










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista de la Facultad de Medicina
Print version ISSN 0120-0011
rev.fac.med. vol.59 no.2 Bogotá Apr./June 2011
Actualización
ALERGIA EN EL HUMANO INDUCIDA POR LA SALIVA DE INSECTOS DE LA FAMILIA CULICIDAE
Allergy in humans induced by saliva from insects from the family culicidae
Lígia Inés Moncada-Álvarez1, Myriam Janeth Salazar-Terreros2, Myriam Consuelo López-Páez1
1Departamento de Salud Pública, Facultad de Medicina, Universidad Nacional de Colombia, Bogotá.
2Maestría en Microbiología, Facultad de Ciencias, Universidad Nacional de Colombia, Bogotá.
Correspondencia: limoncadaa@unal.edu.co
Resumen
Se hace una revisión de las moléculas que inoculan los insectos de la familia Culicidae al momento de la picadura y los mecanismos que muestran sus hospederos para contrarrestarlos y cómo algunas de esas moléculas, especialmente las enzimas se convierten en alérgenos que inducen una respuesta de amplio espectro, que va desde una pápula al momento de la picadura hasta una reacción anafiláctica. De la misma manera se analizan las posibilidades de diagnóstico con mol&oecute;culas silvestres y antígenos recombinantes, lo mismo que pautas de tratamiento.
Palabras clave: hipersensibilidad, saliva, mordeduras y picadura de insectos, insectos, culicidae, alergenos.
Summary
A review was made of the molecules which are inoculated by insects from the family Culicidae when biting and the mechanisms used by their hosts to contract them, as well as some of these molecules, especially enzymes which become allergens inducing a broad spectrum response ranging from a papule when being bitten to a full-blown anaphylactic reaction. The possibility of diagnosis using wild molecules and recombinant antigens was also analyzed, as well as treatment guidelines.
Key word: hypersensitivity, saliva, insect bites and stings, insects, cilicidae, allergens.
Introducción
Los insectos conocidos vulgarmente como “zancudos” o “mosquitos” pertenecen a varios géneros de la familia Culicidae. Las hembras se alimentan de sangre, esta característica las hace vulnerables a infectarse y transmitir diferentes agentes patógenos al hombre y a los animales. De acuerdo con el patrón de alimentación, Tempelis (1) reconoce seis patrones básicos de preferencia de comida:
- Casi completamente sobre mamíferos, en este grupo hay un rango en la preferencia desde los completamente antropofílicos como An. darlingi hasta los completamente zoofílicos como An braziliensis.
- Los que son casi completamente ornitofílicos ejemplo: Culiseta melanura,
- Los que ingieren sangre de aves y de mamíferos ejemplo: Culex quinquefasciatus,
- Los que se alimentan casi exclusivamente sobre anfibios ejemplo: Culex terrinas,
- Los que se alimentan sobre reptiles Deinocerites dyari,
- La especie Uranotenia lateralis que come sobre un pez que vive en bancos de lodo fuera del agua.
Las reacciones a la picadura de los insectos se puede dar por las densidades que pueden alcanzar, se ha reportado que en zonas donde la malaria es endémica, los humanos pueden recibir 200 picaduras de Anopheles por día, lo que significaría que recibirían aproximadamente 10.000 picaduras al año (2,3). Debido a que Culex quinquefasciatus se cría en colecciones de aguas tanto limpias como contaminadas, en los centros urbanos ha alcanzado densidades muy altas y se ha convertido en un problema de salud pública por la molestia que representa debido a su hábito nocturno, lo que impide el descanso de los habitantes de las zonas altamente infestadas y por la alergia que producen por su picadura (4-6).
La saliva de los mosquitos
Los mosquitos son los artrópodos hematófagos más comunes y de mayor importancia en el mundo. Hasta el momento se han descrito cerca de 3000 especies y sólo un pequeño porcentaje de ellas necesita ingerir sangre para el desarrollo de sus huevos.
Generalmente, la picadura de los mosquitos es inocua, pero en muchas ocasiones puede convertirse en el vehículo de transmisión de agentes patógenos, y desencadenar reacciones inmunológicas indeseables en el hospedero; esta respuesta es producida en su mayor parte por las proteínas que se encuentran en la saliva de los insectos, que es una mezcla compleja de diversas sustancias secretadas por las glándulas salivares tanto en machos como en hembras; sin embargo, existen proteínas que solo se encuentran en uno u otro género, debido al hecho de que las hembras son las únicas que se alimentan de la sangre de los vertebrados, mientras que los machos se alimentan exclusivamente de exudados vegetales.
Estas características de la dieta, corresponden con diferencias morfológicas y fisiológicas en las glándulas salivares. En todos los géneros de mosquitos hematófagos las glándulas de los machos están compuestas de tres lóbulos similares, mientras que en las hembras, el lóbulo central tiene tamaño diferente. La porción anterior de los lóbulos laterales de las hembras es morfológica y funcionalmente similar a las glándulas de los machos ya que producen enzimas involucradas en la digestión de los azúcares, pero las células de la porción distal de los lóbulos laterales y del lóbulo central son distintas y específicas de las hembras y están relacionadas con la hematofagia (7). La salivación de los mosquitos puede ser regulada directamente por factores químicos, térmicos o táctiles, o indirectamente a través de estímulos generados por el mismo proceso de la alimentación (8).
Para localizar el vaso sanguíneo deben introducir varias veces sus partes bucales en la piel del hospedero hasta alcanzar la red profunda de vasos capilares, la saliva es secretada durante este período de búsqueda intradérmica (probing o tanteo) (9), en el que se acumulan pequeñas hemorragias, hasta que finalmente las partes bucales descansan dentro de una vénula, arteriola o un hematoma (10,11).
Entre las funciones de la saliva de los mosquitos está la lubricación y protección de las piezas bucales (11,12), facilitar la localización de los vasos sanguíneos y proporcionar enzimas como α-glucosidasas y α-amilasas que inician la digestión de los azúcares de la dieta (11,13,14). También se ha demostrado que la saliva de los mosquitos interactúa con moléculas propias de la respuesta inmune de los vertebrados y modula su expresión, de forma tal que esta relación puede facilitar las infecciones por microrganismos (15,16). Varios autores han demostrado la participación de proteínas y otros componentes de la saliva en la respuesta inmunosupresora local y alérgica del hospedero (17-21).
Los vertebrados han desarrollado mecanismos inmunológicos, inflamatorios y de hemostasis para contrarrestar el daño producido por la picadura de los insectos hematófagos. La hemostasis es una respuesta fisiológica compleja que consta de varios procesos como la agregación plaquetaria, la coagulación sanguínea y vasoconstricción, todos los cuales contribuyen a disminuir la pérdida de sangre ocasionada por el deterioro de un vaso sanguíneo. La inflamación es una respuesta del hospedero a un daño tisular, caracterizada por dolor y vasodilatación local, lo que conlleva al enrojecimiento y aumento de temperatura en la zona afectada; adicionalmente, la exposición de determinantes antigénicos en este contexto inflamatorio, activa el sistema inmune del hospedero para que reaccione contra estos estímulos (22).
