











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCTION
Infectious diseases remain among the leading causes of morbidity and mortality in the world. Moreover, antimicrobial (AMR) and multidrug (MDR) resistance have risen to alarming levels in Staphylococcus aureus (i.e., chiefly to penicillin and cross-resistance to other ß-lactams). From the global burden associated with AMR/MDR in microorganisms, most frequent are the threat to therapeutics, prolonged illness, decreased effectiveness, creating easy targets for immunocompromised conditions, high medical costs, and high mortality rates [1-4].
Indeed, the accelerating growth and global expansion of oxacillin (OXA) resistant S. aureus (ORSA) strains and hospital (HAI) or community (CAI) associated infections related to them are ultimately both a global public health problem and a critical problem in clinical settings where immunocompromised patients or those who have had surgical wounds are present. Resistance to current antibiotics has limited the therapeutic armory to treat S. aureus infections [6, 7].
In that way, approaches such as screening of plant products alone or in combination with antibiotics that can tackle AMR/MDR have been extensively reported [7-11]. Plants have been used in folk medicine since ancient times, and the chemical diversity and therapeutic potential are great reasons for the current interest taken in them [10]. Talinum paniculatum (Jacq.) Gaertner (Talinaceae family) is a plant widely spread throughout the world, commonly known as "Erva-gorda" and "Língua-de-vaca" in Brazil or Tu-ren-shen in Traditional Chinese Medicine [12]. Its green leafy is a non-conventional vegetable for human consumption [12]. Furthermore, several biological activities have been attributed to T. paniculatum and its bioactive compounds, including antibacterial and antifungal [7], estrogenic [13], antifertility [14], antinociceptive [15], and other effects: on uterine contractility and those related in folk medicine [12, 16, 17].
Over the last two decades, the underlying mechanism of action for the bactericidal ß-lactam antibiotics (including OXA) against a broader set of microorganisms has been shown to be overt dependent of reactive oxygen species (ROS) accumulation [18-20]. On this knife's edge, OXA-induced ROS-associated damage to proteins and/ or DNA contribute to S. aureus loss of viability, even when there is increased expression of systems of ROS detoxification (i.e. condition that characterizes the so-called oxidative stress) [21-23]. Indeed, ß-lactams (including OXA) can generate oxidative stress-induced autolysis. Besides these factors, oxidative stress tolerance-induced antibiotic resistance in S. aureus biofilms with a maintenance of the steady-state levels of ROS in S. aureus cells, has been described [21-27].
Despite the controversy and opposing viewpoints over the mechanism of ROS accumulation behind the activity of antibiotics, OXA activity could be hampered by providing microbial cells with exogenous antioxidants, as attested in previous studies [20, 28-31]. Moreover, antioxidants could prime (priming) microbial cells to counteract the oxidative stress induced by antibiotics [1, 22, 24, 32, 33]. Therefore, the elucidation of possible interactions between antioxidants-rich foods and antibiotic is important because of many nutrition products and medicines, including the leaves from T.paniculatum that are often used as a green leafy vegetable for human consumption, may occur during medical treatment and modify the action of the antibiotic hence this could have a great impact on clinical practice and patient outcomes [7, 12].
In this study, we evaluated the antistaphylococcal potential of the extract and fractions from the leaves of T. paniculatum alone or in association with the ß-lactam antibiotic OXA against ORSA and OXA-sensitive S. aureus (OSSA). Furthermore, a possible toxicity of T. paniculatum (in vivo) was checked.
