










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista de la Facultad de Medicina
Print version ISSN 0120-0011
rev.fac.med. vol.54 no.1 Bogotá Mar. 2006
INVESTIGACIÓN ORIGINAL
Luz Stella Rodríguez1, Carlos Arturo Guerrero F.2,
1. Química, Estudiante de Maestría en Bioquímica, Facultad de Medicina, Universidad Nacional de Colombia, Bogotá.
2.Profesor Asociado, Laboratorio de Biología Molecular y Celular de virus, Unidad de Bioquímica. Facultad de Medicina, Universidad Nacional de Colombia, Bogotá.
Correspondencia:guerra039@yahoo.com
Resumen
Introducción: los rotavirus son la principal causa de gastroenteritis en niños menores de cinco años. Hasta el momento se desconoce cómo entran estos virus a la célula huésped. La proteína de choque térmico, HSC70 se ha visto involucrada en la entrada del virus, sin embargo no se han relacionado bien todos los sitios de unión entre las proteínas virales del rotavirus y la HSC70.
Objetivos: el propósito de este trabajo fue determinar cuáles proteínas virales del rotavirus son responsables de la unión de la proteína de choque térmico, HSC70.
Material y métodos: se realizaron ensayos de coinmunoprecipitación utilizando lisados de células infectadas así como proteínas virales recombinantes. Estos ensayos se revelaron mediante SDS-PAGE y Western blot utilizando anticuerpos policlonales que reconocen las proteínas del rotavirus.
Resultados. En los experimentos realizados se encontró que la proteína viral VP6 coinmunoprecipita con HSC70 en las células MA104 durante la infección con rotavirus. La VP6, producida con vaccinia recombinante en MA104, también coinmunoprecipitó con HSC70.
Conclusión. La proteína VP6 de rotavirus interacciona con la proteína HSC70 en lisados de células MA104 infectadas con rotavirus. La unión de la VP6 recombinante a la HSC70 sugiere que la interacción de estas proteínas es independiente de otras proteínas virales.
Palabras claves: infección por rotavirus, vaccinia, reoviridae, rhesus (macaca mulatta).
Summary
Introduction: Rotaviruses are the main cause of gastroenteritis in children under five years old. At this moment is unknown how these viruses entry to host cell. The heat shock cognate protein HSC70 has been involved in rotavirus cell entry. However, the relationship between rotavirus proteins and HSC70 has not been established.
Objectives: the purpose of this work was to determine wich rotaviruses proteins interact with HSC70.
Material and methods: coimmunoprecipitation assays were done on a rotavirus infected supernatant and recombinant viral proteins. These assays were revealed by SDS-PAGE and Western blot using policlonal antibodys against rotavirus structural proteins.
Results: data show that viral protein VP6 coimmunoprecipated with HSC70 in MA104 cells during infection. Recombinant vaccinia of VP6 produced in MA104 cells also coimmunoprecipitated with HSC70.
Conclusions: these results suggest the interaction between VP6 and HSC70 during rotavirus infection. Furthermore, this union may be independent of other rotaviruses proteins.
Key words: rotavirus infections, vaccinia, reoviridae, rhesus (macaca mulatta).