A su vez a lo largo de su historia evolutiva, los insectos hematófagos han desarrollado sistemas para evadir todos estos mecanismos, principalmente a través de proteínas presentes en la saliva que incluyen factores anticoagulantes, antiplaquetarios, vasodilatadores, antiinflamatorios e inmunosupresores compuestos por moléculas bioactivas que son inoculadas en la piel del hospedero durante la picadura (7).
Por varias décadas se han descrito las propiedades de la saliva de los mosquitos que contribuyen a evitar la agregación de plaquetas y prevenir la hemostasis del hospedero (11). Las funciones antihemostáticas e inmunomoduladoras también se piensa que pueden tener algún papel que favorezca la transmisión de los microorganismos que son portados por aquellas especies vectoras (23,24).
Adicionalmente a los factores descritos anteriormente, la saliva de los mosquitos ha sido identificada como fuente de toxinas, inmunógenos y alergenos que son reconocidos por el sistema inmune de los vertebrados. Las reacciones alérgicas a la saliva son consideradas como funciones del hospedero que disminuyen el éxito de la alimentación de los artrópodos hematófagos. Debido a que la saliva parece tener efectos alérgicos contraproducentes para estos insectos, se ha postulado que las moléculas involucradas podrían representar vestigios de aquellas presentes en la saliva de ancestros fitófagos (11). Las reacciones del hospedero a los componentes de la saliva de los mosquitos varían desde la aparición de pequeñas pápulas hasta el shock anafiláctico en casos muy esporádicos, pasando por diversos estados inflamatorios y grados de prurito, dependiendo de la especie de mosquito (25).
Inmunología de la reacción a la picadura de los mosquitos
La fisiopatología de la respuesta inmune a la picadura de mosquitos es compleja e involucra diferentes factores que incluyen susceptibilidad genética, ruta de exposición, dosis de alergenos y en algunos casos su estructura (26). La sensibilización a los alergenos resulta en la producción de citocinas Th2 como la IL-4 e IL-13 que genera la producción de IgE por los linfocitos B. La IgE sensibiliza los mastocitos y basófilos que se unen al receptor de alta afinidad para la IgE (FceRI) expresado en su superficie, lo que induce su degranulación y la liberación de aminas vasoactivas (principalmente histamina), mediadores lipídicos como prostaglandinas y leucotrienos, citocinas y quimoquinas características de la fase inmediata de la reacción alérgica.
Después de la fase de sensibilización, la inflamación alérgica se puede ver no solo en el órgano blanco sino que se puede convertir en anafilaxis sistémica (27,28). En individuos sensibilizados, la tolerancia de células T representa el mecanismo clave en la respuesta inmune saludable a los alergenos.
Normalmente, hay cinco etapas en el curso de la reacción a la picadura de los mosquitos. Cuatro de estas etapas las describió Mellanby (29), expuso 25 individuos a la picadura de Aedes aegypti. Cuando se expusieron por primera vez la respuesta cutánea se redujo a un punto rojo de 1mm de diámetro en el sitio de la picadura, sin comezón. Luego de 24 a 48 horas se presentó una reacción tardía caracterizada por la aparición de una mancha roja de 3 cm. de diámetro que rodeaba el sitio de la picadura, cuyo centro se transformó posteriormente en una roncha diminuta. Esta condición perduró varios días y algunas veces mostró fluctuaciones en los diámetros de la roncha y la mancha. Al cabo de un mes de exposición continua, se presentaba una reacción inmediata justo después de la picadura, que se manifestó como una roncha rodeada de un área eritematosa.
Luego de exposiciones adicionales, la reacción inmediata permanecía sin cambios pero la reacción tardía perdía severidad gradualmente. La quinta etapa, se caracteriza por la desaparición de las reacciones inmediatas y tardías y es conocida como desensibilización (30). A partir de los estudios de Mellanby y otros investigadores como Heilesen se hizo una clasificación de las reacciones ante las picaduras de los mosquitos, que aún hoy tiene validez (Tabla 1).
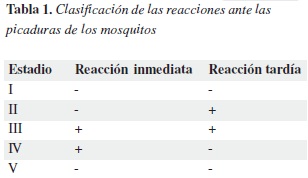
La alergia a la picadura de los insectos se define como reacción tipo I inmediata o retardada mediada por anticuerpos IgE y se desencadena por la inoculación de un alergeno en la picadura y las sustancias vasoactivas y enzimas son los posibles agentes de hipersensibilización (31). La reacción clínica puede ser del tipo tuberculina, urticaria o eczematoide. Estas reacciones han sido replicadas mediante el empleo de parches y pruebas intradérmicas en extractos acuosos crudos de mosquitos.
Las reacciones a la inoculación de fluidos salivares pueden ser inmediatas, con la aparición de ronchas acompañadas de eritema y prurito a los pocos minutos, o tardías que aparecen algunas horas después y se caracterizan por lesiones papulares, asociadas frecuentemente con edema y prurito que persisten por varias horas (29). También se pueden producir reacciones como urticaria y angioedema, síntomas menos definidos como náuseas, dolor de cabeza y letargo (32).
Los hallazgos característicos son eritema localizado, calor, prurito y dolor (29). En personas sin sensibilización previa, las reacciones pueden ser leves o severas, se pueden desencadenar respuestas de tipo I (hipersensibilidad inmediata dependiente de IgE) y de tipo IV (hipersensibilidad retardada mediada por células); los mastocitos son los mediadores principales de la respuesta y han sido relacionados con procesos de modulación de la respuesta inmune. Los factores liberados por estas células son los responsables de la formación de pápulas, la sensación de quemadura, el edema y el prurito luego de una picadura. Estas reacciones indican la naturaleza alergénica de algunas moléculas presentes en la saliva.
Los casos de hipersensibilidad retardada exagerada a la picadura de los mosquitos son escasos en personas adultas; sin embargo, en pacientes con patologías crónicas basales se pueden producir este tipo de reacciones, caracterizadas por el desarrollo lento de las lesiones (entre 12 y 24 horas posteriores a la exposición), induración, edema, eritema, prurito intenso y lesiones más graves (33). Ocasionalmente se puede desarrollar una reacción complicada, pero un alto porcentaje de estos pacientes tienen una historia familiar de alergias (29,31). En muy pocas ocasiones, se reportan casos de shock anafiláctico producido por la picadura de los mosquitos (34,35). La severidad de la reacción depende de la sensibilidad personal y la gravedad de las reacciones locales está vinculada al rascado y al número de picaduras (31).
De acuerdo con algunos estudios realizados en ratones, la picadura de A. stephensi induce la degranulación de los mastocitos dérmicos, conduciendo a la extravasación de los fluidos y el reclutamiento de neutrófilos. Luego de la activación de los macrófagos, se produce una hiperplasia de los nódulos linfáticos debido a la acumulación de leucocitos CD3+, B220+ y CD11c+. Estos hechos conllevan al enriquecimiento de los nódulos con linfocitos T y comprueban que los mastocitos periféricos juegan un papel importante en el reclutamiento de células efectoras y por ende, en la activación subsecuente de la respuesta humoral; la degranulación de los mastocitos se produjo de manera independiente de IgE. La activación de los mastocitos conducen a la liberación de varios mediadores tales como histamina, FNTα y eicosanoides (36).
