










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkRevista Colombiana de Psiquiatría
versão impressa ISSN 0034-7450
rev.colomb.psiquiatr. v.33 supl.1 Bogotá out. 2004
ARTÍCULO DE REVISIÓN
EL CEREBRO: DE LA ESTRUCTURA Y LA FUNCIÓN A LA PSICOPATOLOGÍA
Segunda parte: La microestructura y el procesamiento de la información
THE BRAIN: FROM STRUCTURE AND FUNCTION TO PSYCHOPATOLOGY.
Second Part: Microestructure and the Processing of Information.
César Augusto Arango-Dávila* * Médico psiquiatra de la Pontificia Universidad Javeriana, magíster en Ciencias Básicas Médicas, candidato a Ph. D. en Neurociencias del Centro de Estudios Cerebrales, Facultad de Salud, Universidad del Valle.
Resumen
Las últimas dos décadas han sido de fundamental importancia para el estudio y entendimiento del sistema nervioso. Este progreso no sólo ha incluido el abordaje de fenómenos subyacentes a la masa cerebral de tipo neuroquímico, neurofisiológico, neuroanatómico, citoarquitectónico, imagenológico, etc., sino la relación de estos conocimientos con aspectos complejos de la conducta y el psiquismo.
La intención de recalcar con una visión moderna el alcance de las neurociencias en la comprensión del psiquismo humano ha de reflejarse necesariamente en una comprensión más amplia de la enfermedad mental, lo que permite una confluencia entre nuestra condición biológica, conductual, psíquica y social. En este artículo, basados en la organización macro y microestructural del sistema nervioso, se explican con detalle los fenómenos de procesamiento de la información en el cerebro y el resultado de las alteraciones de los sistemas de neurotransmisores en la génesis de las ideas delirantes, las experiencias de desrealización y despersonalización, las alucinaciones, las ilusiones, los trastornos del afecto y los trastornos de ansiedad.
Palabras clave: sistema nervioso central, cognición, afecto, neurobiología, psicobiología, columnas corticales, módulos corticales.
Abstract
The last two decades have been of fundamental importance in the study and understanding of the nervous system, this progress has not only addressed the neurochemical, neurophysiological, neuroanatomical, cytoarchitectonical, imageological phenomena underlying the cerebral mass but also the relationship of this knowledge with complex aspects of behavior and the psyche. The purpose of emphasizing a modern vision of the grasp of neuroscience must necessarily reflect a wider understanding of the reality of mental illness allowing a confluence between our biological, behavioral, psychic and social conditions.
Based on the macro and microstructural organization of the nervous system this article explains in detail the phenomena of processing of information in the brain and the result of alterations in the neurotransmitter systems in the genesis of delusions, derealization and depersonalization, hallucinations, illusions, mood disorders and anxiety disorders.
Key words: Central nervous system, cognition, affect, neurobiology, psychobiology, cortical columns; cortical modules.
Introducción
En el presente número los doctores Gabriel Arteaga y Hernán Pimienta, en su artículo "Sobre la organización columnar de la corteza cerebral", hacen una magistral exposición sobre la forma como se ha desentrañado la organización microestructural de la corteza cerebral. Se puede considerar que el descubrimiento importante es la existencia en la corteza cerebral de grupos de neuronas, mejor o peor definidos, que llegan hasta diez mil tal vez (enlazados mediante conexiones mutuas, lo que les confiere cierta existencia unitaria) y que presentan una gran confluencia sináptica que tiene un efecto inhibitorio sobre las columnas próximas. Si bien una columna puede tener una definición fisiológica muy clara, en sentido anatómico generalmente se imbrica o traslapa con otras columnas (1).
De esta complicada disposición citoarquitectónica de la corteza cerebral se desprende una organización funcional global que se manifiesta en dos órdenes microestructurales diferentes (2),(3). Un primer orden que corresponde a las conexiones neuronales de las láminas II, III, IV y V y, un segundo orden que se relaciona con el tipo de organización citoarquitectónica de la lámina I.
Primer orden: organización modular y convergencia
El primer orden radica en la compleja función localizada de dendritas, cuerpos neuronales y fibras aferentes, donde se pueden delimitar complejos estructurales y circuitos locales que caracterizan lo que se ha denominado columnas o módulos corticales. Éstos hacen parte de las láminas II, III, IV y V de la corteza y en ellos se concentra una importante actividad sináptica y metabólica. En estas láminas están situados el soma y las dendritas de las grandes células piramidales y es donde las fibras aferentes (provenientes de áreas subcorticales como los núcleos específicos del tálamo) ejercen su influencia sináptica principal, directa o indirectamente, a través del sistema de interneuronas (Figura 1).
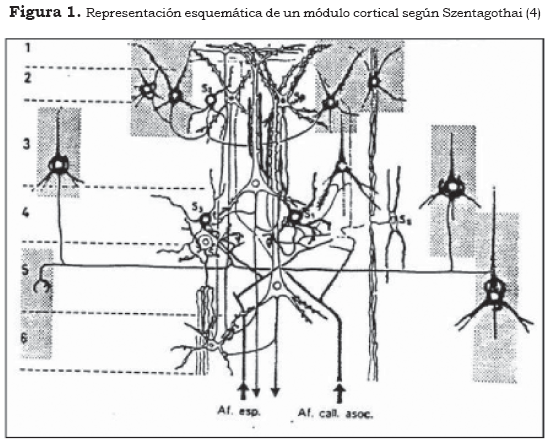
En la Figura 1, que es la representación esquemática de un módulo cortical según Szentagothai, se observan en blanco las neuronas glutamatérgicas excitadoras y en negro las neuronas gabérgicas inhibidoras. Se muestra, asimismo, la inhibición de las estructuras modulares adyacentes.
Hubel y Wiesel (5) han mostrado cómo en la corteza cerebral de gatos y monos la actividad de un módulo implica una respuesta global, en la cual se puede observar —gracias a imágenes autorradiográficas— que la columna se activa como conjunto neuronal, mientras que las columnas adyacentes se inhiben. Como veremos más adelante, se puede considerar que cada módulo activado corresponde a la confluencia de una multitud de sinapsis excitadoras e inhibidoras distribuidas de forma tridimensional que generan un paquete de información específico y simultáneo a través de frecuencias particulares de descarga de cada uno de sus axones eferentes, los cuales constituyen cuantos de información en forma de lo que denominaremos esquemas espaciotemporales. De esta manera se considera la función modular como organizaciones neuronales con una importante dosis de convergencia y síntesis, con capacidad de respuesta en códigos neurofisiológicos que representan el procesamiento de la información.
El procesamiento de información cortical requiere la existencia de varias constelaciones operativas estables (módulos), que controlan los procesos intracorticales a través de un mecanismo de sincronización neuronal, el cual se distribuye para producir una descarga cortical; por lo tanto, precisan de una amplia organización de convergencia interna (6). Este tipo de procesamiento es el sustrato de los procesos cognitivos.
Segundo orden: lámina I y divergencia
El segundo orden lo conforma una multitud de axones que corre tangencialmente a la superficie cortical en el espesor de la lámina I (prácticamente acelular) y que en su trayecto forma sinapsis sobrecruzadas con las dendritas apicales de las neuronas piramidales y en menor proporción sinapsis con otras células de la lámina II (neuronas estrelladas e interneuronas gabaérgicas). Algunos de estos axones que corren de forma paralela a la lámina I tienen una longitud de hasta cinco milímetros y se imbrican unos sobre otros y éstos sobre dendritas para formar un espeso e intrincado sistema de fibras (2),(3). La lámina I, así constituida, aparte de que tiene una fuerte dosis de divergencia, recibe tanto fibras de asociación (paleocorticales, arquicorticales y neocorticales), que son axones de la corteza límbica y de módulos variablemente remotos, incluso de la corteza contralateral (fibras comisurales), como los axones de los núcleos intralaminares (inespecíficos) del tálamo (Figura 2).
