher ZOO-016).
Los extractos etanólicos totales a partir de las hormigas secas se prepararon de la siguiente manera: se tomaron 10 gramos de hormigas secas de cada especie, y se maceraron con 50 mL de etanol 96% por 24 horas, a final de las cuales el alcohol se evaporó en estufa a 40 °C durante una semana hasta obtener un sólido marrón (Crematogaster) y un líquido aceitoso rojo (Solenopsis), que pesaron 253 mg y 3.000 mg para rendimientos del 2,53% y 30%, respectivamente, que se guardaron bajo refrigeración en tubos plásticos herméticos.
El ensayo actividad antibacterial se desarrolló sobre cepas de K. pneumoniae (ATCC 13883), P. aeruginosa (ATCC 17503), B. subtilis spizizenii (ATCC 6633) y S. aureus (ATCC 10832). Además, se utilizó una cepa de S. aureus aislada en el laboratorio de microbiología del Hospital de Bocagrande (Cartagena), resistente a los antibióticos Amoxicilina/Ac. Clavulánico, Ampicilina/Sulbactam y Ceftriazona/Oxacilina. Se verificó la viabilidad de cada uno de los microorganismos usados replicándolos en agar tripticasa de soya (TSA), e incubándolos durante 24 horas a 37 °C, para asegurar que las bacterias se encontraran en su fase de crecimiento exponencial. De allí, las colonias más homogéneas se suspendieron en caldo de tripticasa de soya (TSB) y se estandarizó la densidad bacteriana de cada cepa utilizando una solución estándar de turbidez de BaSO4, equivalente a 0,5 en la escala de McFarland (1,5 x 108 UFC/ml) [32]. La turbidez del estándar fue verificada en espectrofotómetro (Shimadzu UV-1700), a valores entre 0,08 – 0,10 a 625 nm. Cada caja de Petri con agar TSA dividida en cuatro partes, se sembró con el inóculo estandarizado con asa estéril frotando de ida y vuelta de un borde al otro garantizando la distribución homogénea del mismo.
Los extractos de las hormigas se diluyeron para evaluarse a diferentes concentraciones. En cabina microbiológica de flujo laminar, el extracto de Crematogaster se adicionó con 100 µL de dimetil sulfóxido (DMSO) y 400 µL de agua estéril, para obtener una solución madre de 506.000 ppm. Una alícuota se diluyó con agua estéril (AE) para obtener una solución a 100.000 ppm, en las que la concentración de DMSO no superó el 4%, de allí, se hicieron soluciones seriadas para obtener concentraciones de 50.000, 25.000 y 12.500 ppm. Soluciones menores no se pudieron preparar, pues por ser extractos tan lipofílicos, se enturbian al adicionar el AE. El extracto de Solenopsis se manejó de forma un poco diferente, debido a sus características aceitosas. 300 mg del aceite obtenido, se adicionaron con 10 µL de DMSO y se diluyó a la mitad con AE, pero al intentar hacer la siguiente disolución, esta se enturbiaba, de manera que este extracto solo se pudo trabajar puro y al 50%, ya que no era posible emplear cantidades mayores de DMSO, debido a su toxicidad sobre las bacterias.
Para la siembra de los extractos, se impregnaron 10 µL de cada dilución en discos de papel de filtro estériles de 5 mm de diámetro, que se dispusieron sobre las cajas recién inoculadas con cada una de las bacterias. Se incubó por 48 horas a 37 °C para determinar el halo de inhibición logrado por cada dilución. El ensayo se realizó por cuadruplicado, empleándose como control negativo discos de papel impregnados con 10 µL de DMSO al 4% (experimentalmente, se encontró que a esta concentración no se afectaron las bacterias) y gentamicina (60 µg/disco) como control positivo.
Se calcularon las medias de los halos de inhibición y se empleó el siguiente criterio cualitativo para determinar la actividad del extracto: (-) inactivo, ausencia de halo de inhibición; (±) poco activo, inhibición del crecimiento en el lugar de contacto del papel; (+) medianamente activo, halo de inhibición menor de 8 mm y (++) activo, halo de inhibición mayor de 9 mm [33].
RESULTADOS Y DISCUSIÓN
La aparición cada vez más frecuente de cepas resistentes a los antibióticos actuales preocupa cada vez más a la comunidad científica [34-36]; por tanto, la búsqueda de nuevos agentes antibióticos, sean de fuentes naturales o sintéticas, es una necesidad apremiante. De acuerdo con los resultados alcanzados, que se presentan en la Tabla 1, las hormigas pueden ser un buen punto de partida. Se evaluaron tres tipos de bacterias de reconocida patogenicidad (S. aureus, P. aeruginosa y K. pneumoniae) que presentan con gran frecuencia problemas a la comunidad médica y científica en general.