Estudios posteriores utilizando el mismo modelo animal, demostraron que la inflamación mediada por mastocitos regula la magnitud de la respuesta inmune específica contra los antígenos de la saliva de los mosquitos, por medio del aumento en la producción de MIP-2 e IL-10 (37). Por otra parte, ratones expuestos a múltiples picaduras de An. stephensi producían mayores niveles de IFN-γ (respuesta Th1) y menores niveles de IL-4 (respuesta Th2) que los controles; la reacción no fue solamente local sino que se detectaron niveles elevados de esta citocina a nivel del hígado y el suero (38). Sin embargo, cuando se observó la producción de citocinas en esplenocitos, se detectó que el IFN-γ se redujo hasta el séptimo día luego de la exposición a las picaduras, mientras que la IL-4 y la IL-10 aumentaban durante el mismo período (39).
Se ha descrito que en la saliva de los mosquitos existe una sustancia que inhibe la producción de FNT-α, liberado por los mastocitos (40); por otra parte, también se ha registrado una actividad inmunomodulatoria en lisados de glándulas salivares de Ae. aegypti, que se caracteriza por suprimir in vitro, la producción de IFNγ e IL-2 y la inhibición de la proliferación de células T dependientes de esta (41).
Se han hallado anticuerpos IgG específicos contra proteínas de mosquitos en sueros humanos pero su papel en el desarrollo de sensibilidad no se ha aclarado (32). Se ha establecido que las respuestas IgG e IgE específicas corren paralelas a las reacciones cutáneas en humanos, de manera que la IgG puede estar involucrada en el desarrollo de la alergia a la picadura de los mosquitos (42). Sin embargo, en ensayos con conejos la IgG específica parece actuar como un anticuerpo bloqueador y se correlaciona con el desarrollo de la desensibilización.
La IgG específica antimosquito ha sido identificada en el suero humano utilizando la prueba de ELISA y otras pruebas inmunohistoquímicas; estos anticuerpos son principalmente de las subclases IgG4 e IgG1. Investigaciones previas hallaron niveles más altos de IgG específica en adultos que en niños y los niveles mayores en trabajadores forestales, sugiriendo que esta clase de anticuerpos se relaciona con la exposición a las picaduras (Ailus et. al, 1985) (43). Otros investigadores hallaron niveles de IgG que se incrementan hasta los dos años y luego se estabilizan (Konishi, 1990) (44).
Los resultados de Peng et al (19) revelan que no solamente se incrementa la IgE específica en sujetos con reacciones cutáneas, sino que también lo hace la IgG. Wilson y Clements (en 32), relacionaron la presencia de anticuerpos precipitantes en el suero humano con el desarrollo de reacciones cutáneas, mientras que otros hallaron que sujetos con una exposición extensa a las picaduras tienen poco o ningún nivel detectable de IgG específico. Sin embargo, es importante destacar que este tipo de anticuerpos específicos contra los componentes antigénicos de la saliva de las especies de mosquitos de una región son encontrados frecuentemente en la población expuesta Por otro lado, se ha demostrado la reactividad cruzada entre especies de mosquitos en sueros de pacientes alérgicos a la picadura (45).
Se ha planteado también el uso potencial de las respuestas inmunes a la saliva de los mosquitos como un método indirecto para medir la exposición a las picaduras de especies vectoras; en un estudio de esta clase, se evaluó mediante inmunoblot la respuesta IgG en niños expuestos a la picadura de An. gambiae en una zona endémica para malaria; en este estudio se identificaron anticuerpos específicos contra varias bandas, principalmente la de 72 y 175 kDa. La respuesta contra la banda de 175 kDa era más fuerte durante el período de transmisión máxima de malaria (46).
Se ha sugerido que puede existir hipersensibilidad a la picadura de los mosquitos mediada por células T, de manera similar a lo que ocurre con la alergia a las abejas, avispas y hormigas; se ha logrado la transferencia de reacciones cutáneas tardías al introducir células del bazo de animales sensibilizados en conejillos de Indias; por otra parte, estudios de proliferación de linfocitos incubados con extractos de mosquitos muestran correlación con las reacciones tardías; estos resultados sugieren que existe hipersensibilidad del tipo IV en la alergia a la picadura de los mosquitos (32,47,48).
Las complicaciones debidas a la picadura se asocian a factores de riesgo como inmunodeficiencia, y la no exposición previa a las picaduras de una especie determinada. En algunas poblaciones de ascendencia asiática se han descrito reacciones necróticas en la piel, asociadas a un síndrome hemofagocítico. La presentación clínica de este síndrome se relaciona con la presencia de leucemia de células NK (Natural killer), cuando las células están infectadas con el virus de Epstein-Barr (49).
Moléculas involucradas en las reacciones inmunes a la picadura de los mosquitos
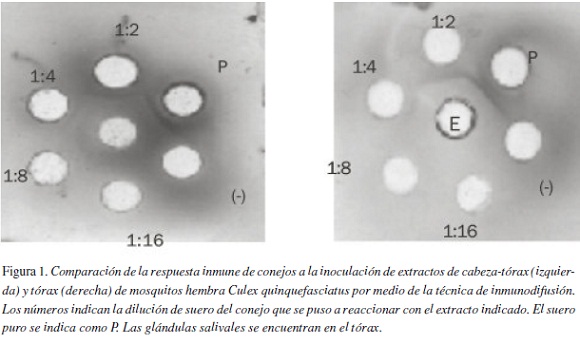
Por lo general, el descubrimiento de las proteínas salivares de interés se inicia con la identificación de los componentes activos de la saliva en los extractos de glándulas salivares, para finalmente obtener la secuencia genética y la estructura primaria de la proteína de interés (50). Frecuentemente, se utilizan modelos animales para establecer las propiedades antigénicas de los extractos de mosquitos (Figura 1) (51). Sin embargo la producción de extractos de glándulas salivares es un proceso laborioso y el volumen obtenido es escaso, por lo que se han utilizado extractos de cuerpo entero de los mosquitos en los estudios de caracterización de antígenos y de la respuesta inmune. La identificación y caracterización de los antígenos de los mosquitos, especialmente aquellos compartidos por varias especies, pueden ser utilizadas en estudios relacionados con el diagnóstico e inmunoterapia de la alergia a la picadura de estos insectos.
La alergia a la picadura de mosquitos es un problema común y generalizado en todo el mundo para el que no hay un tratamiento efectivo estándar. La inmunoterapia con extractos de mosquito no es de uso generalizado, puesto que los extractos disponibles comercialmente para realizarla no han sido caracterizados según su composición proteica, antigénica y actividad biológica (25,32).
La comparación de perfiles electroforéticos de 12 especies de mosquitos (Aedes aegypti, Ae. albopictus, Ae. communis, Ae. punctor, Ae. togoi, Anopheles darlingi, An. gambiae, An. stephensi, Culex molestus, Cx. pipiens, Cx. quequectates y Cx- quinquefasciatus), mostró que las bandas halladas con mayor frecuencia son las de 16, 27, 64, 65 kDa (60% de las especies) y 66 kDa (70%); lo que puede indicar un alto grado de conservación de estas proteínas dentro de la familia Culicidae.