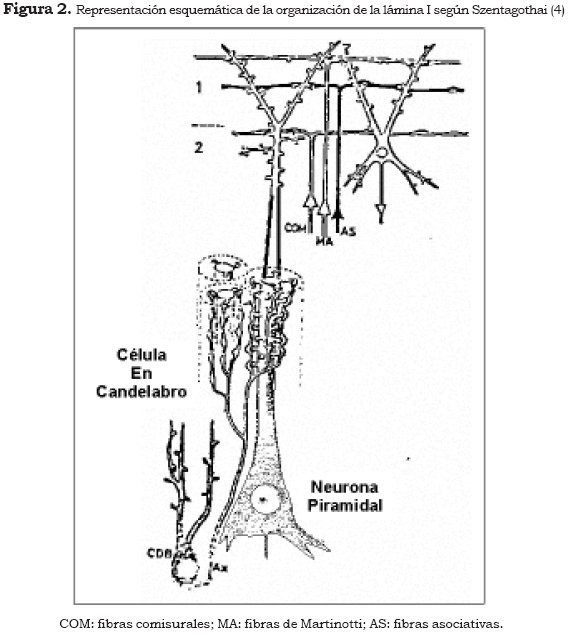
En la Figura 2 se observa la importante tendencia a la divergencia. Es importante señalar que en la corteza prefrontal dorsolateral confluyen en la lámina I fibras provenientes de la corteza prefrontal límbica y fibras de los núcleos inespecíficos del tálamo.
La lámina I conforma un espeso reticulado donde las aferencias ejercen una excitación sináptica extensa, divergente, pero global e inespecífica, que estimula o inhibe las porciones más distales de las dendritas apicales de las neuronas piramidales y las dendritas de otras neuronas glutamatérgicas (estrelladas) y gabérgicas (4). Como veremos más adelante, el tipo de procesamiento de información de la lámina I es el sustrato de los fenómenos motivacionales y afectivos.
Organización modular y procesamiento de información específica (cognitiva)
Aunque hemos definido los módulos corticales como unidades, no podemos descartar que en virtud del sobrelapamiento y del encabalgamiento que presentan unos sobre otros (1) puedan tener un funcionamiento múltiple integrado, pues de hecho se considera que su organización es de estas características. Sin embargo, el hecho de que se insinúen como 'unidades' nos permite identificar características subyacentes a la función cortical. Con fines explicativos procuraremos imaginar un módulo hipotético. Este módulo lo conforman cuatro células piramidales con sus respectivos axones encargados de representar la información eferente, y cada axón descarga impulsos a una determinada frecuencia así (Figura 3A):
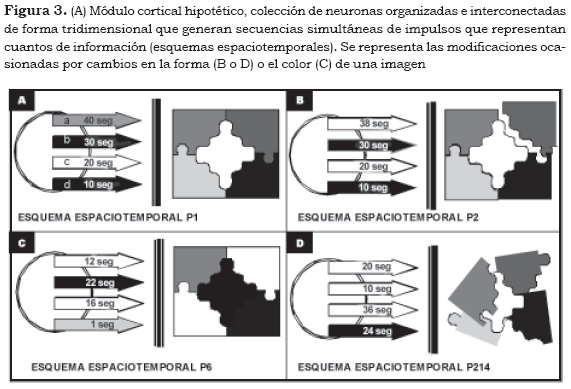
Axón a: 40 imp/s
Axón b: 30 imp/s
Axón c: 20 imp/s
Axón d: 10 imp/s
A esta colección específica de impulsos la denominaremos esquema espaciotemporal P1 (perceptivo 1), porque consideramos que esta información proviene de vías perceptivas, ya que si procediera de registros de memoria, la denominaríamos esquema espaciotemporal M1 (Mnésico 1). Para que el esquema espaciotemporal P1 pueda mantener la actividad de sus axones constante y estable ha debido ocurrir una serie de eventos sinápticos intramodulares tridimensionales inhibitorios-excitatorios con una definida organización temporal a corto, mediano o largo plazo en diferentes puntos del módulo. La reunión de estos eventos espaciales y temporales conforma lo que hemos denominado esquema espaciotemporal de información nerviosa, que se puede definir como paquetes o cuantos de información.
El esquema espaciotemporal P1, así conformado, se proyecta a través de las frecuencias específicas de sus axones a otros sectores de la corteza que reciben su simultaneidad y la integra al resto de esquemas espaciotemporales suministrados por otras estructuras modulares. Sin embargo, lo menos probable es que el esquema espaciotemporal P1 pueda mantenerse estable, pues constantemente recibe impulsos de otras áreas de la corteza y de estructuras subcorticales que ocasionan nuevas organizaciones excitatorias-inhibitorias tridimensionales, con nuevas coordenadas temporales, que se expresan en cambios de la frecuencia de descargas de sus axones.
Si continuamos con el ejemplo de un módulo hipotético, podría ocurrir que las nuevas frecuencias de descarga de los axones, como consecuencia de los cambios en el interior del módulo, sea (Figura 3B):
Axón a: 38 imp/s
Axón b: 30 imp/s
Axón c: 20 imp/s
Axón d: 10 imp/s
Observamos que el axón a ya no descarga con una frecuencia de 40 imp/s, ahora su frecuencia es de 38 imp/s. Este aparente breve cambio es suficiente para que ya no exista el esquema espaciotemporal P1 y que en su lugar se sitúe lo que denominaremos esquema espaciotemporal P2. El esquema espaciotemporal del complejo P2 comprende una información diferente al esquema del complejo P1 (véase el cambio en la imagen en la Figura 3B).
Con nuestro ejemplo hipotético de un módulo con cuatro axones eferentes que se activan de forma integrada en secuencias espaciotemporales, sería infinita la cantidad de esquemas espaciotemporales posibles, si consideramos que variaciones de impulsos por segundo en solo uno, de cada uno de sus axones, podría ya cambiar el esquema previo.
Nuestro ejemplo corresponde a una extraordinaria simplificación de lo que en realidad ocurre, pues un módulo como el descrito por Szentagothai, citado por Fuster (2), está constituido por diez mil neuronas con varios cientos de células piramidales y varios cientos de axones, cada uno en un determinado momento con una definida frecuencia de impulsos, que en su simultaneidad podría representar información de evidente complejidad y cambios infinitos. Se ha planteado que dentro de un módulo de estas características se organizan subgrupos de neuronas con funciones específicas a las cuales se les ha denominado minimódulos.
Existe una intensa interacción entre los módulos. La relación más clara consiste en una acción inhibitoria sobre los módulos inmediatamente adyacentes y en acciones excitadoras a través de sus axones de proyección. Dado que cada módulo posee unos cientos de células piramidales con axones que salen de éste para ir a otras partes, al estimular áreas remotas pueden ocasionar, como un efecto sumatorio, la activación de módulos subyacentes propios de estas áreas que así pasarán a activar otros sitios. Cada sitio de relevo logra una síntesis informativa antes de proyectarse a otro lugar.
Así, en el equilibrio y control dinámico de la corteza cerebral operante, existen todos los tipos de niveles posibles que podrían corresponder a una amplia gama de representaciones perceptivas o evocables.
La función modular ha sido estudiada por muchos investigadores, quienes han aportado importantes conceptos para el entendimiento, aunque sea parcial, de semejante complejidad. Podemos resumir los siguientes aspectos propios de la fisiología de los módulos corticales (véase Arteaga y Pimienta en este mismo número):
• Interacciones complejas de convergencia, inhibición frente a estimulación y retroalimentación frente a prealimentación.
• Inhibición intracortical.
• Respuestas sincronizadas en secuencias temporales.
• Respuestas variables con latencias mínimas y largas que expresan plasticidad modular.