S. aureus es un patógeno que puede causar múltiples infecciones en individuos tanto sanos como inmunodeficientes. Su resistencia a los antibióticos comunes está en constante crecimiento, tanto que se estima que del 50% al 90% de las cepas asiladas presentan resistencia a la oxacilina y meticilina y muchas a la vancomicina [37-39]; resulta, por tanto, interesante encontrar que extractos de ambas especies de hormigas inhibieron el crecimiento de ambas cepas de S. aureus (ATCC 10832 y la resistente). En particular, el extracto puro de Solenopsis alcanzó un halo de inhibición sobre la ATCC comparable a la gentamicina, en tanto que los de Crematogaster fueron activos sobre la resistente a 100.000 y 50.000 ppm. P. aeruginosa es un germen oportunista que también está relacionado con enfermedades nocosomiales, ocasionando la mayoría de aislamientos principalmente en pacientes inmunodeprimidos; provocando múltiples infecciones [40]. Presenta preocupante resistencia a muchos antimicrobianos de uso clínico, incluyendo la mayoría de las penicilinas, las cefalosporinas de primera, segunda y tercera generación, las monobactamas, aminoglicósidos y fluoroquinolonas, entre otras [41, 42]. Los extractos de ambas hormigas fueron eficaces sobre ellas a niveles similares a los de la cepa anterior. Por su parte, K. pneumoniae, bacilo aerobio, también está asociado con infecciones nocosomiales en infantes prematuros y pacientes en cuidados intensivos; producen neumonías, infecciones urinarias e infecciones de heridas quirúrgicas [43, 44]. Sobre esta cepa, Crematogaster logró cierto grado de inhibición, en tanto que Solenopsis escasamente logró inhibir el crecimiento en el punto de contacto del papel con el agar inoculado. B. subtilis subsp. spizizenii, no suele ser una bacteria patógena sino benéfica; de hecho, se emplea como un probiótico que ayuda a regular la flora intestinal [45-47]. En este trabajo se empleó por ser una bacteria Gram positiva, pues se pretendía saber si los extractos son de amplio espectro (activos sobre gérmenes Gram positivos y negativos), lo cual resultó ser cierto.
Los resultados antibacterianos alcanzados por Crematogaster sp. son muy interesantes, pues su estilo de vida no parece estar relacionado con hábitats particularmente contaminados, aunque sí con la defensa de los mismos. De hecho, las hormigas obreras de la especie Crematogaster inflata, poseen las GMP más grandes entre las hormigas (respecto a su tamaño corporal), cuya secreción es usada como defensa química contra predadores [48]. Otra posibilidad puede estribar en el hecho de que las hormigas de este género conviven en mutualismo con ciertas plantas [49, 50]. El nido de la especie estudiada no se encontró en un agujero determinado en el guayacán rosado que correspondía a su hábitat, sino que estaba repartido debajo de secciones de corteza apartadas unas de otras, allí fueron hallados huevos al cuidado de sus hermanas; por tanto, es muy probable que esta hormiga proteja con sus secreciones al árbol para prevenirle infecciones, mientras que este les proporciona hospedaje y alimento. Los resultados encontrados sobre la Solenopsis sí eran más predecibles, dado sus hábitos carroñeros observados. Su extracto etanólico no se pudo llevar a sequedad, sino que se obtuvo un producto aceitoso de color rojo brillante, muy diferente al obtenido de Crematogaster. Esto significó dos cosas: que los agentes antimicrobianos son muy poco polares, y que el cuerpo de esta especie acumula más grasa que otras. Este material de aspecto lipídico no permitió hacer diluciones seriadas de concentraciones conocidas para siembras en agar, donde se requiere la migración de los compuestos en un medio polar; pero los resultados, positivos en todas las cepas bacterianas, son promisorios, pues estimamos que de llegar a sequedad, los compuestos secos estarían diez veces más concentrados.
Los extractos obtenidos de las hormigas analizadas, demostraron tener actividad antibacteriana tanto en bacterias Gram positivas como negativas, que merecen seguir siendo explorados. Por tratarse de extractos totales, los compuestos activos pueden encontrarse muy diluidos; además, por ser tan lipofílicos, su migración en el agar se pudo ver comprometida. No obstante lo anterior, los resultados son muy promisorios; por tanto, sería muy recomendable un fraccionamiento cromatográfico biodirigido para aislar el o los compuestos activos responsables de la actividad antibacterial encontrada.