En cuanto a las bandas de pesos moleculares bajos (10-15 kDa), son más comunes en los representantes del género Culex; mientras que el rango de 65-70 kDa es compartido por los géneros Culex, Aedes y Anopheles. Algunas de las bandas que aparecen en el perfil electroforético de la saliva de una especie determinada, no aparecen en el perfil obtenido a partir de los extractos de glándulas salivares.
Tales proteínas pueden provenir del intercambio activo que ocurre entre la hemolinfa y la saliva en el estilete del mosquito, por lo cual no se producen ni almacenan en las glándulas salivales (Shroek, en 52). Se han aislado y caracterizado bioquímicamente varias proteínas a partir de las glándulas salivares de varias especies de mosquitos y que al parecer están involucradas en la hematofagia, tales como la apirasa y muchos de los antígenos compartidos entre diferentes especies de mosquitos reconocidos por anticuerpos IgE e IgG específicos contra la saliva de estos insectos. Desde el punto de vista de su antigenicidad, los géneros Aedes, Culex y Culiseta, comparten un 23% de antígenos salivares, lo que indica que sus mecanismos bioquímicos de hematofagia podrían ser similares, y por tanto, producir respuestas antigénicas similares en los hospederos vertebrados (51).
Los sialotranscriptomas de An. darlingi (especie del Nuevo Mundo) y An. gambiae (especie del Viejo Mundo), muestran que el 53% de las proteínas salivares son diferentes y algunas de las proteínas de An. darlingi están más relacionadas con las proteínas propias de las especies de la subfamilia Culicinae aunque se presentan siete proteínas que se han encontrado solamente en el género Anopheles de Africa, A. gambie, de América, A. darlingi, como la antitrombina llamada anofelina (53). Otro aspecto importante de los trabajos de descripción de sialomas es la posibilidad de complementar estudios sobre la evolución de las moléculas presentes durante la hematofagia.
Pero también se ha informado reactividad cruzada entre diferentes especies de mosquitos, porque comparten antígenos lo cual se ha logrado evidenciar en estudios proteómicos y bioinformáticos; sin embargo, siempre se presentan antígenos específicos de cada mosquito (20,54). Estos últimos autores registran que las reacciones de inflamación inmediata y reacción retardada a la picadura de Cx. quinquefasciatus muestran una correlación estadísticamente significativa con las desarrolladas por la picadura de Ae. vexans; sin embargo, las reacciones mediadas por IgE son similares entre estas dos especies y también con Ae. aegypti.
Es claro que la desensibilización natural o las reacciones alérgicas a la picadura son generadas por proteínas presentes en la saliva de los mosquitos; se han identificado más de 30 proteínas en la saliva de Ae. aegypti (55,56) y otras especies de mosquitos de las cuales, varias son inmunogénicas para diferentes modelos animales y seres humanos (42,48,51,55) y muestran alto grado de conservación en diferentes poblaciones (56). Gracias a la secuenciación del genoma de algunas especies de mosquitos y a la aplicación de técnicas de bioinformática, biología molecular y proteómica, se ha logrado la identificación de un número mucho mayor de proteínas putativas en las glándulas salivares (más de 136 secuencias que codifican para proteínas secretadas), entre las secuencias identificadas, se encuentran algunas específicas de las glándulas salivares de hembras pero que no se conoce su función específica (57). Los análisis de expresión indican que alrededor del 90% de los genes de las glándulas salivares de Ae. aegypti no cambian o disminuyen su nivel de expresión luego de la ingestión de sangre (58).
A continuación se describen algunas moléculas que han sido caracterizadas, no solamente a nivel bioinformático, sino también en ensayos in vitro o in vivo.
Enzima nucleotidasas
Apirasa: inhibe la agregación de plaquetas al hidrolizar el ADP y ATP este sirve como una señal para la activación de los neutrófilos. De acuerdo con algunos estudios, las especies de mosquitos que son vectoras de parásitos o virus poseen mayor actividad de esta enzima en su saliva que las especies cercanas no vectoras (15); sin embargo, este no es un comportamiento universal para todas los géneros y especies (59).
Deaminasa de adenosina: se ha detectado actividad de esta enzima en la saliva de Cx. quinquefasciatus y Ae. aegypti, pero no en An. gambiae. Se cree que sus funciones en Ae. aegypti son remover la adenosina, (molécula asociada a la sensación de dolor), la inducción de la degranulación de mastocitos, y la producción de inosina, (molécula que inhibe la producción de citocinas inflamatorias) (60).
Fosfolipasa C: se ha descrito la actividad en la saliva de Cx. quinquefasciatus, y tiene acción específica contra el factor activador de plaquetas (FAP) (61).
Proteínas salivares de la familia D7
El primer gen D7 fue identificado en Ae. aegypti, y a partir de entonces, se han encontrado variables de este gen en casi todos los sialotranscriptomas de las especies de mosquitos estudiadas (55). Su expresión se da primordialmente en glándulas salivares de las hembras (57). Se ha demostrado que algunas proteínas de esta familia se unen e inhiben la acción de aminas biogénicas como la serotonina, histamina y norepinefrina, función que podría ser de utilidad durante la hematofagia (62).
Se ha postulado que el papel principal de las proteínas de la familia D7 es el secuestro de la serotonina liberada por los trombocitos en el sitio de la picadura, lo cual requiere que esta familia sea un componente mayor de la saliva (55).
Probablemente, la aparición de nuevas proteínas que previenen la agregación de plaquetas en el transcurso de la evolución, permitieron la diversificación de funciones de las proteínas D7, como en el caso de los anofelinos, en los cuales, se han identificado proteínas D7 con la capacidad de unirse a aminas diferentes o con función anti-bradiquinina (57,63,64). La hamadarina, es una proteína de esta familia, que ha sido identificada en An. stephensi (64).
Inhibidores de proteasas
En la saliva de Ae. aegypti, se han encontrado serpinas, una de las cuales participa en la inhibición del factor Xa de la cascada de coagulación (65,66) estas serpinas se expresan principalmente en glándulas salivares de hembras (57).
A nivel de secuencias genéticas, en Ae. aegypti se han descubierto homólogos de péptidos con el dominio Kazal descritos previamente para Ae. albopictus y que inhiben la trombina; estas secuencias se distribuyen en todos los tejidos de los mosquitos y pueden tener funciones housekeeping (57).
Sialoquinina: es una proteína presente específicamente en las glándulas salivares de las hembras; es un vasodilatador dependiente de endotelio, se caracterizó en Ae. aegypti (23, 67,68).
Diagnóstico y tratamiento
La mayoría de picaduras de mosquitos son indoloras (48), pero entre 0,2 – 2,5% de las personas pueden hacer alguna reacción moderada (69). En personas sensibilizadas con producción de niveles elevados de IgE e IgG específicos contra proteínas salivares de diferentes especies de mosquitos, se ha demostrado la desensibilización luego de la exposición continua a las picaduras (42).