Introducción
Los rotavirus constituyen el agente etiológico más importante de diarreas en niños. Afecta al 95 por ciento de la población menor de cinco años (1) e igualmente, infecta todas las especies animales de importancia económica para el hombre. Estos virus son transmitidos de manera fecal-oral e infectan las células epiteliales del intestino delgado en donde se replican y producen citólisis. Después de la replicación en el tracto gastrointestinal, son excretados y pueden ser dispersados en aguas, donde son resistentes a cambios fisicoquímicos, lo que facilita su transmisión (2). Estos virus pertenecen a la familia Reoviridae. El genoma viral consta de once segmentos de RNA de doble cadena, codifica para seis proteínas virales estructurales, (VP1, VP2, VP3, VP4, VP6, VP7) y seis proteínas no estructurales (NSP1-NSP6) (3-6). La infección eficiente por rotavirus, in vitro, se encuentra limitada a una serie de líneas celulares epiteliales de origen renal, como las de riñón de mono MA104 ó intestinal como las célul as de cáncer de colon CaCo-2 (4). Dos estudios previos sugieren que la proteína de choque térmico HSC70 puede estar involucrada en la entrada del virus a la célula (7,8). A pesar de los indicios, se desconoce si existe una unión entre las proteínas virales de rotavirus y esta proteína. La HSC70 pertenece a la familia de proteínas de choque térmico de 70KDa, esta familia se clasifica según su expresión en inducible y de expresión constitutiva perteneciendo a esta última la HSC70. Esta proteína se ha visto asociada a proteínas virales de poliomavirus, adenovirus, simian virus (SV40) y se ha detectado en viriones maduros cuyo material genético es RNA de doble cadena, como el virus de la estomatitis vesicular y el virus de la enfermedad del Newcastle, entre otros (9). En este trabajo investigamos la unión de HSC70 a las proteínas virales cuando las células MA104 están infectadas con rotavirus. Demostramos que la proteína viral, VP6, coinmunoprecipita con HSC70 en un lisado viral y para esta unión no se requiere de la presencia de otras proteínas virales.
Material y métodos
Células y condiciones de cultivo. La línea celular de mono rhesus, MA104, (obtenida del laboratorio del doctor Carlos Arias, Universidad Nacional Autónoma de México), se cultivó en eagle´s minimal essential medium (MEM), suplementado con 10 por ciento (v/v) de suero fetal bovino (SFB) y 100μg/ml de estreptomicina y penicilina en una incubadora de 5 por ciento CO2 a 37°C.
Virus. Partiendo de un lisado celular infectado, (obtenido del laboratorio del doctor Carlos Arias, Universidad Nacional Autónoma de México), el rotavirus RRV se activó durante 30 minutos a 37°C con tripsina (10μg/ml) en MEM. El inoculo viral se adicionó a las células MA104 confluentes, (previamente lavadas con MEM con el fin de retirar el SFB presente). Se dejó adsorber el virus por 45 minutos a 37°C. Posteriormente se retiró el inóculo viral no adherido y se adicionó medio de cultivo MEM. Las células infectadas se incubaron a 37°C hasta observar un efecto citopático completo (CPE), generalmente 24-48 horas post-infección. Las células infectadas se congelaron y descongelaron dos veces con el fin de liberar el virus intracelular y el adherido en la membrana. El lisado celular infectado se dividió en alícuotas y se congeló a 70°C hasta su uso. Para concentrar el virus, partiendo del lisado se centrifugó a 101.000 x g durante 90 minutos. El pellet fue resuspendido en buffer TSMC (0.01M Tris, 0.15M NaCl, 0.001 M MgCl2, 0.01M CaCl2).
Preparación de VP6 recombinante. Cuando las células MA104 obtuvieron confluencia se infectaron con vaccinia recombinante de VP6 (obtenida del laboratorio de los doctores Juanita Angel y Manuel Franco de la Pontificia Universidad Javeriana). 48 horas post infección se congelaron y descongelaron tres veces con el fin de desprender las células infectadas. Posteriormente, el lisado se resuspendió en buffer RIPA 4X, se centrifugó a 80000 x g durante 15 minutos, se retiró el sobrenadante y se llevó a un volumen final de 500ul.
Anticuerpos. Se usaron anticuerpos policlonales antirotavirus de conejo que reconocen todas las proteínas estructurales del rotavirus (donado por el laboratorio del doctor Carlos Arias, Universidad Nacional Autónoma de México). Anticuerpos policlonales de conejo generado con una proteína HSC70 recombinante hechos en Mexico (7) y Colombia. Como anticuerpo control se uso el policlonal de conejo dirigido contra la proteína c-neu (oncogene research).