AGRADECIMIENTOS
Los autores agradecen a la Universidad de Cartagena, entidad que hizo posible la realización del presente trabajo. Especialmente agradecemos la colaboración del profesor Fernando Fernández Castiblanco del Instituto de Ciencias Naturales de la Universidad Nacional de Colombia, por su apoyo en la identificación de las especies.
REFERENCIAS
1. S.M.K. Rates, Plants as source of drugs, Toxicon, 39, 603 (2001). [ Links ]
2. A. Gurib-Fakim, Medicinal plants: Traditions of yesterday and drugs of tomorrow, Mol. Aspects Med., 27, 1 (2006). [ Links ]
3. A. Fleming, Classics in infectious diseases: On the antibacterial action of cultures of a penicillium, with special reference to their use in the isolation of B. influenzae by Alexander Fleming (Reprinted from the British Journal of Experimental Pathology 10: 226-236, 1929), Rev. Infect. Dis., 2, 129 (1980). [ Links ]
4. L.A. Betancur-Galvis, A.C. Mesa-Arango, J.G. Bueno-Sánchez, Productos naturales con actividad antimicótica, Rev. Esp. Quimioter., 17, 325 (2004). [ Links ]
5. J.L. Dimarcq, I. Hunneyball, Pharma-entomology: When bugs become drugs, Drug Discov. Today, 8, 107 (2003). [ Links ]
6. N.H. Fefferman, J.F.A. Traniello, R.B. Rosengaus, D.V. Calleri, Disease prevention and resistance in social insects: Modeling the survival consequences of immunity, hygienic behavior, and colony organization, Behav. Ecol. Sociobiol., 61, 565 (2007). [ Links ]
7. J. Montoya, P. Chacón, M. Rosario, Caracterización de nidos de la hormiga arriera Atta cephalotes (Hymenoptera: Myrmicinae) en Cali (Colombia), Rev. Colomb. Entomol., 32, 151 (2006). [ Links ]
8. L.G. Gusmâo, F.H. Caetano, O. Nakano, Ultramorphology of the metapleural gland in three species of Atta (Hymenoptera, Formicidae), Iheringia, Ser. Zool., Porto Alegre, 91, 33 (2001). [ Links ]
9. A.S. Vieira, O.C. Bueno, M.I. Camargo-Mathias, The functional morphology of the metapleural gland of the leaf-cutting ant Atta laevigata (Formicidae: Attini), Micron, 41, 149 (2010). [ Links ]
10. D. Ortius-Lechner, R. Maile, E.D. Morgan, J.J. Boomsma, Metapleural gland secretion of the leaf-cutter ant Acromyrmex octospinosus: New compounds and their functional significance, J. Chem. Ecol., 26, 1667 (2000). [ Links ]
11. A.N.M. Bot, D. Ortius-Lechner, K. Finster, R. Maile, J.J. Boomsma, Variable sensitivity of fungi and bacteria to compounds produced by the metapleural glands of leaf-cutting ants, Insectes Soc., 49, 363 (2002). [ Links ]
12. D.A. Veal, J.E. Trimble, A.J. Beattie, Antimicrobial properties of secretions from the metapleural glands of Myrmecia gulosa (the Australian bull ant), J. Appl. Microbiol., 72, 188 (1992). [ Links ]
13. A.N.M. Bot, M.L. Obermayer, B. Hölldobler, J.J. Boomsma, Functional morphology of the metapleural gland in the leaf-cutting ant Acromyrmex octospinosus, Insectes Soc., 48, 63 (2001). [ Links ]
14. S. Haeder, R. Wirth, H. Herz, D. Spiteller, Candicidin-producing Streptomyces support leaf-cutting ants to protect their fungus garden against the pathogenic fungus Escovopsis, Proc. Natl. Acad. Sci. USA, 106, 4742 (2009). [ Links ]
15. M. Poulsen, A.N. Bot, J.J. Boomsma, The effect of metapleural gland secretion on the growth of a mutualistic bacterium on the cuticle of leaf-cutting ants, Naturwissenschaften, 90, 406 (2003). [ Links ]
16. L.F. Pavon, M.I. Mathias, Ultrastructural studies of the mandibular glands of the minima, media and soldier ants of Atta sexdens rubropilosa (Forel 1908) (Hymenoptera: Formicidae), Micron, 36, 449 (2005). [ Links ]