Del mismo modo, la desensibilización natural puede producirse en la niñez o en la adolescencia; sin embargo, debido en parte a la escasez de preparaciones salivares de mosquito, las reacciones alérgicas a las picaduras no son diagnosticadas con frecuencia (48). Se han desarrollado inmunoensayos para medir anticuerpos IgE antimosquito en personas alérgicas y se ha demostrado que el ELISA de captura que usa como antígeno la saliva de los mosquitos es más específico y sensible que el que utiliza extracto de cuerpo entero para detectar pacientes con reacciones de hipersensibilidad local o severa; sin embargo, la máxima sensibilidad alcanzada en el ensayo fue del 56% (25).
A pesar que la anafilaxis producida por las picaduras de mosquitos puede ser más frecuente de lo que se puede deducir a partir de los reportes bibliográficos, no existen estudios que definan la prevalencia de esta condición en el mundo; de igual forma, se desconoce el posible papel de este tipo de anafilaxis como causa de muerte súbita o de anafilaxis idiopática. Debido a que la presencia de IgE no es específica de la anafilaxis, no existen pruebas de tamizaje para detectar la hipersensibilidad a la saliva de los mosquitos; el único test es el de exponer al paciente a la picadura de los insectos (49).
De acuerdo con los reportes bibliográficos, se han tratado pacientes con extractos de cuerpo entero de mosquitos para la hipersensibilidad a la picadura, principalmente para reacciones que no son de naturaleza anafiláctica (35,49,70-72). Una de las complicaciones puede ser la anafilaxis asociada a pruebas cutáneas que puede ocurrir incluso con extractos de cuerpo entero a muy bajas concentraciones (entre 1:1000 y 1:10000 por volumen); sin embargo, la mayoría de reportes son favorables al uso de este sistema para tratar la hipersensibilidad; aunque no se descartan las complicaciones causadas por el uso de extractos sin caracterizar (35).
A pesar de los resultados obtenidos con la inmunoterapia, no está claro si la desensibilización se logra por el tratamiento o simplemente se trata de la evolución natural de la condición, ya que la hipersensibilidad a la picadura generalmente decrece en reactividad con el paso del tiempo (49).
En la actualidad se dispone comercialmente de extractos de cuerpo entero para algunas especies de los géneros Aedes y Culex; sin embargo, no están completamente caracterizados y estandarizados, por lo tanto, varían tanto en calidad como en actividad biológica y algunos de ellos carecen de proteínas salivares; por el contrario, estos extractos contienen proteínas que no están presentes en la saliva de los mosquitos y pueden causar efectos adversos (73).
Como la obtención de saliva de los mosquitos es un proceso difícil y de poco rendimiento, en los últimos años se han utilizado métodos moleculares para producir alergenos puros, que han facilitado la investigación en el campo de la alergia a la picadura de los mosquitos y que en un futuro posiblemente servirán para desarrollar técnicas diagnósticas y de tratamiento estandarizadas y eficaces.
Los mayores avances al respecto se han dado con la saliva de Ae. aegypti, especie para la cual se han obtenido cuatro proteínas recombinantes (rAed a 1, 2, 3 y 4), entre las cuales se encuentran una apirasa de 68 kDa y una alfaglucosidasa de 67kDa (73-76). Estos alergenos recombinantes se han utilizado en estudios serológicos en los cuales se ha demostrado que los niveles de IgG e IgE específicas son mayores en individuos alérgicos (77).
Se ha evaluado el uso de rAed1 como método diagnóstico con sujetos alérgicos a la picadura de Ae. aegypti, comparando pruebas epicutáneas entre el antígeno recombinante y un extracto comercial de cuerpo entero del insecto; se observó que la antigenicidad y la actividad biológica del alergeno recombinante es similar a la de los extractos y presenta mayor especificidad; estos resultados son alentadores en la búsqueda de un método diagnóstico para la alergia a la picadura por Ae. aegypti (73).
Se han logrado adelantos con los alergenos Derp 1 y Der-f 1, que muestran actividad proteolítica que les permite penetrar la mucosa; por ser proteínas recombinantes, se supera el inconveniente de la diferencia en calidad y cantidad de alergeno que dan los extractos naturales; pero pueden tener sustancias indefinidas, baja cantidad de alergenos o inducir nuevas reacciones alérgicas, entre otros inconvenientes (78).
A pesar de los avances, aún no se ha avalado el uso de inmunoterapia basada en extractos de proteínas recombinantes, el primer tipo basado en alergeno silvestre y el segundo tipo en alergenos recombinantes que exhiben menor actividad alergénica que la proteína silvestre o alergenos híbridos (79).
Para que sea posible el uso de cualquiera de estas moléculas a nivel diagnóstico y clínico, se debe tener en cuenta que el producto esté constituido por sustancias bien estandarizadas, debe inducir tolerancia en células T efectoras específicas para el alergeno, suprimir la producción de IgE y promover la producción de anticuerpos IgG4 o IgA. Además, no puede producir efectos secundarios severos y debe ser bien tolerado y fácilmente administrado. Por otra parte, el tratamiento debe alcanzar éxito clínico y protección a largo plazo en un tiempo corto y usando dosis pequeñas. Adicionalmente, se deben tener en cuenta los marcadores biológicos e inmunológicos para evaluar el éxito clínico antes que sean identificables en el curso del tratamiento (80).
Discusión y conclusiones
Aunque se han publicado artículos que describen los sialotranscriptomas de varios insectos hematófagos, incluyendo los Culícidos, aún no se ha realizado la caracterización de los transcriptos en términos de la alergenicidad de sus productos. Entre otros aspectos, se debe estudiar la posibilidad de que la expresión de los transcriptos varíe con la edad fisiológica de las hembras o según la preferencia alimentaria de las especies en estudio. Por ejemplo, es posible que la expresión de proteínas sea diferente en especies de mosquitos cuya alimentación sea marcadamente antropofílica en comparación con especies preferentemente ornitofílicas o aquellas que se alimentan de fuentes sanguíneas diversas. Estas diferencias podrían ser el reflejo de mecanismos moleculares distintos que pue-den ser rastreados a nivel intra e interespecífico mediante el uso de técnicas de biología celular y molecular.
La secuenciación de genomas de varias especies de insectos hematófagos como Ae. aegypti, An. darlingi, An. gambiae, An. funestus (en curso), Cx. quinquefasciatus, Glossina morsitans (en curso), Rhodnius prolixus, Lutzomyia longipalpis (81) entre otros, ha facilitado la identificación de genes específicos de las glándulas salivares que regulan de alguna manera la función de la saliva de los mosquitos durante el proceso de la picadura, pero para muchas de ellas no se ha estabalecido la acción que tienen sobre el hospedero vertebrado, ni capacidad alergénica.
De acuerdo con estudios recientes de detección de productos de transcripción de Ae. aegypti mediante técnicas de hibridización in situ y RT-PCR, se detectó que las regiones proximales de los lóbulos laterales de las glándulas salivares tienen actividad transcripcional y que posiblemente, parte de la modulación de la composición de la saliva se da a nivel post-transcripcional (82).