MATERIALS AND METHODS
Chemicals
Oxacillin (OXA); Mueller Hinton broth (MHB); 7-Hydroxy-3H-phenoxazin-3-one 10-oxide (Resazurin) were purchased from Sigma-Aldrich, St. Louis, MO, USA. Nutriente Agar (Difco); Agar/broth Brain Heart Infusion (BHI, HiMedia Laboratories); Dimethyl sulfoxide (DMSO, Neon Comercial); Solutions: Sodium chloride 0.9% (145 mM NaCl); Phosphate-buffered saline (PBS, pH 7.4): NaCl (120 mM), KH2PO4 (1.7 mM), Na2HPO4 (8.3 mM) e KCl (5 mM). Other chemicals and solvents were analytical grade. The reagents were sterilized into an autoclave (whenever possible) or by filtration (Millipore Corporation, hydrophilic Durapore* PVDF, 0.22 μm, Ø 47 mm).
Ethics statement
This study did not involve any endangered or protected species and no specific permits were required for the described studies. Botanical material from T. paniculatum was collected in a particular area, with access permitted to researchers.
All animal experiments were carried out in strict accordance with the recommendations of the Guide for the Care and Use of Laboratory Animals (National Institutes of Health [NIH], Washington DC: The National Academy Press, 2011). The animals were gently conceded from a previous approved study by the ethics committee on the use of animals (CEUA) of the Federal University of Alfenas (protocol #490/2013).
Plant material and preparation of the crude leaf extract and its fractions
T. paniculatum leaves were collected in the mornings of June, near the town of Fama, MG state, Brazil (Geographic coordinates obtained from GPS observations: 21° 24' 53.4" S; 45° 52' 15.8'' W). The climate of this region is classified as humid temperate, with a hot summer and a dry winter (type Cwa in the classification of Köppen). A voucher specimen was deposited in the UALF Herbarium at Federal University of Alfenas (UNIFAL-MG), after identification and taxonomic authentication by a botanical (Herbarium number 2338). The collected T. paniculatum leaves were weighed, arranged in thin layers, and dried in a circulating air oven at 40 °C for eleven days (250 h), aiming at a stable and constant weight to be reached. The final percentage of dry mass was 5.26%. The material underwent through a rough division followed by a spray obtained in a Wiley mill. After grinding, the size of the powder particles was measured according to WHO, as shown in table 1 [34]. The dried leaf powder (200 g) from T. paniculatum was percolated (with an alcohol to water ratio of 7:3) at 1.0 mL/min/kg. After fluid extraction, the fluid was placed in a rotary evaporator under reduced pressure and at a temperature of 45 °C. The extract was then lyophilized to completely remove water and obtain the dry extract, hereafter referred to as CLE (17.0% yield). To obtain the fractions, the dried extract was subjected to a liquid-liquid partition with hexane (1:1, v/v [4X]) and ethyl acetate (1:1, v/v [6X]), yielding hexane (HX), ethyl acetate (EtOAc), and a final residue termed the aqueous fraction. During this process, the solvents, hexane, and ethyl acetate, are removed. The CLE or fractions were solubilized in DMSO (which at used concentration does not present antimicrobial activity) and adjusted at used concentrations.
Strains
Reference strains of S. aureus were from the American Type Culture Collection (Staphylococcus aureus subsp. aureus [ATCC® 25923™ and 6538], Manassas, VA, USA). The two ORSA isolates were from clinical environment and identified as shown in table 2. The classification of AMR in S. aureus was as following: OXA MIC ≤ 2 μg ml-1= Sensitive and ≥ 4 μg ml-1= resistant [35].
Table 2 Used microorganisms, clinical sources, identification, and code.
AInsertion sequence.