• Convergencia espaciotemporal tópica y convergencia espaciotemporal confluente.
• Superposición de mapas y superposición de esquemas espaciotemporales.
• Modulación de información aferente y aferente.
Es necesario confrontar lo explicado hasta aquí con algunos planteamientos hechos por el neurobiólogo J. P. Changeux (7) al referirse al mismo aspecto. Este autor propone la existencia de un mecanismo neuronal combinatorio de naturaleza epigenética, es decir, un mecanismo en el que no media la modificación estructural de la neurona. Por lo tanto, sería no el resultado de un proceso de neuronas individuales, sino el de un conjunto muy organizado de células nerviosas (módulos). Su origen es la topología de la red de conexiones que se establecen entre neuronas a lo largo del desarrollo y se "inscribe en la multiplicidad de figuras geométricas que se dibujan transitoriamente en las tres dimensiones del espacio en el momento de la formación de esa red" (esquemas espaciotemporales).
Lo que aquí hemos denominado módulo perceptivo, para Changeux es el percepto primario; por otro lado, lo que hemos llamado módulo mnésico, Changeux lo denomina imagen. Ambos elementos actúan como origen del objeto mental. El objeto mental se identifica en el estado físico creado por la entrada en actividad (eléctrica o química), correlativa y transitoria, de una amplia población o conjunto de neuronas distribuidas en varias áreas corticales definidas. Este conjunto, que se describe matemáticamente por medio de un grafo, es discriminado, cerrado y autónomo, pero no homogéneo. Changeux hace hincapié en esta característica de la actividad mental y refiere que el objeto mental tiene una organización que es a la vez local y deslocalizada.
El percepto primario es un objeto mental cuyo grafo y actividad están determinados por la interacción con el mundo exterior (vía perceptiva). El grafo neurónico (módulo perceptivo con su respectivo esquema espaciotemporal) asociado con éste, debe su existencia al hecho de que está en conexión directa con el objeto exterior. La imagen es un objeto de memoria autónoma y fugaz, cuya evocación no requiere una interacción directa con el entorno (vía mnésica). Su autonomía no se concibe si no existe un acoplamiento de neuronas del grafo (módulo mnésico con su respectivo esquema espaciotemporal) estable en el tiempo y que preexiste a su evocación.
Para comprender la cantidad o el cuanto de información que puede contener un módulo debemos remitirnos a los fenómenos experienciales que resultan de la estimulación directa o disrítmica de la corteza temporal. Se ha comprobado que en este lóbulo se encuentra en gran medida el sustrato de este tipo de respuesta (8). Gloor (9) señala que los fenómenos experienciales epilépticos o artificiales por estimulación eléctrica del lóbulo temporal son la expresión positiva de la función de dicho lóbulo, cuando la alteración no refleja una interferencia ictal o parálisis de esa función. Postula, además, que en la base de la experiencia de evocación hay una matriz con patrones específicos fundamentado en grupos de neuronas (módulos). Tales agrupaciones han codificado la información perceptual, mnésica y afectiva (esquema espaciotemporal) y han logrado la representación de una experiencia específica que se fortalece, en el caso de la epilepsia, por las repetidas descargas.
La expresión de estos módulos activados incluye una gama importante, pero específica de fenómenos mentales, vivenciales y de evocación, como trozos de piezas musicales, situaciones cotidianas (como hacer de comer o ver televisión), fragmentos de poemas o libros, recuerdos específicos de la infancia, etc.
Lámina I y procesamiento de información inespecífica (afectiva)
Basados en lo expuesto, podemos pensar que hay un significado esencial en todas las interacciones neuronales, que finalmente se expresan como patrones espaciotemporales en miríadas de puntos que se transmiten de una a otra parte de la corteza. En la lámina I los procesos inhibitorios y excitatorios son ampliamente dispersos, no se observan las profusas interacciones sinápticas de las láminas más profundas. Así, se considera que las sinapsis de la lámina I ejercen su actividad de una manera más general e inespecífica. Hay en estas láminas un importante flujo divergente y resulta interesante que en la corteza prefrontal dorsolateral este flujo provenga de fibras de la corteza límbica, inespecíficas del tálamo y aferentes callósicas y de asociación.
La función global de la lámina I es evidente: ésta no integra por contigüidad anatómica, sino por comunicación funcional paso a paso. Hay fuertes motivos para pensar que el cerebro de relación comprende una gran parte del hemisferio dominante, particularmente las áreas del lóbulo prefrontal, y aquí es donde la función integradora de la lámina I presenta una importante expresión: integra la información suministrada por el tálamo, por el sistema límbico y otras áreas corticales. Las miríadas de esquemas espaciotemporales, junto a la capacidad moduladora de la lámina I, podría ser el sustrato de la experiencia autoconsciente con los componentes cognitivo y afectivo.
Organización modular y lámina I (cognición y afecto)
La corteza prefrontal presenta dos importantes sistemas aferentes. Por un lado está la información suministrada por los órganos de los sentidos, que después de múltiples procesos de segregación e integración finalmente se proyectan a la corteza prefrontal en forma de esquemas espaciotemporales, que aquí hemos denominado esquemas espaciotemporales P (perceptuales) o perceptos primarios, según Changeux (7). Por otro se encuentran un tipo de esquema suministrado por módulos que han registrado información reciente o remota y que la han guardado para presentarla a disposición de la corteza prefrontal, que hemos denominado complejos modulares M (mnésicos) o imágenes, según Changeux. Estos complejos corresponden a los bancos de memoria reciente y remota, localizados principalmente en los lóbulos temporales en los cuales el hipocampo cumple una especial tarea.
Nuevamente tomaremos con fines explicativos un módulo hipotético, con cuatro células piramidales y cuatro axones que emiten definidas frecuencias de impulsos así:
Axón a: 20 imp/s
Axón b: 40 imp/s
Axón c: 30 imp/s
Axón d: 16 imp/s
A este paquete de información lo denominaremos esquema espaciotemporal P1 (perceptual). Cuando este complejo perceptual remite su esquema espaciotemporal a la corteza prefrontal, inmediata y automáticamente se relaciona con los complejos similares entre los complejos modulares M, mecanismo posibilitado por el sistema comparador. Pueden ocurrir así dos cosas:
• En caso de que haya un registro previo y se encuentre un esquema espaciotemporal M igual, la información suministrada por el esquema pasa inadvertida por la corteza prefrontal, pues ya se encuentra registrada.
• En caso de que el complejo modular P1 no esté registrado, el sistema comparador 'pone en alerta' a la corteza prefrontal para que ésta procese la información o la incorpore en la corteza temporal como complejo modular M1 (Figura 4).
Es importante señalar aquí que este sistema comparador (10), cuyos componentes están relacionados con las estructuras límbicas, realmente lo que hace es activar la corteza prefrontal gracias a su importante flujo de proyecciones hacia la lámina I, y 'ponerla en alerta' consiste en incrementar el grado de activación de los módulos subyacentes a la lámina I —recuérdese que en las láminas más superficiales de la corteza se encuentra no sólo la aferencia proveniente de la corteza límbica, sino una importante inervación noradrenérgica y serotoninérgica (11)—.
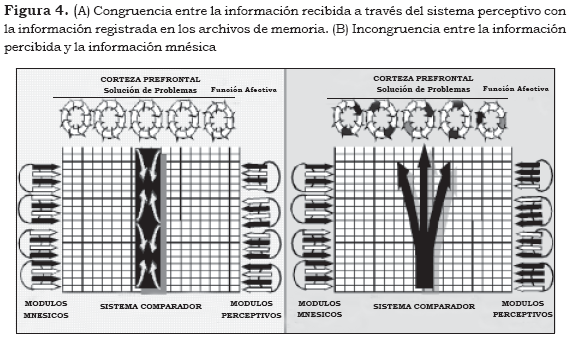
En la Figura 4A se observa la congruencia entre la información recibida a través del sistema perceptivo con la información registrada en los archivos de memoria, por lo tanto, el sistema comparador se encuentra inactivo y no estimula a la corteza prefrontal. En la Figura 4B se observa la incongruencia entre la información percibida y la información mnésica (flechas blancas y negras en espejo), se activa el sistema comparador, el cual obliga a la corteza prefrontal a buscar la congruencia entre ambos sistemas (solución de problemas).