Otra variable importante que debe ser tenida en cuenta en estos estudios es la influencia de las modificaciones metabólicas de los insectos hematófagos originadas por diferentes causas, tales como el desarrollo de la resistencia a los insecticidas o la infección con microrganismos transmitidos por vectores en el desarrollo de las reacciones alérgicas en el momento de la picadura. Puede suceder que la reacción alérgica se vea favorecida o inhibida por la presencia del agente infeccioso en las glándulas salivares o, desde otro punto de vista, las reacciones inmunológicas producidas en la piel del hospedero en el momento de la picadura pueden facilitar o impedir la entrada y supervivencia del mismo en el hospedero.
Por lo tanto, en este campo se abre una vasta posibilidad de estudios con el objetivo de identificar las moléculas implicadas en estos procesos e identificar las posibles reacciones cruzadas que puedan interferir con el desarrollo de técnicas de diagnóstico y tratamiento. Una herramienta de biología molecular que podría tener una amplia aplicación en este campo es la del silenciamiento genético por RNA de interferencia (RNAi).
Actualmente, se adelantan varios estudios que utilizan esta técnica para bloquear el desarrollo de agentes patógenos en el interior de sus vectores y con el objetivo de modificar genéticamente estos vectores para hacerlos refractarios o resistentes a la infección (83). Utilizando la técnica de RNAi sería posible caracterizar funcionalmente genes específicos de las glándulas salivares que afecten la composición de la saliva y por lo tanto, sería posible evaluar in vivo el efecto de la ausencia del producto génico específico sobre el desarrollo de la respuesta alérgica en el hospedero.
De este modo, sería posible el desarrollo de terapias de desensibilización utilizando moléculas sintéticas específicas, con lo que aumenta la eficacia del tratamiento y disminuye los efectos adversos producidos durante las terapias con extractos de glándulas salivares.
Referencias
1. Tempelis CH. Host feeding patterns of mosquitoes, with a review of advances in analysis of blood meals by serology. J Med Entomol. 1975; 11:635-53. [ Links ]
2. Gil LH, Alves FP, Zieler H, Salcedo JM, Durlacher RR, Cunha RP, et al. Seasonal malaria transission and variation of anopheline density in two distinct endemic areas in Brazilian Amazonia. J Med Entomol. 2003; 40:636-41. [ Links ]
3. Lines JD, Wilkes TJ, Lyimo EO. Human malaria infectiousness measured by age-specific sporozoite rates in Anopheles gambiae in Tanzania. Parasitology. 1991; 102:167-77. [ Links ]
4. Calhoun LM, Avery M, Jones L, Gunarto K, King R, Roberts J, et al. Combined sewage overflowes (CSO) are major urban breeding sites for Culex quinquefasciatus in Atlanta, Georgia. Am J Trop Med Hyg. 2007; 77:478-84. [ Links ]
5. Morais SA, Marrelli MT, Natal D. Aspectos da distribuição de Culex (Culex) quinquefasciatus Say (Diptera, Culicidae) na região do rio Pinheiros, na cidade de São Paulo, Estado de São Paulo, Brasil. Revista Brasileira de Entomologia. 2006; 50:413-18. [ Links ]
6. Sarmiento MI, Idrovo AJ, Restrepo M, Díaz MP, González A. Evaluación del Impacto de la Contaminación del Embalse del Muña Sobre la Salud Humana. Revista de Salud Pública Universidad Nacional de Colombia. 1999:159-71. [ Links ]
7. James A, Rossignol P. Mosquito salivary gland: parasitological and molecular aspects. Parasitology today. 1991; 7:267-71. [ Links ]
8. Rossignol PA. Parasite modification of mosquito probing behavior. Miscellaneous Publications Entomological Society of America. 1987; 68:25-8. [ Links ]
9. Pérez de León A, OToole D, Schmidtmann S, Ti-tus R, Tabachnick W. Insect blood feeding and the transmimssion of Arbiviruses and Vesiculoviruses. United States Animal Health Association Proceedings. 1997. http://www.usaha.org/speeches/insect97.html. Consultado 12/12/2010. [ Links ]
10. Ribeiro JMC. Role of mosquito saliva in blood vessel location. J Exp Biol. 1984;108:1-7. [ Links ]
11. Ribeiro JMC. Role of saliva in blood-feeding by arthropods. Ann Rev Entomol. 1987; 32:463-78. [ Links ]
12. Titus R, Ribero JMC. The role of vector saliva in transmission of arthropod-borne disease. Parasitol today. 1990; 6:157-60. [ Links ]
13. Ribeiro JMC. Blood feeding in mosquitoes: probing time and salivary gland anti-haemostatic activities in prepresentatives of three genera (Aedes, Anopheles, Culex). Med Vet Entomol. 2000; 14:142-48. [ Links ]
14. Pereira R, Dos Santos Malafronte R, Marinotti O. Salivary gland proteins of the mosquito Culex quinquefasciatus. Arch Insect Biochem Phys. 2000; 43: 9-15. [ Links ]
15. Cupp EW, Cupp MS, Ramberg FB. Salivary apyrase in African and New World vectors of Plasmodium species and its relationship to malaria transmission. Am J Trop Med Hyg. 1994; 50:235-40. [ Links ]
16. Moreira C, Mauro T, Marrelli S, Lima P, Marinotti O. Analysis of salivary gland proteins of the mosquito Anopheles darling (Díptera: Culicidae). J Med Entomol. 2001; 38:763-67. [ Links ]
17. Brummer-Korvenkontio HP, Lappalainen T, Reunala T, Palosuo T. Immunization of rabbits with mosquito bites: immunoblot analysis of IgG antimosquito antibodies in rabbit and man. Int Arch Allergy Immunol. 1990; 93:14-8. [ Links ]
18. Ramasamy M, Srikrishnaraj A, Wukoone S, Jesuthasan B, Ramasamy R. Host immunity to mosquitoes: effect of antimosquito antibodies an Anopheles tesellatus and Culex quinquefasciatus (Diptera: Culicidae). J Med Entomol. 1992; 29:934-38. [ Links ]
19. Peng Z, Yang M, Simons E. Measurement of mosquito Aedes vexans salivary gland specific IgE and IgG antibodies and the distribution of these antibodies in human sera. Ann Allergy Asthma immunol. 1995; 74: 259-64. [ Links ]
20. Peng Z, Simmons FER. Comparison of proteins IgE and IgG binding antigens and skin reactivity in commercial and laboratory-made mosquito extracts. Ann Allergy Asthma Immunol. 1996; 131:1730. [ Links ]