Genomic DNA: Clinical isolates of S. aureus was grown in Luria-Bertani medium (pH 7, Laboratory Bottles, Hamburg, Germany) for 16 hrs at 35 °C with continuous shaking (150 r.p.m). Then, bacteria were washed two times with Tris-buffered saline (TBS: 50 mM Tris HCl pH 8.0, 100 mM EDTA, 150 mM NaCl), followed by a centrifugation at 1500 X g (Centrifuge 5417R, Eppendorf do Brasil Ltda., São Paulo, SP, Brasil) for 5 min. The resulting S. aureus cells containing-pellet was incubated at 37 °C for 30 min and cells were lysed in 500 μL TBS containing SDS (1% m/v), 100 μg lysostaphin (Sigma Chemical Co., St. Louis, MO, USA) and 100 μg RNAse (Sigma Chemical Co., St. Louis, MO, USA) into Eppendorf Thermomixer® comfort (Eppendorf™, Hamburg, Germany). The final lysed S. aureus cells were added with 200 μg proteinase K (Sigma Chemical Co., St. Louis, MO, USA) and 200 μg lysozyme (Sigma Chemical Co., St. Louis, MO, USA). Next, after the addition of phenol:chloroform:isoamyl alcohol (25:24:1), a high-speed centrifugation (12000 ' g for 10 min) was performed and the culture supernatants underwent a further centrifugation, and then resuspended in chloroform:isoamyl alcohol (24:1). Genomic DNA was pelleted by subsequent addition of ethanol at -20 °C for 30 min and centrifugation at 12000 X g for 10 min, dried at 30 °C for 90 min, and solubilized in 500 μL TE buffer (10 mM Tris, 1 mM EDTA, pH 8) and stored at -20 °C. PCR multiplex assays - PCR was performed using the following set up: 100 ng genomic DNA in assay buffer (20 mM Tris HCl pH 8.4, 50 mM KCl), 1.5 mM MgCl2, 0.2 mM of each dNTP, 100 pmol of each primer (described in table 1), and 1.5 U Taq DNA polymerase (Invitrogen™ Brasil Ltda.) were mixed and underwent to an initial program for DNA denaturation at 92 °C for 3 min, followed by 30 cycles at 92 °C for 1 min (denaturation), 56 °C for 1 min (annealing) and 72 °C for 1 min (extension). The final cycle was performed at 72 °C for 5 min for the final extension. Amplicons patterns - The PCR products in loading buffer (10% v/v glycerol, 0.083% m/v bromophenol blue, 0.083% m/v xylene cyanol in H2O milliQ) were analyzed on a 2% agarose gel (prepared with TBE buffer 0.5': 445 mM trizma base, 445 mM boric acid, and 10 mM EDTA) and 0.5 μg mL-1 ethidium bromide) using horizontal gel electrophoresis apparatus (Sunrise™ 96), with a voltage at 5 V cm-1 of gel length (Thermo EC 3000-90 power supply, Thermo Electron Corporation, Milford, MA, United States) and a continuous gradient (TBE buffer 0.5 X ) for 3 hrs. The gels were visualized by UV light at 300 nm (Electronic UV Transilluminator mod. EB-40, Ultra Lum. Inc., Paramount, CA). Images were captured with a Photodocumentation System (Mod. DP001. FDC version 10; Vilber Lourmat, Marne la Vallée, France) and analyzed with a PhotoCapt MW (version 10.01 for Windows; Vilber Loumat, Marne la Vallée, France). S. aureus ATCC® 43300 (methicillin resistant S. aureus-MRSA), ATCC® 25923 (methicillin sensitive S. aureus-MSSA) and a maker of DNA (100 bp DNA, Ladder Cat. # 15628-019, Invitrogen) were used as controls.