Se impone examinar un poco cómo puede la corteza prefrontal dorsolateral ser el asiento de la experiencia autoconsciente. En primer lugar, muchos investigadores han señalado el carácter unitario en la experiencia autoconsciente (3),(7), (12). En cada instante se da una concentración en uno o en otro aspecto del funcionamiento cerebral. Ya sea en lo que hemos denominado esquemas espaciotemporales P o esquemas espaciotemporales M, esta secuencia constituye el fenómeno conocido como atención. El carácter cíclico de este proceso, merced al cual determinados contenidos de la actividad (complejos modulares M o P) adquieren valor dominante o lo pierden, se corresponde con lo que Luria ha denominado oscilaciones de la atención (13). Este autor diferencia la atención voluntaria de la reacción de orientación elemental, y considera que la atención voluntaria no es en principio biológico, sino que supone la existencia de complejos factores estimulantes formados a través de la historia del individuo.
Aquí es donde es necesario señalar el carácter automático del sistema comparador, el cual no es consciente aunque desencadene respuestas complejas. Es la corteza prefrontal dorsolateral la que cuenta con la posibilidad de activarse gracias a este sistema comparador y hacer un manejo 'voluntario', gracias a su capacidad de búsqueda y confrontación, atributo dado por su posición central como estructura solucionadora de problemas, la cual debe encontrar la congruencia entre los esquemas espaciotemporales P y los esquemas espaciotemporales M.
En segundo lugar, hay una relación continua entre los sucesos de las áreas prefrontales y los registros almacenados en los lóbulos temporales. La motivación cognoscitiva surge de la asimetría entre estos dos tipos de información; por ello cuando los complejos modulares perceptivos no se corresponden con los complejos modulares mnésicos, entonces se movilizan procesos para integrar la nueva información o desecharla.
Si hemos considerado la maquinaria neuronal como un complejo de estructuras radiantes y receptoras, la unidad experimentada como autoconciencia es resultante de la integración de todas las modalidades de información. Tal como se ha descrito, las áreas sensoriales primarias se proyectan a las secundarias y éstas a las terciarias, etc. En estas etapas ulteriores las diferentes modalidades se proyectan a áreas comunes, las áreas polimodales. En dichas áreas, la información más diversa y de amplio espectro se procesa en los componentes unitarios que hemos denominado esquemas espaciotemporales perceptivos, los cuales integran la información.
Pero no toda esta información pasa inadvertida (compatibilidad complejos modulares P y M). La corteza prefrontal procesa la información en la cual los complejos P y M no fueron compatibles y se pone en el trabajo de seleccionar módulos, ya sea perceptuales o mnésicos, en un intento por relacionarlos con la información incomprendida. Así, la corteza prefrontal desempeña un papel activo de búsqueda y sondeo, que desvía y moldea la información que ha sido influenciada por los sucesos mnésicos y perceptivos, e interviene a su vez sobre ellos.
Esta importante localización jerárquica de la corteza prefrontal está magnificada por su capacidad para activar módulos remotos en busca de información, además por el hecho de poder explorar el mundo exterior, por su especial influencia sobre los sucesos motores voluntarios. Esta influencia incluye una sucesión neuronal progresiva hacia partes posteriores de la corteza frontal, por lo que se observa un paulatino cambio en que las células nerviosas presentan menos conexiones con estímulos sensoriales y más conexiones con la función motora, que se concretan finalmente los aspectos operativos de la acción (14).
Mención aparte debe hacerse de la percepción. El acto perceptivo como tal constituye una forma superior de conocimiento a través de la cual aprehendemos el mundo circundante y nuestra propia corporalidad. Por lo tanto, es un proceso activo en cuanto su puesta en marcha no es absolutamente casual, sino que, por el contrario, supone una actitud previa por parte del sujeto, que de esta forma tiene cierta capacidad de elección del objeto percibido en función de su situación psíquica, fisiológica o de ambas. La particularidad del objeto percibido modifica, asimismo, la situación del sujeto perceptor, lo cual corrobora también la categoría de proceso dinámico y activo (10),(12).
Aunque ya a nivel de receptor de un órgano sensorial determinado existen ciertas posibilidades de control o filtraje cuantitativo y cualitativo de la información, el verdadero proceso activo se produce, tal como hemos dicho, a nivel superior. En efecto, una vez que el mensaje alcanza estas zonas elevadas, debe ser reconocido y, por así decirlo, calibrado. Ahora bien, esta labor, que comprende los procesos de selección y reorganización aludidos, precisa de toda la información acumulada en la memoria de mediano y largo plazo, la cual presta al sujeto en su situación presente toda su experiencia histórico-biográfica, por lo que la percepción se convierte en un acto personal intrínsecamente subjetivo.
Si retomamos todo lo expuesto, encontramos que la acción conjunta de lo que hemos denominado organización modular y procesamiento de información específica (cognitiva) y lámina I y procesamiento de información inespeíifica (afectiva), en la zona de la corteza prefrontal, constituye la base fundamental donde se centran los procesos de pensamiento. La reunión de estas dos funciones permite la movilización y operación de la memoria, la atención, la percepción y el afecto, todo lo cual, en los términos que lo hemos expuesto, podría posibilitar la experiencia autoconsciente.
El pensamiento es la estructura del psiquismo humano que enlaza percepciones, representaciones, evocaciones y afectos para encaminarlos a una finalidad determinada, en general, con la intención de solucionar un problema (asimetría entre complejos modulares mnésicos). Así, el problema mismo, el obstáculo, es la motivación cognitiva del proceso.
Las láminas superficiales e intermedias de la corteza prefrontal contienen la mayor densidad de receptores, cuyos neurtransmisores clásicamente se han asociado con el afecto (serotonina y norepinefrina). Esta organización de las láminas superficiales se opone a la de las láminas más profundas, donde se encuentra la mayor densidad de receptores D2 (11). Se plantea la hipótesis de que en la lámina I existe, a más de la capacidad integradora fundamental para los procesos afectivos y del pensamiento, una segunda capacidad de crucial importancia en el psiquismo: poder contener el afecto en su expresión superior, es decir, ligado a la autoconciencia.
La lámina I de la corteza prefrontal posibilita la integración de los componentes cognitivo y afectivo, que son marcadamente manifiestos en la vida psíquica, y se encuentra así un sustrato biológico concomitante al afecto y al pensamiento de la corteza prefrontal (Cuadro 1). En el caso de los fenómenos afectivos, se deducen circunstancias que indican que éstos ocurren de una manera menos específica y más general. En su forma más simplificada se puede considerar el afecto como un aumento o una disminución de la conductancia de las fibras nerviosas de la lámina superficial de la corteza así:
• Un aumento en la conductancia ocasiona un incremento en la velocidad y cohesión de los procesos integrativos cognitivos.
• Una disminución en la conductancia causa una lentificación del flujo integrativo.
La conductancia determina así la proporción cohesiva, y ésta con la integración de los esquemas espaciotemporales se correspondería al pensamiento con su tono afectivo.
Se convierte entonces el afecto en matriz sobre la que los procesos cognitivos instalan su función y la efectividad de estos últimos depende del estado del primero.
Los procesos psíquicos siempre están acompañados de un colorido que los matiza, el cual le da al pensamiento su característica subjetiva a cada momento y contribuye a orientar la conducta. Si resaltamos los siguientes tres aspectos del afecto, podremos entender su posición central referida en los párrafos anteriores (15):
• La trascendencia, que es la difusión que ejerce el afecto sobre todas las dimensiones del psiquismo.