21. Jariyapan N, Harnnoi T. Preliminary study of salivary gland proteins of the mosquito Aedes togoi (Theobald). Chiang Mai Med Bull. 2002; 41:21-8. [ Links ]
22. Ribeiro JMC, Francischetti IMB. Role of Arthropod saliva in blood feeding: Sialome and Post-Sialome Perspectives. Annu Rev Entomol. 2003; 48:73-88. [ Links ]
23. Champagne D, Ribeiro JMC. Sialokinins I and II: Two salivary tachykinins from the Yellow Fever mosquito, Aedes aegypti. Proc Natl Acad Sci (USA). 1994; 91:138-42. [ Links ]
24. Stark K, James A. Salivary gland anticoagulants in Culicinae and Anophelinae mosquitoes (Diptera: Culicidae). J Med Entomol. 1996; 33:645-50. [ Links ]
25. Peng Z, Simons FER. Advances in mosquito allergy. Current Opinion in Allergy and Clinical Immunology. 2007; 7:350-54. [ Links ]
26. Akdis CA. Allergy and hypersensitivity mechanisms of allergic disease. Curr Opin Immunol. 2006; 18:718-26. [ Links ]
27. Larche M, Akdis CA, Valenta R. Immunological mechanisms of allergen-specific immunotherapy. Nat Rev Immunol. 2006; 6:761-71. [ Links ]
28. Romagnani S. Immunologic influences on allergy and the TH1/TH2 balance. J Allergy Clin Immunol. 2004; 113:395-40. [ Links ]
29. Frazier CA. Insect allergy and toxic reactions to insects and other arthropods. Warren H. Green Inc. St Louis, Missouri, Estados Unidos. 1969. [ Links ]
30. Brummer-Korvenkontio H. Mosquito bite allergy: characterization of saliva antigens and ssaliva specific antibodies. Helsinki, Finlandia. Tesis. 1996. [ Links ]
31. Sánchez M, de la Vega W. Manual Latinoamericano de Alergia e Inmunología. Editora Guadalupe, Bogotá, Colombia. 1987. [ Links ]
31. Peng Z, Yang M, Simons FER. Immunological mechanisms in mosquito allergy: Correlation of skin reactions with specific IgE and IgG antibodies and lymphocyte proliferation response to mosquito antigens. Ann Allergy Asthma Immunol. 1996; 77:238-44. [ Links ]
32. Weed R. Exaggerated delayed hypersensitivity to mosquito bites in chronic lymphocytic leukemia. 1965. Blood. 1965; 26:257-68. [ Links ]
33. Garcia-Ortiz JC, Cosmes M. Anaphylactic shock following a mosquito Bite. Med. Clin., Barcelona. 1994; 102:157. [ Links ]
34. McCormick DK, Salata K, Harshey J, Carpenter G, Engler R. Mosquito bite anaphylactic: immunotherapy with whole body extracts. Ann Allergy Asthma Immunol. 1995; 74:39-44. [ Links ]
35. Demeure CE, Brahimi K, Hacini F, Marchand F, Péronet R, Huerre M, et al. Anopheles Mosquito Bites Activate Cutaneous Mast Cells Leading to a Local Inflammatory Response and Lymph Node Hyperplasia. J. Immunol. 2005; 174:3932-40. [ Links ]
36. Depinay N, Hacini T, Beghdadi W, Peronet R, Mécheri S. Mast cell-dependent down-regulation of antigen-specific immune responses by mosquito bites. T J Immunol. 2006; 176:4141-6. [ Links ]
37. Donovan MJ, Messmore AS, Scrafford DA, Sacks DL, Karmhavi S, McDowell MA. Uninfected mosquito bites confer protection against infection with malaria parasites. Infect Immun. 2007; 75:2523-30. [ Links ]
38. Zeidner NS, Higgs S, Happ CM, Beaty BJ, Miller BR. Mosquito feeding modulates Th1 and Th2 cytokines in flavivirus susceptible mice: an effect mimicked by injection of sialokinins, but not demonstrated in flavivirus resistant mice. Parasit Immunol. 1999; 21:35-44. [ Links ]
39. Bissonnette EY, Rossignol PA, Befus AD. Extracts of mosquito salivary gland inhibit factor alpha release from mast cells. Parasite Immunol. 1993; 15:27-33. [ Links ]
40. Cross ML, Cupp EW, Enríquez FJ. Differential modulation of murine cellular immune responses by salivary gland extract of Aedes aegypti. Am J Trop Med Hyg. 1994; 51:690-6. [ Links ]
41. Peng Z, Li H, Simons FER. Immunoblot analysis of salivary allergens in 10 mosquito species with world wide distribution and the human IgE responses to these allergens. J Allergy Clin Immunol. 1998; 101:498-505; erratum 746. [ Links ]
42. Ailus K, Palosuo T, Brummer-Korvenkontio M, Rantanen T, Reunala T. Demonstration of antibodies to mosquito antigens in man by immunodiffusion and ELISA. Int Arch Allergy Appl Immunol. 1985; 78: 375-79. [ Links ]
43. Konishi E. Distribution of immunoglobulin G and E antibody levels to salivary gland extracts of Aedes albopictus (Diptera: Culicidae) in several age groups in Japanese population. J Med Entomol. 1990; 27:519-22. [ Links ]
44. Peng Z, Simons FER. Cross reactivity of skin and serum specific IgE responses and allergen analysis for three mosquito species IgE and IgG levels in children. J Allergy Clin Immunol. 1997; 101:498-505. [ Links ]
45. Cornelie S, Remoue F, Doucoure S, N´Diaye T, Sauvage F-X, Boulanger D,et al. An insight into immunogenic salivary proteins of Anopheles gambiae in African children. Malaria Journal. 2007; 6:75 doi:10.1186/1475-2875-6-75. [ Links ]
46. Oka K. Correlation of Aedes albopictus bite reaction with IgE antibody assay and lymphocyte transformation test to mosquito salivary antigens. J Dermatol. 1989; 16:341-47. [ Links ]
47. Peng Z, Simons FER. Mosquito Allergy: Immune Mechanisms and recombinant salivary allergens. Int Arch Allergy Immunol. 2004; 133:198-209. [ Links ]
48. Engler R. Mosquito bite pathogenesis in necrotic skin reactors. Curr Opin Allergy Clin Immunol. 2001; 1:349-52. [ Links ]
49. Charlab R, Valenzuela J, Rowton E, Ribeiro J. Toward an understanding of the biochemical and pharmacological complexity of the saliva of a hematophagous sand fly Lutzomyia longipalpis. Proc Natl Acad Sci USA. 1999; 96: 15155-60. [ Links ]
50. Salazar M. Caracterización de los antígenos presentes en extractos provenientes del mosquito Culex quinquefasciatus (Diptera:Culicidae) y de la respuesta humoral del tipo IgG en conejos inmunizados. Trabajo de grado. Departamento de Biología, Facultad de Ciencias, Universidad Nacional de Colombia, sede Bogotá. 2002. [ Links ]
51. Al-Ahdal M, Al-Hussasin K, Thorogood R, Reilly H, Wilson J. Protein constituents of mosquito saliva: studies on Culex molestus. J Trop Med Hyg. 1990; 93:98-105. [ Links ]