Minimum Inhibitory Concentration (MIC) and determination of interactions between plant product and OXA
The MIC values were determined using the broth checkerboard microdilution assay on 96-well microplates (flat-bottom, Corning Inc., NY) containing 100 mL of MHB, following the CLSI document (M7-A6) [35, 36]. CLE or fractions (EtOAc or HX) were diluted into the wells (at final concentrations ranging from 4000 to 1.95 μg ml-1 [CLE], and from 500 to 0.244 μg ml-1 [fractions]), either alone or coupled with OXA (from 8000 to 3.9 μg ml-1). S. aureus suspensions (cultures of ORSA [isolates] or OSSA [ATCC® 25923™] during the early exponential growth phase, after an overnight incubation on BHI) were centrifuged (900 X g for 6 min), the pellet diluted in sodium chloride 0.9%, and then spectrophotometrically adjusted at OD625 = 0.1 = 1 X 108 CFU mL-1 (further diluted, 1 X 105 CFU mL-1 final concentration). Afterwards, 10 μL of inoculums were added to the wells. The plates were then incubated at 37 °C for 24 hours. Finally, readings were performed visually as previously determined [7], wherein the presence of turbidity in the wells was considered indicative of microbial growth and a specific dye (resazurin 0.01%) was used to evaluate the metabolic activity of the microorganism. The MIC of the CLE, fractions (EtOAc or HX) or OXXA alone, and all isoeffective combinations (association between plant product and OXA) were defined when the growth of the microorganism was inhibited at the lowest concentration. The growth control was composed of 100 μL of MHB and 10 μL of inoculum. The extract control was composed of 100 μL of MHB and 100 μL of the CLE or fraction (EtOAc or HX) and the sterility control contained only 100 μL of MHB.
Minimum Bactericidal Concentration (MBC)
The MBC was defined as the lowest concentration of OXA/extract/fractions that killed 99.9% of the original inoculums from each well where growth inhibition occurred in the analysis of MIC/Synergism. For each strain of ORSA and OSSA, MBC values were determined by removing 100 µ of microbial suspension from each well demonstrating no microbial growth and inoculating them with three serial dilutions of 1:10 on Petri plates (90 X 15 mm) containing the nutrient agar. Then, the plates were incubated at 35 °C for 24 h. After that, colony counts were performed to determine which concentrations showed bactericidal or bacteriostatic effect.
Toxicity test: in vivo acute toxicity
Adult male Swiss mice weighing 30 to 45 g were obtained from the Central Animal Facility of the Federal University of Alfenas and housed in a controlled 12 h light/ dark cycle at 23 °C and received water and food ad libitum. The acute toxicity in mice (n = 10) was performed with an oral dose of 5 g kg-1 of the CLE. During a seven-day period with adequate acclimatization (21 °C) and food and water given to the mice, ad libitum, the following parameters were assessed: hyperactivity, sedation, changes in stool characteristics (consistency), and food and water intake [37].
Data analysis
MIC and MBC values were from two replicate wells from two independent experiments. To assess the interactions between the CLE, HX, or EtOAc and OXA, the data obtained from the checkerboard microdilution assays were analyzed as previously described. The model-fractional inhibitory concentration index (FICI) was calculated as follows: FICA = MICAB/MICA and FICB = MICBA/MICB, where MICA and MICB are the MICs of samples A and B when acting alone, and MICAB and MICBA are the MICs of samples A and B when acting in combination, respectively. A and B represent the treatments (exposures) under investigation (i.e., A: can be an extract or fraction and B: OXXA); then, FICI = FICA + FICB. "Synergy" was defined as an FICI ≤ 0.5, while "antagonism" was defined as an FICI value > 4.0. An FICI between 0.5 and 1.25 was considered to have an "additive effect" and between 1.25 and 4 "no interaction" [38, 39]. According to the ratio of MBC/MIC, the type of antimicrobial action displayed by each treatment was considered. If the ratio of MBC/MIC = 1 or 2, the effect was considered bactericidal, but if the ratio of MMC/MIC = 4 or 16, the effect was defined as bacteriostatic [40]. The selectivity index (SI) was calculated as follows: SI = CC90/MIC9W, being CC90 adopted from our previous study [7].
RESULTS
MIC and MBC values for OXA and the CLE, EtOAc, and HX from T. paniculatum
The MIC and MBC values for OXXA and CLE, EtOAc, and HX against OSSA and the two clinical samples are shown in table 3. Increased MIC values were demonstrated for OXA against the isolates (samples G11.39 and G11.19, both resistant to this antibiotic [MIC ≥ 4 μg ml-1]), confirming the ORSA profiles. We found MIC values of 500 μg ml-1 for EtOAc and HX and 4000 μg ml-1 for CLE against both OSSA and ORSA. The CLE was as effective as OXXA against one of the isolates (sample G11.39). Both EtOAc and HX were more effective (lower MICs) than OXXA against the two ORSA isolates.