• La polaridad, que se refiere a la variedad cualitativa en que se imponen los afectos, por ejemplo, depresión-manía, huida-ataque.
• La comunicabilidad, que se refiere a la influencia y participación del afecto en el vínculo entre el individuo sus congéneres y el medio.
Se observa que estos tres atributos del afecto se relacionan con una respuesta psíquica global, lo que le confiere, siguiendo nuestro orden de ideas, el carácter inespecífico. El afecto no consiste en la formación de esquemas espaciotemporales específicos, como ocurre en los procesos cognitivos, sino en un incremento o disminución de la conductancia en la lámina I, lo cual se refleja en un incremento o disminución de la velocidad de formación de los esquemas espaciotemporales que son generados por los módulos corticales que subyacen a esta lámina I. Las psicologías cognitiva y psicodinámica muestran que el afecto y la cognición no ocurren aislados, sino que están fuertemente combinados como lo exponen Freud (16), Rapaport (17), Piaget (18), entre otros. En el Cuadro 1 se señalan las razones para considerar que las láminas más superficiales de la corteza prefrontal integran funciones cognitivas y afectivas.

Alteración funcional de los módulos corticales como fundamento de la psicopatología de los procesos mentales superiores
A la complicada citoarquitectura de los módulos corticales se sobreponen diferentes sistemas de neurotransmisores que, en últimas, son los responsables de la fisiología específica de las neuronas. En el interior del módulo, los neurotransmisores modelan la información que habrá de expresarse a través de lo que hemos llamado esquemas espaciotemporales, es decir, organizaciones definidas de impulsos, cada una con una frecuencia determinada que fluye entre los cientos de axones del módulo de una manera global y simultánea.
Esta información es integrada en la corteza prefrontal donde hace parte del pensamiento. La información de los complejos modulares perceptivos es confrontada con los complejos modulares mnésicos con el fin de ser reconocida. Si no es reconocida, se movilizan procesos encaminados a incorporar, asociar o desechar la nueva información presentada por este nuevo esquema espaciotemporal. Este proceso activo de incorporación, asociación o exclusión requiere la participación integrada de la corteza temporal (especialmente el hipocampo y el sistema comparador) y de la corteza prefrontal, las cuales permiten que la nueva representación se asocie con otras representaciones de características similares y se incorpore ligada a circunstancias históricobiográficas o cognitivas del individuo registrada en los complejos modulares mnésicos.
El lóbulo temporal actúa cumple una función muy activa así como coadyuvante de la corteza prefrontal: le presenta, por disposición de ésta, esquemas que se aproximan cada vez más al requerido. En el caso de una alteración de la fisiología modular por un defecto funcional en uno o varios de sus neurotransmisores, las consecuencias repercutirán a través de todos los procesos implicados en el sistema, y el resultado tendrá significativas peculiaridades.
Fisiopatología de la psicosis
Si los complejos modulares perceptivos o los complejos modulares mnésicos presentan una alteración inducida o espontánea en uno o varios neurotransmisores de su interior, la resultante de este defecto es la generación de esquemas espaciotemporales que no se relacionan de una manera definida con lo percibido. Sin embargo, estos esquemas espaciotemporales no congruentes con las regularidades almacenadas, son automáticamente reconocidos por el sistema comparador, el cual activa la corteza prefrontal, la cual debe hacer congruentes los módulos perceptivos con los módulos mnésicos por su papel biológico en la solución de problemas.
Como la función integradora de la corteza prefrontal se encuentra indemne, recibe los esquemas espaciotemporales anómalos y los confronta con los esquemas de los complejos modulares mnésicos o perceptivos. En estas circunstancias, como es de esperarse, no localiza los correspondientes, pues no existen. La corteza prefrontal 'insiste' en la búsqueda (debido a la activación irreducible que le confiere el sistema comparador, que detecta permanentemente la incongruencia), necesita encontrar explicación a esa información, pero la corteza temporal y sus módulos M y los sistemas perceptivos con sus módulos P, por más que busque en sus registros, no encuentran esquemas comparativos.
La alteración de los complejos modulares perceptivos con sus correspondientes esquemas espaciotemporales aberrantes, irreconocibles, genera en la síntesis final de la corteza prefrontal la vivencia de "algo diferente", como si el ambiente fuera distinto, con una alteración sutil que lo envuelve todo, pero que no puede ser identificada, algo incierto. La posibilidad de percibir el ambiente como "algo diferente" está en directa relación con la incompatibilidad de los complejos modulares perceptivos con los mnésicos y con una función prefrontal íntegra.
En la Figura 5 aparece un defecto en la particularidad bioquímica de los módulos perceptivos o mnésicos, que genera esquemas espaciotemporales aberrantes, irreconocibles, con los elementos histórico-biográficos o la percepción del individuo. Se observa que el sistema comparador activa la corteza prefrontal, la cual está obligada a solucionar el 'problema'. En este estado la vivencia es de desrealización, si el defecto es más pronunciado en los módulos P, o de despersonalización, si el defecto es más pronunciado en los módulos M. La perplejidad es una manifestación semiológica frecuente en este estado.
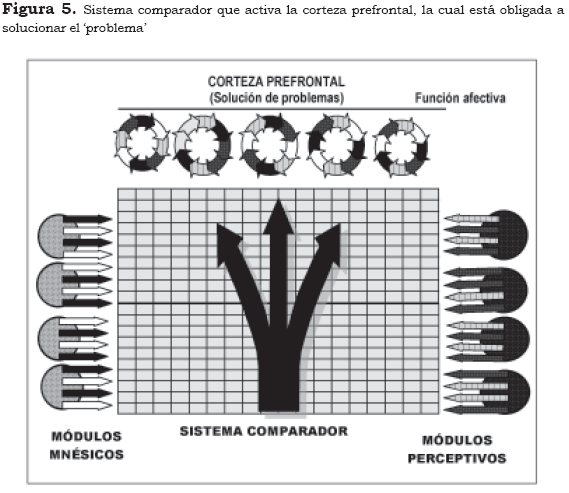
Así surge lo que semiológicamente se ha denominado desrealización. La corteza prefrontal, al no poder aclarar lo indefinido, se suspende en un estado de indeterminación que cada vez se torna menos tolerable y la impulsa a persistir en la búsqueda. El individuo se observa desazonado, inquieto, alarmado, inhibido por algo que ni siquiera puede definir con su lenguaje. Observa detenidamente los objetos, mira a su alrededor y su sensación de extrañeza es cada vez más paralizante. Su función prefrontal está íntegra, puede inhibir parcialmente los complejos modulares perceptivos y destacar los complejos mnésicos, pero tampoco habrá respuesta, más bien, por la invasión de lo perceptivo y su confrontación fallida con lo mnésico, surgirá la experiencia de despersonalización: "algo cambia afuera o algo cambia dentro de mí".
Si este proceso continúa, si los cambios de los esquemas espaciotemporales son tan marcados que no pueden ser procesados de ninguna manera por la función laminar de la corteza prefrontal, el paciente queda sumido en un estado de perplejidad paralizante, reflejo de la desestructuración del campo de su conciencia. No ocurre lo mismo si las alteraciones modulares no son tan graves o si la función prefrontal puede contener el proceso sin paralizarse. Presionada por la activación del sistema comparador, la corteza prefrontal, acosada por la necesidad de elaborar la información presentada por los complejos modulares perceptivos o mnésicos anómalos, crea una teoría, pero no una teoría cualquiera, sino aquélla en que encuentre compatibilidad entre los complejos modulares perceptivos y los complejos modulares mnésicos, de tal manera que pueda solucionar el 'problema'.