Inmunoprecipitación. Se tomaron alícuotas de lisado de células infectadas con rotavirus (1000 UFF) en un volumen final de 100μl y con aproximadamente 10 μg de proteína cuantificada por el método de Bradford. Cada alícuota se incubó con 10μL de antiHSC70, utilizando como control el isotipo (ab-1) dirigido contra la oncoproteína c-neu. Las alícuotas se incubaron con el respectivo anticuerpo a 4°C toda la noche. Posteriormente, sobre cada alícuota se adicionaron 25μL de proteína A-sepharosa (50%p/v) y se agitaron suavemente por 30 minutos. El precipitado (proteína A-sepharosa-Antígeno-Anticuerpo) se lavó ocho veces, tres veces con buffer RIPA (50mM tris [pH 7.5], 150mM NaCl, 1 por ciento nonidet P-40 [NP-40], 0.5 por ciento deoxicolato de sodio, 0.1 por ciento dodecilsulfato de sodio [SDS], con 1mM de fenilmetilsulfonilfloruro [PMSF]); uno con buffer 50mM HEPES [(N-2hidroxietilpiperazina-N´-ácido etanosulfónico) pH: 7.0, 1% NP-40], uno con NaCl 2M, dos con buffer RIPA y dos con agua destilada. A cad a precipitado se le adicionó 25μL de buffer de muestra Laemmli 1X y se hirvieron durante cinco minutos para su análisis por Western blot.
Western Blot. Las muestras se separaron mediante electroforesis SDS-PAGE 10% (10). El gel se transfirió a membranas de PVDF (polivinildifluoruro), utilizando un sistema de western blot húmedo (mini trans-blot cell de Biorad) durante 16 horas a 100mA y 45V. Las membranas se bloquearon una hora con leche descremada en PBS solución 5 por ciento (p/v). Luego se incubaron con el anticuerpo primario, antirotavirus hecho en conejo, que reconoce todas las proteínas virales estructurales, seguido por un anticonejo biotina. La membrana se reveló con ABC (avidina-biotina-peroxidasa) y luminol super signal west dura extended de Pierce.
Resultados
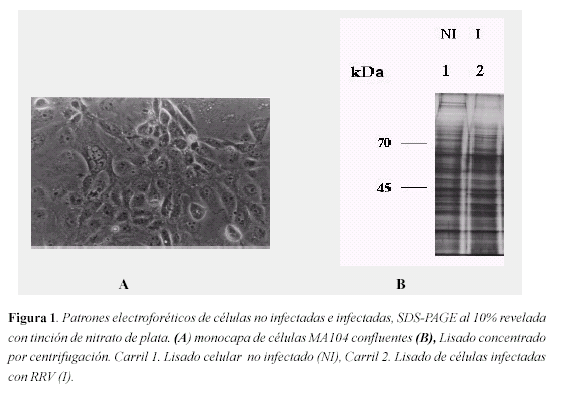
Patrones electrofréticos de células no infectadas e infectadas. Para conocer el patrón electroforético de un lisado de células MA104 no infectadas (Figura 1 A), y de células infectadas con el rotavirus RRV, se realizaron electroforesis SDS-PAGE al 10 por ciento y se revelaron mediante tinción con nitrato de plata. En la figura 1B, se observa el diferente patrón de corrido entre los lisados de células no infectadas (Carril 1) y lisado de células infectadas (Carril 2).
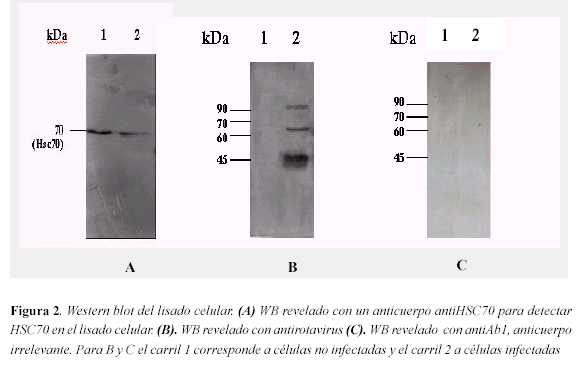
Coinmunoprecipitación de las proteínas virales con HSC70. Para determinar las interacciones de HSC70 con las proteínas del rotavirus primero descartamos que los anticuerpos usados en los procedimientos hicieran reconocimientos inespecíficos en los lisados celulares; como se muestra en la figura 2A el anticuerpo contra HSC70 reconoce específicamente a la proteína tanto en el lisado de células no infectadas como infectadas con rotavirus, sin reconocer proteínas virales. Así mismo, el anticuerpo policlonal que reconoce rotavirus no reconoce proteínas celulares y el anticuerpo usado como irrelevante para el control en la inmunoprecipitación (antiAb1) no reconoce ninguna proteína viral de rotavirus, figura 2B y 2C.