Table 3 Minimum inhibitory (MIC) and minimum bactericidal (MBC) concentrations for the crude leaf extract (CLE) and fractions from Talinum paniculatum and for Oxacillin against Staphylococcus aureus and the types of antibacterial effects.
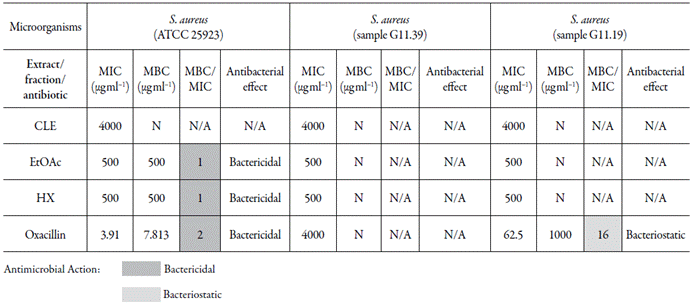
For plants products: MIC up to 100 μg ml-1 = promising inhibitory potential, MIC between 100 and 625 μg ml-1 = moderate inhibitory activity [7]. Fractions from the CLE of Talinum paniculatum; EtOAc = Ethyl acetate fraction and HX= Hexane fraction; S. aureus: samples G11.39 and G11.19 are from clinical envi ronment; S. aureus: 5 ´ 105 CFU mL-1; Antimicrobial resistance (AMR) classification: OXA MIC ≤ 2 μg ml-1 = sensitive and ≥ 4 μg ml-1 = resistant [35]. Ratio MBC/MIC = 1 or 2 → Effect was considered as bactericidal; Ratio MBC/MIC = 4 or 16 → Effect was defined as bacteriostatic [40]. N: inhibition of microbial growth was not observed at maximum used concentration; N/A: not applicable due to absence of an MBC value.
Antistaphylococcal effects for the CLE and fractions
Tn table 3, it is shown that the fractions EtOAc and HX showed bactericidal effects against OSSA and ORSA. MBC values were not observed for the CLE and fractions at maximum concentrations used in this study, 4000 μg ml-1 and 500 μg ml-1, respectively, against ORSA strains.
Final MIC values for the associations between plant product (CLE, EtOAc, and HX) and OXA, and FICI values from these associations
The associations between plant products (CLE, EtOAc, or HX from T. Paniculatum) and OXA, were investigated. As demonstrated in table 3, the treatment of OSSA or ORSA with CLE, EtOAc, or HX, resulted in outstanding MTC values (low MTC values). On the other hand, when the association between plant product (extract/ fraction) and OXA was performed (table 4), OXA MTCs increased 2- to 4092-fold, whereas the MTCs of each fraction decreased up to 1020-fold. Taken together, data from tables 3 and 4 were interpreted as FTCT values (table 4), which demonstrated that there was no synergy for any of the associations. The most common effect for the associations was "no interaction" (8 associations).
Table 4 Results for the contribution of each compound or plant product in the final MIC value during the association between (1) plant product (Crude leaf extract [CLE], EtOAc, or HX) from Talinum paniculatum and (2) Oxacillin against Staphylococcus aureus, correspondent Fractional inhibitory concentration index (FICI), and interpretation of the FICIs from the associations with the resulting type of interaction.