En este proceso de crear una teoría que haga compatible el sistema perceptivo y el sistema mnésico el individuo empieza a "entender", estructura una escala de significaciones de lo perceptivo aplicable a lo mnésico o de lo mnésico aplicable a lo perceptivo. Todo lo percibido lo carga de nuevas significaciones y lo confronta con los complejos modulares mnésicos, o interpreta el entorno con relación a sus registros mnésicos. En este momento, cuando la percepción adquiere un significado determinado y propio que de alguna manera se hace compatible con los complejos modulares M o P, nos encontramos ante el fenómeno de la interpretación delirante. Así, se centra la actividad psíquica en reestructurar lo percibido, o el mundo del paciente se interioriza cada ves más y de igual forma cada vez mas va siendo incompresible para sus congéneres.
El proceso de cargar de significaciones lo percibido y de crear una teoría es indicio de un adecuado desempeño de la función prefrontal, que recurre a todos los registros al macenados en otros sitios de la corteza (como la corteza temporal) y articula multitud de elementos en un sistema interno. De tal forma, toda la personalidad queda limitada al delirio, el cual se constituye en el eje de la vida del paciente y marca una ruptura histórico-biográfica entre su pasado y su situación actual.
Por provenir toda esta cascada de acontecimientos de una alteración en la forma de percibirse el mundo externo, la idea generada adquiere una certeza subjetiva y no existen dudas sobre su veracidad, es decir, la idea se torna irreductible. Al tratar de convencer a la persona de lo erróneo de su razonamiento, lo que se logra es motivarlo aún más a encontrar justificaciones, aspecto que muestra el carácter de irreversibilidad de la ideación delirante. El individuo no puede dejar su delirio, pues de lo contrario sería entregarse nuevamente a la incertidumbre. La función de la corteza prefrontal es solucionar problemas y en este caso lo logró, aunque sumiendo a la persona en un seudosistema lógico estructurado y sistematizado típico de la alienación delirante. El tipo de delirio dependerá de lo que tenga previamente registrado el paciente y de su capacidad habitual de asociación, por ello es viejo el concepto de que "el paciente delira con lo que tiene", pues su antecedente histórico-biográfico es componente básico de la estructuración.
Otros casos de afectación del campo de la conciencia se dan cuando a la alteración de los complejos modulares perceptivos se asocian defectos de iguales características en los complejos modulares mnésicos.
La función laminar, que puede estar parcial o totalmente preservada, continúa integrando, pero la compatibilidad de los complejos es viciada, tangencial e impropiamente, ocasionando la disgregación del pensamiento. Las asociaciones se desligan de la experiencia, son influenciadas de forma anómala por los estímulos externos y adquieren el característico matiz de autonomía propio del individuo psicótico. Si además de la alteración en los dos tipos de complejos modulares se instala una franca disfunción prefrontal, las consecuencias sobre los procesos de pensamiento se tornan más comprometedoras.
En la Figura 6 se muestra un defecto en la particularidad bioquímica de los módulos perceptivos, que genera esquemas espaciotemporales aberrantes, irreconocibles, con los elementos histórico-biográficos del individuo. La corteza prefrontal, en su función de solucionar problemas, carga de significados nuevos lo percibido y su propia historia, de esta manera se logra la compatibilidad entre los esquemas espaciotemporales perceptivos y mnésicos. El sistema comparador deja de activar a la corteza prefrontal, pero el individuo queda sumido en una estructutura delirante.
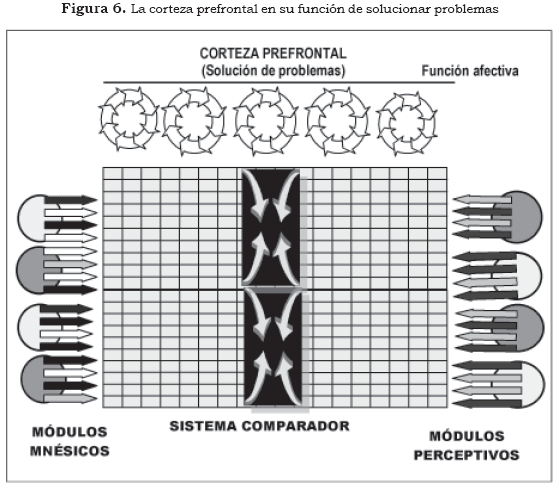
En la idea delirante un problema bioquímico se soluciona con una teoría psicológica.
En resumen, podemos decir que la alteración de los complejos modulares perceptivos desencadena un proceso consistente en cargar de significaciones lo percibido. La corteza prefrontal recurre a la información almacenada en otros sitios y articula multitud de elementos en su sistema, y en este proceso la corteza temporal adopta un papel activo referido a buscar todo aquello que tiende a estructurar la función prefrontal y su papel de solucionar problemas. En la idea delirante, un problema bioquímico se ha solucionado con una teoría psicológica.
La influencia que tiene el sistema comparador en la corteza prefrontal, y ésta sobre la corteza temporal, hace que se active de tal forma que en determinadas circunstancias le presente esquemas espaciotemporales complejos de registros mnésicos elaborados compatibles con su creencia. Cuando estos esquemas adquieren gran intensidad, nos encontramos frente a una alucinación, fenómeno experiencial en que el individuo percibe sin estímulo real, aunque la realidad debe estar fundamentada en la percepción. En la alucinación esta 'realidad' se transfiere a los registros amnésicos, para reconfirmar aún más la teoría generada por la función solucionadora de problemas de la corteza prefrontal. En otros casos, la interpretación que hace la corteza prefrontal de una percepción real se distorsiona en relación con la teoría elaborada y se manifiesta el fenómeno semiológico de la ilusión. Siempre el tipo de ilusión y alucinación constituida es coherente con el entramado de la idea delirante como si ésta se corroborara.
La fisiopatología de la ilusión y la alucinación, en términos generales, es similar a la fisiopatología de la idea delirante. Henry Ey (19) considera que toda enfermedad mental se caracteriza por la aparición de síntomas de déficit, que son la expresión de una desestructuración de la conciencia en diferentes campos. La disolución de un determinado nivel conduce a la liberación de otros niveles inferiores que se aparecen como una nueva reorganización de la vida psíquica. Esta dinámica implica la necesidad de ligar el trastorno mental, no tan sólo a un proceso de desorganización, sino también a una nueva organización. Para Ey la alucinación constituye un trastorno creencial, no sensorial, secundario a las ideas delirantes de las que es una prolongación.
Fisiopatología de los tras tornos del afecto
Como hemos visto en apartados anteriores, la corteza prefrontal dorsolateral es un sector con importante integración psíquica, donde confluyen en un estado superior el pensamiento y el afecto. Vimos también que el mecanismo fisiológico de los fenómenos afectivos son menos complejos y más generales basados en la conductancia aumentada o disminuida de la lámina I de la corteza cerebral, por lo que el afecto se constituye en la matriz sobre la que los procesos cognitivos instalan su función, pues éstos son producto de la integración ocurrida en el mismo sustrato.
Cuando hay un cambio subcortical o límbico que afecta la fisiología del grupo superficial de receptores de la corteza prefrontal, la resultante es un defecto cuantitativo de la fisología de la lámina I de la corteza prefrontal, que en su expresión más gruesa repercutirá en un aumento o disminución en la velocidad de procesamiento de la información.
En el caso de un aumento en la conductancia de la lámina I, los módulos subyacentes incrementan su actividad cualitativa y las relaciones de la corteza prefrontal con otras áreas se aceleran, hecho que clínicamente se manifiesta con un aumento cuantitativo de los procesos psíquicos. Aparece un desarrollo exageradamente rápido del proceso racional y se verbalizan un gran número de representaciones evocables por unidad de tiempo con una disminución en el período de latencia entre una pregunta y una respuesta o entre un problema y su solución. Cuando la conductancia laminar continúa incrementándose, se exagera la aceleración y se instalan el pensamiento ideofugal con taquipsiquia y fuga de ideas. En este caso, se aumenta la velocidad del mecanismo de la asociación de ideas con una gran susceptibilidad del pensamiento para desviarse de su curso central, fenómeno denominado distraibilidad. Se relacionan las percepciones y conceptos más próximos en el tiempo, pero el conjunto carece de sentido, de unidad significativa, por lo tanto, hay un desorden y falta aparente de finalidad completiva de las operaciones intelectuales.