52. Calvo E, MyPham V, Marinotti O, Andersen JF, Ribeiro JMC. The salivary gland transcriptome of the neotropical malaria vector Anopheles darlingi reveals accelerated evolution of genes relevant to hematophagy. BMC Genomics, 2009; 10:57 doi:10.1186/ 1471-2164-10-57. [ Links ]
53. Killby V, Silverman P. Hypersensitive reaction in man to specific mosquito bites. Am J Trop Med Hyg. 1967; 16:374-80. [ Links ]
54. Valenzuela JG, Charlab R, Gonzalez EC, Miran-da-Santos IKF, Marinotti O, Francischetti IM, et al. The D7 family of salivary proteins in blood sucking Diptera. Insect Mol Biol. 2002; 11:149-55. [ Links ]
55. Almeras L, Fontaine A, Belghazi M, Bourdon S, Boucomont-Chapeaublanc E, Orlandi-Pradines E, et al. Salivary Gland Protein Repertoire from Aedes aegypti Mosquitoes. Vector Borne Zoonotic Dis. 2009; Oct 30. [Epub ahead of print]. [ Links ]
56. Ribeiro JMC, Arca B, Lombardo F, Calvo E, MyPhan V, Chandra PK, et al. An annotated catalogue of salivary gland transcripts in the adult female mosquito, Aedes ægypti. BMC Genomics. 2007; 8:6 doi:10.1186/1471-2164-8-6. [ Links ]
57. Thangamani S, Wikel S. Differential expression of Aedes. Aegypti salivary transcriptome upon blood feeding. Parasites and Vectors. 2009; 2:34. [ Links ]
58. Reno HE, Novak RJ. Characterization of Apyrase-Like Activity in Ochlerotatus triseriatus, Ochlerotatus hendersoni, and Aedes aegypti. Am J Trop Med Hyg. 2005; 73:541-45. [ Links ]
59. Ribeiro JMC, Charlab R, Valenzuela JG. The Salivary Adenosine Deaminase Activity of the Mosquitoes Culex quinquefasciatus and Aedes aegypti. J Exp Biol. 2001; 204:2001-10. [ Links ]
60. Ribeiro JMC, Francischetti IMB. Platelet-activatingfactor-hydrolyzing phospholipase C in the salivary glands and saliva of the mosquito Culex quinquefasciatus. J Exp Biol. 2001; 204:3887-94. [ Links ]
61. Calvo E, Mans BJ, Andersen JF, Ribeiro JM. Function and evolution of a mosquito salivary protein family. J Biol Chem. 2006; 281:1935-42. [ Links ]
62. Rai KS, Black WC IV. Mosquito genomes: Structure, organization, and evolution. Adv Genet. 1999; 41:1-33. [ Links ]
63. Isawa H, Yuda M, Orito Y, Chinzei Y. A Mosquito Salivary Protein Inhibits Activation of the Plasma Contact System by Binding to Factor XII and High Molecular Weight Kininogen. J Biol Chemis. 2002; 277: 27651-8. [ Links ]
64. Stark KR, James AA. A factor Xa-directed anticoagulant from the salivary glands of the yellow fever mosquito Aedes aegypti. Exp Parasitol. 1995; 81:321-31. [ Links ]
65. Stark KR, James AA. Isolation and characterization of the gene encoding a novel factor Xa-directed anticoagulant from the yellow fever mosquito, Aedes aegypti. J Biol Chem. 1998; 273:20802-9. [ Links ]
66. Ribeiro JMC. Characterization of vasodilator from the salivary glands of the yellow fever mosquito, Aedes aegypti. J Exp Biol. 1992; 165:61-71. [ Links ]
67. Beerntsen BT, Champagne DE, Coleman JL, Campos YA, James AA. Characterization of the Sialokinin I gene encoding the salivary vasodilator of the yellow fever mosquito, Aedes aegypti. Insect Mol Biol. 1999;8: 459-67. [ Links ]
68. Arias-Cruz A, Avitia-Valenzuela E, González-Díaz SN. Galindo-Rodríguez G. Epidemiology of mosquito bite allergy in the centre of allergy and clinical Immunology of Monterrey, Mexico. J Allergy Clin Immunol. 2006; 117:128. [ Links ]
69. Galindo PA, Gomez E, Borja J, Feo F, García F, Lombardero M, et al. Mosquito bite hypersensitivity. Allergol Immunopathol. 1998; 26:251-54. [ Links ]
70. Hassoun S, Drouet M, Sabbah A. Anaphylaxis caused by a mosquito: 2 case reports. Allerg Immunol. 1999; 31:285-7. [ Links ]
71. Benain-Pinto C, Fassrainer M. Intradermal immunotherapy in children with severe skin inflammatory reactions to Aedes aegypti and Culex quinquefasciatus mosquito bites resulted in significant reduction in morbidity of subsequent natural bites. Int J Dermatol. 1990; 29:600-1. [ Links ]
72. Peng Z, Xu W, James AA, Lam H, Sun D, Cheng L. Expression, purification, characterization, and clinical relevance of rAed a 1: a 68 kDa recombinant mosquito Aedes aegypti salivary allergen. Inter Immunol. 2001; 13:1445-52. [ Links ]
73. Peng Z, Xu W, Lam H, Cheng L, James AA, Simons FE. A new recombinant mosquito salivary allergen, rAed a 2: allergenicity, clinical relevance, and crossreactivity. Allergy. 2006; 61:485-90. [ Links ]
74. Xu W, Peng Z, Simons FER. Isolation of a cDNA encoding a 30 kDa IgE-binding protein of mosquito Aedes aegypti saliva. J Allergy Clin Immunol. 1998;101: S203. [abstract]. [ Links ]
75. Li C, Beckett AN, James AA, Simmons FER, Arthur G, Peng Z. Diagnosis of mosquito allergy: role of a new 67 kDa recombinant Aedes aegypti salivary allergen rAed a 4 [abstract]. Allergy Clin Immunol Int J World Allergy Org. 2003; (Suppl):195. [ Links ]
76. Beckett AN, Sun W, Simons FER, Ma Y, Peng Z. Role of recombinant mosquito salivary allergens in the diagnosis of individuals with allergic reactions to mosquito bites. J Allergy Clin Immunol. 2004;113:S74. [ Links ]
77. Kraft D, Ferreira F, Ebner G, Valenta R, Breiteneder H, Susani M, et al. Recombinante allergen the future of the diagnosis anda treatment of atopic allergy. Allergy. 1998; 53:62- 6. [ Links ]
78. Valenta R, Niederberger V. Recombinant allergens for immunotherapy. J. Aller Clin Immunol. 2007; 9: 26-30. [ Links ]
79. Mübeccel A, Cezmi A. Mechanisms of allergen-specific immunotherapy. J Allergy Clin Immunol. 2007; 119:780-9. [ Links ]
80. NCBI Entrez Genome Project. URL: http://www.ncbi.nlm.nih.gov/genomes/leuks.cgi. Consulta: 15 de mayo de 2011. [ Links ]
81. Juhn J, Naeem-Ullah U, Guedes B, Majid A, Coleman J, Pimenta P, et al. Spatial mapping of gene expression in the salivary glands of the dengue vector mosquito, Aedes aegypti. Parasit Vectors. 2011; 4:1-13. [ Links ]
82. Wu X, Hong H, Yue J, Wu Y, Li X, Jiang L, Li L, Li Q, Gao G, Yang X. Inhibitory effect of small interfering RNA on dengue virus replication in mosquito cells. Virol J. 2010;14:270-7. [ Links ]