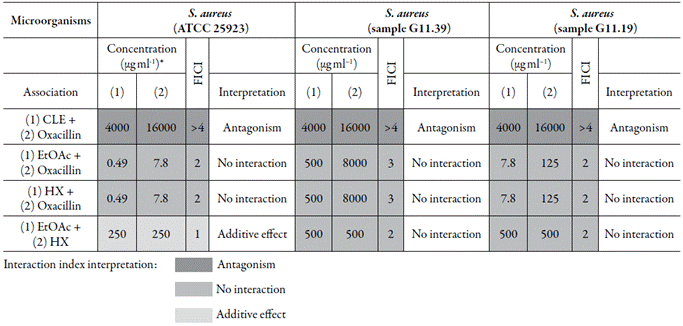
Fractions from the CLE of Talinum paniculatum; EtOAc = Ethyl acetate fraction and HX = Hexane fraction; *Concentration of each substance in the associations between the (1) crude leaf extract (CLE)/fraction (EtOAc, or HX) from Talinum paniculatum and (2) oxacillin or (1) fraction and (2) fraction in which no microbial growth was observed; FICI data interpretation from the interactions/associations: ‘synergy’ (FICI ≤ 0.5), “Additive effect” (FICI > 0.5-1.25) ‘no interaction’ (FICI > 1.25-4.0), and ‘antagonism’ (FICI > 4.0) [38-39].
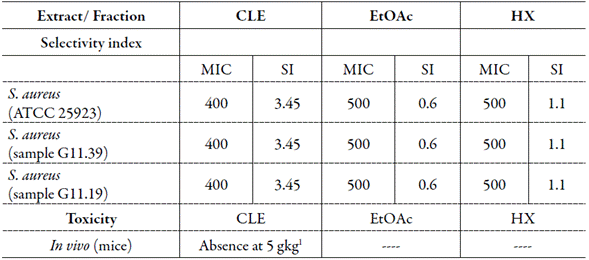
In vivo/in vitro toxicity data for T. paniculatum and selectivity index (SI)
Of clinical relevance, the CLE from T. paniculatum did not present an acute toxicity at a dose of 5 g kg-1 (table 5). Tn table 5, the results of the ST are also presented, and the best ones were from CLE against both OSSA (3.45) and ORSA (3.45).
DISCUSSION
Tn this study, we first demonstrated that the CLE from T.paniculatum and its fractions (EtOA and HX) have antibacterial activity against OSSA and ORSA, and a bactericidal effect for the fractions against OSSA. Currently, resistance to ß-lactam antibiotics in S. aureus is a global and serious health threat, even in the clinical environment such as the healthcare settings, places where the ORSA strains used in this study were isolated [4, 41, 42]. Thus, new antimicrobials are urgently needed.
Over the last few decades, the screening of plants seems to be a good option for possible sources of new compounds with original antistaphylococcal action, including studies that evaluate the association between plant product and antibiotic [7, 43-45]. T. paniculatum was here evaluated because of previous studies about the antimicrobial activity against medically relevant strains of bacteria and fungi; the popular use in folk medicine, and the culinary use [7]. The antimicrobial activity for members of the Talinaceae family or related families have also been described [46, 47].
As for the selectivity toward microbial cells, data have demonstrated a low toxicity for T. paniculatum leaves on other cells in vitro, justifying the antimicrobial potential of this plant [7]. We showed the selectivity of plant products from T.paniculatum toward S. aureus cells. Moreover, we found absence of an in vivo toxicity for T. paniculatum leaves at a dose of 5 mg kg-1.
The screening for the antimicrobial activities of plant products alone, or in association with antibiotic, have been reported in myriad papers that document the effects against different microorganisms, including human pathogens [3, 30, 39, 40, 45, 48-51]. As seen in this study, decreased OXA lethality was observed for the association between plant products and antibiotic, and our results are in line with some previous reports [20, 31, 52]. About this context, the "one drug, one target, one disease" approach has for some time remained the conventional pharmaceutical approach to the search of drugs and treatment strategies. However, this paradigm has been gradually shifted toward the adoption of combination therapies as well as possible interactions between drug and food.