Con el fin de elucidar qué proteínas virales se unen con HSC70, se realizaron inmunoprecipitaciones con el antiHSC70 del lisado celular. Dicho lisado habia sido previamente concentrado por ultracentrifugación de células infectadas con rotavirus. Como se observa en la figura 3A coimunoprecipitaron con HSC70 cuatro bandas de 45, 55, 90 y mayor de 130kDa. Se sabe que la proteína viral VP6 tiene un peso de ~45kDa, la cadena pesada de la inmunoglobulina ~55Kda, VP4 ~90kDa y probablemente la banda de ~130Kda sean agregados virales.
Coinmunoprecipitación de VP6 con HSC70. El resultado anterior sugiere que VP6, la proteína mayoritaria del virión, se une a HSC70. Para corroborarlo y determinar si esta proteína puede unirse a HSC70 sin la presencia de las demás proteínas virales, inmunoprecipitamos VP6 recombinante expresada en vaccinia y producida en células MA104. Como se observa en la Figura 3B, VP6 se une en el lisado celular a HSC70, sugiriendo una unión específica.
Estos resultados fueron reproducibles en experimentos realizados por triplicado con cultivos celulares de MA104 y lisado de células infectadas con RRV producidos en diferentes tiempos o al utilizar otro anticuerpo policlonal dirigido contra HSC70 producido en el grupo de biología molecular de virus de la Facultad de Medicina de la Universidad Nacional, utilizando como control de la inmunoprecipitación el respectivo suero preinmune (dato no mostrado).
Discusión
La HSC70 es una proteína constitutiva de la familia de HSP70 que se encuentra dentro y fuera de la célula, con una estructrura similar al complejo mayor de histocompatibilidad tipo I y a la actina, tiene funciones variadas, aunque de estas las más conocidas son: como chaperona y contribuyente en el correcto plegamiento de las proteínas (11-14).
El mecanismo de entrada del rotavirus es un proceso de múltiples pasos en el cual se presentan interacciones secuenciales entre las proteínas de la cápside externa del virus y las moléculas de superficie celular durante el proceso de adherencia y penetración celular (15-20). Las interacciones tempranas del rotavirus con la célula hospedera son un proceso complejo, que probablemente ocurre por interacciones secuenciales de las proteínas de la superficie de los virus VP4 y VP7 con al menos cuatro moléculas de superficie celular diferentes, dispuestas en microdominios lipídicos "rafts" (15-20). La VP6 es la proteína más inmunogénica de los rotavirus, pero no induce anticuerpos neutralizantes, aunque algunas inmunoglobulinas IgA específicas para VP6 son protectivas in vivo, probablemente vía trancitosis a través de células epiteliales (21). Estudios dirigidos al desarrollo de vacunas han encontrado una elevada protección al utilizar VP6 como inmunógeno mediante diferentes estrategias (22-24). Los péptidos sin téticos que tienen la región 283-307 de VP6 indujeron una completa protección al desafió viral al inmunizar ratones con estos péptidos (23). El mecanismo mediante el cual la proteína VP6 protege del desafío con rotavirus no se encuentra bien establecido, varios estudios apuntan a que esta protección se debe a neutralización intracelular de partículas DLPs por inmunoglobulinas IgA que impiden la liberación y traducción de transcritos al citoplasma (21-24).
Existen diferencias parciales en el patrón electorforético de células MA104 no infectadas e infectadas con rotavirus. Previamente se había evidenciado la interacción directa entre la HSC70 y las proteínas virales del polyomavirus (9). En nuestro trabajo, las proteínas virales VP4 y VP6 coinmunoprecipitan con la proteína celular de choque térmico HSC70 a partir de un lisado de células previamente infectadas con rotavirus sugiriendo que las dos proteínas interacionan con HSC70 durante el proceso infeccioso. La inmunoprecipitación se llevó a cabo con un anticuerpo policlonal contra HSC70 y la VP6 se detectó con un anticuerpo policlonal que reconoce todas las proteínas estructurales del virus. No disponiamos de anticuerpos específicos contra VP4 para corroborar su identificación, pero por el peso molecular y dado que el anticuerpo no reconoce proteínas celulares es muy probable que se trate de la proteína del rotavirus.