Lo llamativo de todo esto es que el fenómeno de incremento de la actividad psíquica se asocia frecuentemente con la experiencia subjetiva de alegría. Los acontecimientos comunes de la vida del sujeto se experimentan entonces con un gran optimismo, sensación de capacidad y energía que generan un talante de omnipotencia típico de los estados de manía e hipomanía.
Cuando se manifiesta plenamente el cuadro, se observa una relación alienada con la realidad, que le confiere el carácter de psicótico. La corteza prefrontal, embarazada por la invasión afectiva inexplicable, queda ligada a la necesidad de generar una teoría que haga compatible el fenómeno afectivo con un contenido cognitivo, y de nuevo la corteza prefrontal cumple su papel de solucionar problemas. Recurre entonces a los complejos modulares mnésicos, los cuales buscan los registros más compatibles y los presentan como componentes del estado afectivo. De igual forma procede con las circunstancias del ambiente, de donde toma elementos que automáticamente se fusionan con su estado afectivo. De todo esto resulta la ideación delirante megalomaniaca en sus diferentes presentaciones clínicas, que dependen de los antecedentes históricobiográficos del individuo.
Cuando la conductancia de la lámina I en vez de acelerarse se lentifica, se observa claramente el desaceleramiento de los procesos mentales. La construcción prefrontal se liga a los contenidos tristes, lo cual explica la circunstancia vital con abatimiento, desánimo y aflicción, fenómeno definido como depresión (Cuadro 2).

Piaget, en su libro Inteligencia y afectividad, considera al afecto no únicamente como el motor de los elementos cognitivos, sino como su 'pegante' o tejido conectivo. Kovacs y Beck (20) explican cómo cuando surge uno u otro afecto, especialmente si el estímulo desencadenante no es evidente, se produce en el individuo una elaboración cognitiva para explicar qué le está ocurriendo. El sujeto trata de organizar lo que siente explicando así el fenómeno sobre la base de normativas contextuales o desde su propia racionalidad. Este factor cognitivo mantiene o incrementa el estado afectivo, que a su vez se implica en el primero.
Fisiopatología de los trastornos de ansiedad
Las manifestaciones clínicas de la ansiedad se relacionan con la afectación de las dos funciones corticales descritas. Un individuo ansioso está hipervigilante, inquieto y tenso; la concentración y la organización perceptiva están limitadas, por lo tanto, el pensamiento se observa atropellado y poco determinativo hasta alcanzar en algunos casos experiencias de despersonalización y desrealización (21) o sensación de pérdida del control (ataque de pánico).
El núcleo de toda esta sintomatología superior es el resultado de una activación súbita del sistema comparador (Figura 7), el cual tiene importantes componentes estructurales que provienen del sistema límbico y de núcleos subcorticales con repercusiones sobre la corteza prefrontal como los núcleos del Rafé (serotoninérgicos) y el locus coeruleus (noradrenérgico).
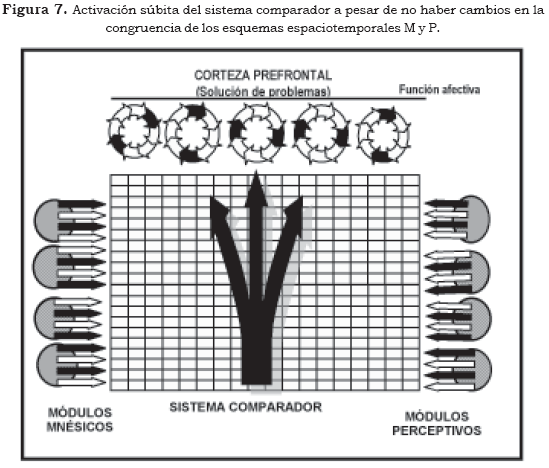
La corteza prefrontal moviliza esquemas espaciotemporales en busca de una congruencia para su dramática experiencia afectiva. Surge la impresión de inseguridad y temor la sensación de que "va a pasar algo" o se "va a perder el control". Lo característico de la ansiedad es su similitud fenomenológica con el miedo, en el cual se observa una función activadora, de forma que en circunstancias normales —ante un peligro o amenaza— se moviliza una tensión que tiene como función estimular la capacidad de respuesta del individuo tanto afectiva como cognitiva.
Gorman (22) considera que la evitación fóbica es secundaria al ataque de pánico o la ansiedad anticipatoria y que es resultado de una elaboración explicativa en la corteza prefrontal. La fobia no ocasiona la ansiedad o el pánico, sino que estos fenómenos no explicables generan una teoría en la zona prefrontal, lo que se aproxima a lo que ya se ha explicado.
Es necesario hacer hincapié en los cambios de los módulos en la corteza prefrontal, que generan diferentes manifestaciones clínicas. La ideación delirante y las alucinaciones son el resultado de alteraciones intrínsecas del módulo perceptivo o mnésico, que desencadena todos los procesos reorganizativos explicados con detalle anteriormente. El pánico y la ansiedad son cambios en el sector de la lámina I y de los módulos, pero estos cambios no son intrínsecos, son el resultado de un influjo de otras estructuras como el locus coeruleus y el sistema límbico. A pesar de estas diferencias, hay similitudes importantes en la expresión clínica de los dos sucesos, pues su estructuración fundamental se basa en los mismos sistemas:
• La desrealización y despersonalización son fenómenos presentes tanto en la ansiedad como en los estados psicóticos, sin embargo, en la ansiedad este estado es transitorio (en cuanto está disparado el sistema comparador), mientras que en la idea delirante es el resultado de un cambio neuroquímico intramodular.
• La idea delirante y la idea fóbica se generan en la misma función integradora y solucionadota de problemas la corteza prefrontal, sin embargo, la idea delirante es una convicción irreductible (cambios intramodulares continuos), mientras que la idea fóbica es susceptible de autocrítica y se considera anormal por el mismo individuo.
• Las conductas motoras asociadas con la ansiedad, como la psicosis, reflejan algún grado de similitud. Algunos estados se describen incluso como ansiedad psicótica.
• En todos los casos es más evidente la alteración cognitiva en los cambios intrínsecos de los módulos (ideas delirantes) y la alteración afectiva en los cambios no intrínsecos (idea fóbica).
Si bien es obvio que los estímulos ambientales y culturales cuentan en la expresión sintomática de un paciente, algunos aspectos del comportamiento y su sorprendente uniformidad en los diferentes individuos sugieren una programación biológica, como si algunas conductas estuvieran previamente imbricadas en los circuitos cerebrales sin necesidad de requerir modelos de aprendizaje.
Konrad Lorenz (23) ha descrito de forma elegante complejísimos rituales de cortejo y defensa en animales jóvenes y no duda que en los humanos también se presentan. Rapoport (17) considera que las conductas de las personas con trastorno obsesivo-compulsivo parecen recordar los modelos de acción determinados descritos por Lorenz y propone que quizá bajo la acción del estrés estos paquetes de información inteligente y programada se liberan y se ponen en marcha, ocasionando en el individuo la necesidad de desarrollar un comportamiento ritual o un pensamiento que parece hallarse cincelado en su cerebro con un importante grado de complejidad.
Si confrontamos el planteamiento de Rapoport (17) con nuestra concepción funcional de la corteza prefrontal, debemos señalar los siguientes puntos, que refuerzan la comprensión de la fisiología diferencial de las capas de la corteza:
• Los trastornos obsesivos-compulsivos están directamente relacionados con la actividad serotoninérgica (24). Si recordamos que las láminas superficiales son ricas en receptores 5HT, podemos inferir la relación entre la función cognitiva y esta lámina.