Contrarily to a desirable synergistic action between plant product and OXA, in this study was demonstrated that such associations caused an augmentation in OXA MTCs. Tn this context, a putative action of antioxidants-containing plant product upon ROS levels should be considered. The action of some antibiotics have shown dependence on intense ROS accumulation and associated oxidative stress (i.e., OXA produces ROS that promotes growth inhibition or kills S. aureus), and other studies have pointed that antioxidants-containing plant products and isolated antioxidant compounds can decrease the susceptibility of microorganisms to antimicrobials, including S. aureus to ß-lactam and other antistaphylococcal drugs [20, 31, 52-55].
Other evidences indicate a role of harnessing oxidative stress as an interesting way to enhance the killing efficacy of OXA and other antistaphylococcal drugs, regardless of macromolecular or drug-target interaction (the main antibiotic mode-of-action in interfering with cell wall biosynthesis), aside from the importance of new insights into the mechanisms behind signaling pathways in S. aureus to control the redox balance and develop high-level AMR/MDR [23, 56, 57]. A metabolic response of S. aureus toward oxidative stress caused by penicillin has been proposed through up-regulation of an environmental sensing and other gene, countering oxidative damage coupled with other effects generated by this ß-lactam antibiotic [29, 58]. Moreover, a successful response of S. aureus vis-à-vis oxidative stress imposed by antibiotics includes overproduction of antioxidant pigments, overexpression of detoxifying enzymes (such as superoxide dismutase [Sod], catalase and peroxiredoxin AhpC) and genes and enzymes related to DNA protection and repair (including mrgA response and pyrophosphohydrolase activity of MutT to remove oxidized guanine [8-oxoG or GO lesion] from the nucleotide pool), and an augmentation in protein damage repair (overexpression of thioredoxin).
It is well-known that ß-lactam antibiotics induce an increase in oxygen consumption to form superoxide (O2 ●-) as a by-product via different pathways (such as the metabolism-related NADH depletion, the tricarboxylic acid (TCA) cycle, the electron transport chain, damage of iron sulphur clusters in proteins, and stimulation of the Fenton reaction). The production of O2 ●- allows the formation of other ROS, including hydrogen peroxide (H2O2), and the derivatives HO● (via Fenton chemistry) and/or peroxinitrite (ONOO-, via nitric oxide [●NO] biosynthesis pathways [58-60]. Moreover, while S. aureus Sod and catalases can enzymatically degrade O2 ●- and H2O2, respectively, no cellular detoxification mechanism is described for HO', and this oxidant can avidly attack the DNA [59-61]. Thus, high-levels HO● and the effects of blocking HO● accumulation (controls) can interfere with MTC and MBC values of antistaphylococcus agents (including OXA), as previously reported [20]. Tn this context, these factors may, at least partially, explain the link between HO● accumulation and oxidative DNA damage, as seen to a broader set of antibiotics and against different microorganisms [59-65], also explaining how antibiotic lethality is enhanced by amplifying basal ROS production and how antioxidants containing-plant products can interfere with this process [53]. Tn that way, we have reasons to believe that the prevention of OXA-induced ROS-mediated growth inhibition or killing of S. aureus is related to a mechanism in which the levels/concentrations of compounds in T. paniculatum generate an effect "purely antioxidant" within S. aureus cells.
Taken together, our findings suggest that the alone CLE, HX and EtOAc from T. paniculatum present antistaphylococcal potential for furthers studies, in vitro involving other strains of S. aureus and in vivo. However, a word of warning should be issued regarding the in vivo association between T. paniculatum and OXA and the use of T. paniculatum should be phased out during OXXA treatment. These in vitro effects carry in vivo implications and caution is required, since patients under OXXA treatment parallel to the intake of leaves or herbal drug preparations from T. paniculatum may cause a decrease in OXA efficacy (jeopardizing therapeutic). Our data supports the fact that antioxidants containing-plant products, being a part of many nutrition products and medicines, may interact with OXXA during antibiotic therapy and modify its action, causing resistance phenomenon or the failure of therapeutic regimens. However, to support this idea, these data should be investigated further, and the physician should take all this into account.