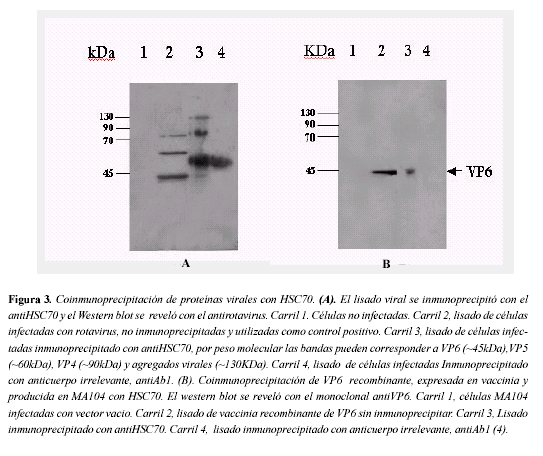
Puesto que están coinmunoprecipitando VP4 y VP6 es difícil determinar si existe unión física de VP4 o de VP6 con HSC70 o si solo se está uniendo una de las dos proteínas virales y la otra se detecta porque permanecen unidas las proteínas del rotavirus, es decir, se precipitan como un complejo. Para resolver esto inmunoprecipitamos HSC70 de un lisado celular que contenia VP6 recombinante, expresada en vaccinia. El resultado indica que VP6 coinmunoprecipita con HSC70 sugiriendo que la unión es independiente de otras proteínas virales como VP4, VP7 o VP2, proteínas que interactúan con VP6 en el virión de rotavirus. Sin embargo, con la técnica empleada en este trabajo no podemos descartar que la VP6 se esté uniendo a otra proteína celular y que dicha proteína a su vez esté ligada a HSC70 y por eso la VP6 esté coinmunoprecipitando con HSC70. Para resolver esto sería interesante realizar ensayos de unión teniendo VP6 purificada y HSC70 recombinante humana, para confirmar que existe una unión directa entre ambas .
Sin embargo, recientemente encontramos que al realizar ensayos de unión de HSC70 con TLPs, es decir, la partícula viral completa y DLPs, la partícula viral sin la capa externa (sin VP7 y VP4) mediante ELISA, la HSC70 recombinante humana se une a ambos tipos de partículas (artículo en preparación).
Agradecimientos
A la doctora Juanita ángel y el doctor Manuel Franco del Instituto de Geneética de la Pontificia Universidad Javeriana por su colaboración en el desarrollo del trabajo. Este proyecto fue financiado por COLCIENCIAS (código 1101-04-12011).
Referencias
1. Ball JM, Tian P , Zeng CQY, Morris AP, Estes MK, Age-dependent diarrhea induced by a rotaviral nonstructural glycoprotein. Science. 1996;272:101-104. [ Links ]
2. Dubois E, Guyader Le F, Haugarreau L, Kopecka H, Cormier M, Pommepuy M. "Molecular Epidemiological Survey of Rotaviruses in Sewage by Reverse Transcriptase Seminested PCR and Restriction Fragment Length Polymorphism Assay". Appl. Environ Microbiol. 1997;63: 1794-1800 [ Links ]
3. Estes MK. "Rotaviruses and their Replication". Fields, B.N. Knipe, D.M. and Howley, eds. En Virology, Philadelphia: Lippencott Raven, 1996; 1625-1655. [ Links ]
4. Granoff A, Webster R. "Rotaviruses" Encyclopedia of Virology, San Diego, USA, Academic Press, 2da. Edición.1999;1576-1578. [ Links ]
5. Leite J, Alfieri AA, Woods PA, Glass RI, Gentsch JR. "Rotavirus G and P types circulating in brazil: characterization by RT-PCR, probe hybridization, and sequence analysis". Arch Virol. 1996;141: 2365-2374. [ Links ]
6. Lundgren O, Peregrin AT, Persson K, Kordasti S, Uhnoo I, Svensson L. Role of the enteric nervous system in the fluid and electrolyte secretion of rotavirus diarrhea. Science. 2000;287:491-495. [ Links ]
7. Guerrero CA, Bouyssounade D, Zarate S, Isa P, López T, Espinosa R, Romero P, Méndez E, López S, Arias CF. "Heat Shock Cognate Protein 70 Is Involved in Rotavirus Cell Entry". J. Virol. 2002; 76: 4096-4102. [ Links ]