• Las ideas obsesivas se imponen al pensamiento.
La corteza prefrontal no reconoce las ideas obsesivas como propias y hasta se siente en la obligación de rechazarlas. Se plantea que los esquemas espaciotemporales que se corresponden con las ideas obsesivas se instalan directamente en la complejidad de la corteza prefrontal, por lo tanto, no la impulsan a crear una teoría, sino que la teoría ya viene desarrollada, pues se corresponde con paquetes de información biológicamente definidos.
Conclusión
Las últimas dos décadas han sido de fundamental importancia en el estudio y entendimiento del sistema nervioso. Este progreso no sólo ha incluido el abordaje de fenómenos subyacentes a la masa cerebral de tipo neuroquímico, neurofisiológico, neuroanatómico, citoarquitectónico, imagenológico, etc., sino la relación de estos conocimientos con aspectos complejos de la conducta y el psiquismo (25),(26),(27).
En este artículo, basados en la organización macro y microestructurla del sistema nervioso, se explicaron con detalle los fenómenos de procesamiento de la información en el cerebro y el resultado de las alteraciones de los sistemas de neurotransmisores en la génesis de las ideas delirantes, las experiencias de desrealización y despersonalización, las alucinaciones, las ilusiones, los trastornos del afecto y los trastornos de ansiedad. Sin embargo, el modelo descrito también permite dar cuenta de las consecuencias de las alteraciones estructurales y de la génesis psicopatológica de otras manifestaciones (28):
• Demencias: lesión estructural de la corteza prefrontal y alteración o inhabilitación de la función solucionadora de problemas de la corteza prefrontal. La alteración estructural puede iniciar o presentar cambios funcionales que dan cuenta no sólo de las alteraciones en las funciones de relación típicas de la demencia, sino también de las frecuentes manifestaciones que en este trastorno se ven en el pensamiento, como las ideas delirantes o las alucinaciones.
• Agnosias: lesión del componente perceptivo del sistema.
• Amnesias: lesión del componente mnésico del sistema.
Un modelo de funcionamiento mental debe tener la capacidad de abarcar diferentes tendencias (29). El psicoanálisis y el proceder psicoanalítico no se alejan de la aplicación del modelo expuesto en este artículo. El proceso de la transferencia descrito por Freud se puede considerar una forma controlada de generar incertidumbre en un individuo y una necesidad de la corteza prefrontal se puede asumir como una manera de "solucionar el problema" proyectando las propias vivencias (registradas en los módulos mnésicos) sobre un psicoterapeuta (aparece por el sistema perceptivo pero incógnito, cuando se deja de ver al acostarse en el diván). La transferencia es una forma de generar respuestas sobre sí mismo a través de un individuo neutral que es el terapeuta.
Bibliografía
1. Goldman-Rakic PS, Schwartz ML. Interdigitation of contralateral and ipsilateral columnar projections to frontal association cortex in primates. Science 1982; 14:755-7. [ Links ]
2. Fuster JM. The prefrontal cortex: anatomy, physiology, and neurophysiology of the frontal lobe. New York: Raven Press; 1989. p. 62-8. [ Links ]
3. Eccles J, Zeiher H. El cerebro y la mente. Barcelona: Herder; 1984. p. 51. [ Links ]
4. Popper K, Eccles J. El yo y su cerebro. Barcelona: Labor; 1985. p. 667. [ Links ]
5. Hubel BH, Wiesel TN. Mecanismos cerebrales de la visión: el cerebro. Barcelona: Labor; 1983. P. 114-28. [ Links ]
6. Benes FM, Majocha RE, Marotta CA. A modular arrangement of neuronal processes in human cortex: disruption with aging and in Alzheimer's disease. J Geriatr Psychiatry Neurol. 1988 Jan;1(1):3-10. [ Links ]
7. Changeux J. El hombre neuronal. Madrid: Espasa-Calpe; 1985. p. 231. [ Links ]
8. Gloor P. The role of the limbic system in experiential phenomena of temporal lobe epilepsy. Ann Neurol 1982;12(2):129-44. [ Links ]
9. Gloor P. Experiential phenomena of temporal lobe epilepsy: facts and hypotheses. Brain 1990;113(6):1673-94. [ Links ]
10. Gray JA. The psychology of fear and stress. Cambridge: Cambridge University Press; 1998. p. 380. [ Links ]
11. Goldman-Rakic PS, Lidow MS, Gallager DW. Overlap of dopaminergic, adrenergic, and serotoninergic receptors and complementarity of their subtypes in primate prefrontal cortex. J Neurosci 1990; 10(7):2125-38. [ Links ]
12. Llinás R. El cerebro y el mito del yo. Bogotá: Norma; 2003. p. 348. [ Links ]
13. Luria A. El cerebro humano y los procesos psíquicos. Fontanella; 1979. p. 533. [ Links ]
14. Dum RP, Strick PL. The origin of corticospinal projections from the premotor areas in the frontal lobe. J Neurosci 1991;11 (3):667-89. [ Links ]
15. Charney D, Nestler E, Bunney B. Neurobiology of mental illness. New York: Oxford University Press; 1999. p. 958. [ Links ]
16. Freud S. El yo y el ello. In: Obras completas. Biblioteca Nueva; 1981. p. 2700-28. [ Links ]
17. Rapoport J. Biología de las obsesiones y las compulsiones. Scientific American 1989;1982:59-64. [ Links ]
18. Piaget J. Biología y conocimiento. México: Siglo XXI; 1997. p. 338. [ Links ]
19. Ey H. La conciencia. Madrid: Gredos; 1976. p. 75-80. [ Links ]
20. Kovacs M, Beck AT. Maladaptive cognitive structures in depression. Am J Psychiatry 1978 May;135(5):525-33. [ Links ]
21. Kandel ER. A new intellectual framework for psychiatry. Am J Psychiatry 1998; 155:457-69. [ Links ]
22. Gorman JM, Liebowitz MR, Fyer AJ, Stein J. A neuroanatomical hypothesis for panic disorder. Am J Psychiatry 1989; 146 (2):148-61. [ Links ]
23. Lorenz K. La ciencia natural del hombre. Metatemas; 1993. p. 398. [ Links ]
24. Arango C, Angulo E, Pimienta H, Escobar M. Sistemas serotoninérgico y de norepinefrina. In: Sistema nervioso: neuroanatomía funcional, neurohistología, neurotransmisores, receptores y clínica. 3rd ed. Cali: Editorial Universidad del Valle; 2003. p. 245-53. [ Links ]
25. Arango C. Mecanismos celulares de los antidepresivos. Avances en Psiquiatría Biológica 2002;3:98-109. [ Links ]
26. Arango C. Aspectos moleculares y funcionales de los receptores, los neurotransmisores, los neuromoduladores y las hormonas. In: Gómez-Restrepo C, Hernández G, Rojas A, Santacruz H, Uribe M, editors. Fundamentos de psiquiatría clínica: niños, adolescentes y adultos. Bogotá: Centro Editorial Javeriano (CEJA); 2002. p. 49-59. [ Links ]
27. Arango C. Aspectos celulares y moleculares en psiquiatría. In: Gómez-Restrepo C, Hernández G, Rojas A, Santacruz H, Uribe M, editors. Fundamentos de psiquiatría clínica: niños, adolescentes y adultos. Bogotá: Centro Editorial Javeriano (CEJA); 2002. p. 36-48. [ Links ]
28. Arango C, Pimienta H, Escobar M. Depresión postisquemia cerebral: una aproximación fisiopatológica y clínica. Revista Colombiana Psiquiatría 2000; 24:321-44. [ Links ]
29. Arango C. Fundamento biológico de la vivencia de la corporeidad. Ciencias Humanas 2000;6:209-32. [ Links ]