17. E.J. Brough, The antimicrobial activity of the mandibular gland secretion of a formicine ant, Calomyrmex sp. (Hymenoptera: Formicidae), J. Invertebr. Pathol., 42, 306 (1983). [ Links ]
18. A.L.M. Junior, T.M.C. Della Lucia, L.C.A. Barbosa, L.A. Maffia, M.A.B. Morandi, Efeito de secreções da glândula mandibular de Atta sexdens rubropilosa Forel (Hymenoptera: Formicidae) sobre a germinação de conídios de Botrytis cinerea Pers. Fr, Neotrop. Entomol., 30, 403 (2001). [ Links ]
19. J. Orivel, V. Redeker, J.P. Le Caer, F. Krier, A.M. Revol-Junelles, A. Longeon, A. Chaffotte, A. Dejean, J. Rossier, Ponericins, new antibacterial and insecticidal peptides from the venom of the ant Pachycondyla goeldii, J. Biol. Chem., 276, 17823 (2001). [ Links ]
20. M.G. Karaman, New data of the genus Crematogaster Lund, 1831 (Hymenoptera: Formicidae) in Crna Gora (Montenegro), Part I: Crematogaster scutellaris group of species, Turk. Entomol. Derg., 34, 419 (2010). [ Links ]
21. B. Bolton, G. Alpert, P.S. Ward, P. Naskrecki, "Bolton´s catalogue of ants of the world, 1758-2005", Harvard University Press, Boston, 2006. [ Links ]
22. M.R. Sharaf, A.S. Aldawood, First occurrence of Solenopsis Westwood 1840 (Hymenoptera: Formicidae), in the kingdom of Saudi Arabia, with description of a new species S. saudiensis n. sp., Ann. Soc. Entomol. Fr., 47, 474 (2011). [ Links ]
23. D.H. Oi, S.M. Valles, Fire ant control with entomopathogens in the USA. En "Use of microbes for control and eradication of invasive arthropods", editado por A.E. Hajek, T. Glare, M. O'Callaghan, Springer, New York, 2009. [ Links ]
24. M.S. Blum, J.M. Brand, R.M. Duffield, R.R. Snelling, Chemistry of the venom of Solenopsis aurea (Hymenoptera: Formicidae), Ann. Entomol. Soc. Am., 66, 702 (1973). [ Links ]
25. L. Chen, H.Y. Fadamiro, Re-investigation of venom chemistry of Solenopsis fire ants. II. Identification of novel alkaloids in S. invicta, Toxicon, 53, 479 (2009). [ Links ]
26. L. Chen, H.Y. Fadamiro, Re-investigation of venom chemistry of Solenopsis fire ants. I. Identification of novel alkaloids in S. richteri, Toxicon, 53, 469 (2009). [ Links ]
27. M.S. La Shell, C.W. Calabria, J.M. Quinn, Imported fire ant field reaction and immunotherapy safety characteristics: the IFACS study, J. Allergy Clin. Immunol., 125, 1294 (2010). [ Links ]
28. G.O. Solley, C. Vanderwoude, G.K. Knight, Anaphylaxis due to red imported fire ant sting, Med. J. Aust., 176, 521 (2002). [ Links ]
29. D.P. Jouvenaz, M.S. Blum, J.G. MacConnell, Antibacterial activity of venom alkaloids from the imported fire ant, Solenopsis invicta Buren, Antimicrob. Agents Chemother., 2, 291 (1972). [ Links ]
30. R.L. Patton, L.J. Edwards, S.K. Gilmore, Delivering safe levels of CO2 for insect anesthesia, Ann. Entomol. Soc. Am., 61, 1046 (1968). [ Links ]
31. T.L. Nilson, B.J. Sinclair y S.P. Roberts, The effects of carbon dioxide anesthesia and anoxia on rapid cold-hardening and chill coma recovery in Drosophila melanogaster, J. Insect. Physiol., 52, 1027 (2006). [ Links ]
32. S. Sutton, Measurement of cell concentration in suspension by optical density, Pharmaceutical Microbiology Forum Newsletter, 12(8), 3 (2006). [ Links ]
33. D.P. Jouvenaz, M.S. Blum, J.G. MacConnell, Antibacterial activity of venom alkaloids from the imported fire ant, Solenopsis invicta Buren, Antimicrob. Agents Chemother., 2, 291 (1972). [ Links ]