8. Zárate S, Cuadras MA, Espinosa R, Romero P, Juárez KO, Camacho-Nuez M, Arias CF, López S. Interaction of Rotaviruses with HSC70 during Cell Entry Is Mediated by VP5. J Virol. 2003; 77 (13): 7254-7260 [ Links ]
9. Cripe T, Delos S, Estes P, Garcea R. "In Vivo and In Vitro Association of Hsc70 with Polyomavirus Capsid Proteins". J. Virol. 1995;69:7807- 7813. [ Links ]
10. Laemmli UK. " Cleavage of structural proteins during the assembly of the head of bacteriophage t4". Nature 1970;227:680-685. [ Links ]
11. Chappell TG, Welch WJ, Schlossman DM, Palter KB, Schlesinger MJ, Rothman JE. Uncoating ATPaes is a member of the 70 kilodalton family of stress proteins. Cell. 1986;45:3-13 [ Links ]
12. Chou Ch, Forouhar F, Yeh Y, Hui-Lin Shr, Wang Ch, Chwan-Deng Hsiao. Crystal Structure of the C-terminal 10-kDa Subdomain of HSC70. J. Biol. Chem. 2003; 278:30311-30316 [ Links ]
13. Flaherty KM, Mackay DB, Kabsch W, Holmes KC. Similarity of the three-dimensional structures of actin and the ATPase fragment of a 70-kDa heat shock cognate protein. Proc. Natl. Acad. USA. 1991;88:5041-5045 [ Links ]
14. Frydman J, Harti FU. Molecular chaperone functions of hsp70 and hsp60 in protein folding. The Biology of heat shock proteins and molecular chaperones. Cold spring Harbor Laboratory Press:1994;251-283. [ Links ]
15. Arias CF, Isa P, Guerrero CA, Mendez E, Zarate S, López T, López S. Molecular biology of rotavirus cell entry. Arch. Med. Res. 2002 ;33: 356- 36 [ Links ]
16. Guerrero CA, Zarate S, Corkidi G, López S, Arias C. Biochemical characterization of rotavirus receptor in MA104 cells. J Virol. 2000; 74;93-102. [ Links ]
17. Méndez E, López S, Cuadras MA, Romero P, Arias CF. Entry of rotaviruses is a multistep process. Virology. 1999;263:450-459. [ Links ]
18. Sánchez-San Martin C, Lopez T, Arias CF, Lopez S. Characterization of rotavirus cell entry. J. Virol. 2004;78, 2310- 2318. [ Links ]
19. Zárate S, Espinosa R, Romero P, Méndez E, Arias CF, López S. The VP5 domain of VP4 can mediate the attachment of rotaviruses to cells. J. Virol. 2000. 74:593-599 [ Links ]
20. Isa P, Realpe M, Romero P, López S, Arias CF. Rotavirus RRV associates with lipid membrane microdomains during cell entry. Virology. 2004;322:370-381 [ Links ]
21. Burns J, Siadat-Pajouh, Krishnaney A, Greenberg H. Protective effect of rotavirus VP6-specific IgA monoclonal Antibodies that lack neutralizing activity. Science. 1996;272: 104-107 [ Links ]
22. Jacobson RM. The current status of the rotavirus vaccine. Vaccine.1999; 17:1690-1699. [ Links ]
23. Choi A, Basu M, Mcneal M, Clements J, Ward R. Antibody-independent protection against rotavirus infection of mice stimulated by intranasal immunization with chimeric VP4 or VP6 protein. J. Virol. 1999;73:7574-7581. [ Links ]
24. Feng N, Lawton J, Gilgert J, Kuklin N, Vo P, Prasad B, Greenberg H. Inhibition of rotavirus replication by a non-neutralizing, rotavirus VP6-specific IgA mAb J Clin Invest.2002;109:1203-1213 [ Links ]