34. Y. Cabrera, A. Fadragas, L. Guerrero, Antibióticos naturales: mito o realidad, Revista Cubana de Medicina General Integral, 21, 3-4 (2005). [ Links ]
35. J. Gérvas, La resistencia a los antibióticos, un problema de salud pública, Atención Primaria, 25, 147 (2000). [ Links ]
36. C.M. Kunin, Why did it take the Infectious Diseases Society of America so long to address the problem of antibiotic resistance?, Clin. Infect. Dis., 46, 1791 (2008). [ Links ]
37. D.M. Bamberger, S.E. Boyd, Management of Staphylococcus aureus infections, Am. Fam. Physician, 72, 2474 (2005). [ Links ]
38. C.A. Mendoza Ticona, R. Velásquez Talavera, L. Mercado Díaz, J. Ballón Echegaray, C. Maguiña Vargas, Susceptibilidad antimicrobiana de Staphylococcus aureus sensible, con sensibilidad "BORDERLINE" y resistentes a la meticilina, Rev. Med. Hered., 14, 181 (2003). [ Links ]
39. E. Velazco, B. Nieves, M. Araque, Z. Calderas, Epidemiología de infecciones nosocomiales por Staphylococcus aureus en una unidad de alto riesgo neonatal, Enferm. Infecc. Microbiol. Clin., 20, 321 (2002). [ Links ]
40. N. Mesaros, P. Nordmann, P. Plesiat, M. Roussel-Delvallez, J. Van Eldere, Y. Glupczynski, Y. Van Laethem, F. Jacobs, P. Lebecque, A. Malfroot, Pseudomonas aeruginosa: Resistance and therapeutic options at the turn of the new millennium, Clin. Microbiol. Infect., 13, 560 (2007). [ Links ]
41. G. Delgado, A.D. García Mayorgas, F. Rodríguez, A. Ibarra, M. Casal, Sensibilidad y resistencia de Pseudomonas aeruginosa a los antimicrobianos, Rev. Esp. Quimioter., 20, 230 (2007). [ Links ]
42. D.M. Livermore, Multiple mechanisms of antimicrobial resistance in Pseudomonas aeruginosa: Our worst nightmare?, Clin. Infect. Dis., 34, 634 (2002). [ Links ]
43. J.R. Hernández, A. Pascual, R. Cantón, L. Martínez-Martínez, Grupo de Estudio de Infección Hospitalaria, Escherichia coli y Klebsiella pneumoniae productores de betalactamasas de espectro extendido en hospitales españoles (Proyecto GEIH-BLEE 2000), Enferm. Infecc. Microbiol. Clin., 21, 77 (2003). [ Links ]
44. P. Martínez, M. Mercado, S. Máttar, Determinación de b-lactamasas de espectro extendido en gérmenes nosocomiales del Hospital San Jerónimo, Montería, Colombia Médica, 34, 196 (2009). [ Links ]
45. G. Dehghan-Noudeh, M. Housaindokht, B.S.F. Bazzaz, Isolation, characterization, and investigation of surface and hemolytic activities of a lipopeptide biosurfactant produced by Bacillus subtilis ATCC 6633, J. Microbiol., 43, 272 (2005). [ Links ]
46. M.R. Oggioni, G. Pozzi, P.E. Valensin, P. Galieni, C. Bigazzi, Recurrent septicemia in an immunocompromised patient due to probiotic strains of Bacillus subtilis, J. Clin. Microbiol., 36, 325 (1998) [ Links ]
47. T. Stein, Bacillus subtilis antibiotics: Structures, syntheses and specific functions, Mol. Microbiol., 56, 845 (2005). [ Links ]
48. J. Billen, R. Hashim, F. Ito, Functional morphology of the metapleural gland in workers of the ant Crematogaster inflata (Hymenoptera, Formicidae), Invertebr. Biol., 130, 277 (2011). [ Links ]
49. S.P. Quek, S.J. Davies, T. Itino, N.E. Pierce, Codiversification in an ant-plant mutualism: Stem texture and the evolution of host use in Crematogaster (Formicidae: Myrmicinae) inhabitants of Macaranga (Euphorbiaceae), Evolution, 58, 554 (2004). [ Links ]
50. W.F. Wood, T.M. Palmer, M.L. Stanton, A comparison of volatiles in mandibular glands from three Crematogaster ant symbionts of the whistling thorn acacia, Biochem. Syst. Ecol., 30, 217 (2002). [ Links ]
