










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkIdeas y Valores
Print version ISSN 0120-0062
Ideas y Valores vol.59 no.145 Bogotá Jan./Apr. 2011
EL ESTATUS FENOMENOLÓGICO DE LA TEORÍA DE LA SELECCIÓN NATURAL*
The Phenomenological Status of the Theory of Natural Selection
SANTIAGO GINNOBILI
Conicet / Universidad de Buenos Aires
Universidad Nacional de Quilmes - Argentina
santi75@gmail.com
Artículo recibido: 4 de febrero de 2010; aceptado: 23 de febrero de 2010.
Resumen
Algunos han señalado la naturaleza fenomenológica -en el sentido clásico de que no propone nuevos términos teóricos- de la teoría de la selección natural. Como otros han sostenido que una característica de las teorías explicativas es que enriquecen conceptualmente su ámbito de aplicación, podría señalarse, consecuentemente, que la teoría en cuestión tiene algún déficit en su capacidad explicativa. Se aborda esta cuestión presentando una propuesta de reconstrucción informal de la teoría de la selección natural darwiniana por medio de la herramienta metateórica estructuralista. El objetivo es mostrar cómo, pese a que todos los conceptos de la teoría son no teóricos en el sentido estructuralista, tal teoría propone al menos un concepto explicativo, que se puede denominar "aptitud".
Palabras clave: C. Darwin, aptitud, estructuralismo, fenomenología, selección natural.
Abstract
Some scholars have pointed out the phenomenological character of the theory of natural selection, in the sense that it does not propose new theoretical terms. Since others have held that one of the characteristics of explanatory theories is that they conceptually enrich their field of application, it could be said that the theory in question is lacking with respect to its explanatory capacity. The article addresses this issue by proposing an informal reconstruction of the Darwinian theory of natural selection using structuralist meta-theoretical tools. The objective is to show that the theory suggests at least one explanatory concept, which could be called "aptitude", although all of its concepts are non-theoretical in the structuralist sense.
Key words: C. Darwin, aptitude, structuralism, phenomenology, natural selection.
Aunque es una de las teorías científicas más fáciles de entender, la teoría de la selección natural (en adelante TSN) escapa a los intentos de elucidación. Entre las diversas reconstrucciones realizadas no se ha acordado ni siquiera cuáles son sus conceptos fundamentales. Parece razonable, frente a esta situación, intentar elucidar esta teoría en su origen. Como la teoría nunca es enunciada de manera explícita por Darwin, es necesario abstraerla de sus diferentes aplicaciones. Al hacer esto, puede notarse que son al menos tres los conceptos fundamentales que conforman su ley fundamental: el de adecuación (un rasgo que cumple una función con cierta efectividad), el de éxito reproductivo diferencial y el concepto que une el rasgo adecuado con el éxito reproductivo diferencial, que he llamado "aptitud".1
Algunos han considerado que TSN es fenomenológica, en el sentido de que no propone, utilizando terminología de la concepción heredada, conceptos teóricos −o no observacionales− (Klimovsky 1994, Putnam 1962). Es conocida, sin embargo, la infinidad de problemas señalados en la distinción clásica teórico/observacional. En consecuencia, es necesario discutir esta cuestión con alguna posición más sofisticada. Asumiré la distinción T-teórico / T-no teórico propuesta por primera vez por Sneed, e incluida dentro del programa metateórico estructuralista (Balzer, Moulines & Sneed 1987). Según esta propuesta, un concepto es teórico en la teoría T si es necesario acudir a T para determinarlo, y es no teórico en T si puede ser determinado a partir de otras teorías distintas a T.
Por otra parte, se ha señalado que las teorías que no proponen términos teóricos (en el sentido estructuralista) no son explicativas, o no lo son tanto como las que sí lo hacen. Bajo esta concepción, explicar un sistema empírico dado con la teoría T consiste en expandirlo teóricamente (es decir, con conceptos teóricos para la teoría T), de modo que se vuelva modelo de esa teoría (Bartelborth 1996, 2002, Diez 2002, Forge 2002, Moulines 2005). El objetivo de este trabajo es mostrar, en contra de la concepción recién comentada que asume que toda expansión conceptual se realiza a través de conceptos T-teóricos, que, si bien TSN carece de términos TSN-teóricos, se puede considerar que propone un nuevo concepto explicativo, el de aptitud, y, por lo tanto, puede considerarse legítimamente explicativa.
Como corolario metateórico general de esta discusión, intentaré mostrar que la distinción estructuralista entre conceptos T-teóricos y T-no teóricos es fructífera para determinar aquellos conceptos que, porque pueden ser determinados con independencia de la teoría, permiten contrastarla (distinción para la que fue originalmente planteada), pero que no es igualmente efectiva para distinguir entre conceptos T-explicativos y T-no explicativos, entendiendo por un concepto T-explicativo un concepto con el que T enriquece la descripción del fenómeno previa a T y del cual se quiere dar cuenta con T, es decir, un concepto que T utiliza para explicar.
1. La ley fundamental de la teoría de la selección natural
Comenzaré exponiendo los conceptos fundamentales de TSN y lo que se pretende explicar con esta, y extraeré los conceptos fundamentales de TSN a partir de casos de aplicación del mismo Darwin.
Como es bien sabido, lo que Darwin quiere explicar con TSN es cierta adecuación o ajuste de los organismos al medio ambiente. Por ejemplo:
La jirafa, con su gran estatura, su muy largo cuello, patas delanteras, cabeza y lengua, tiene su estructura bellamente adaptada para comer en las ramas más altas de los árboles. Puede por eso obtener comida fuera del alcance de otros ungulados que habitan el mismo lugar; y esto debe ser una gran ventaja durante periodos de escasez. (Darwin 1872 178)
La forma en que Darwin explica la fijación de este rasgo en la población de jirafas es la siguiente:
En la naturaleza, en el origen de la jirafa, los individuos que comiesen más alto y que pudiesen durante los periodos de escasez alcanzar aunque sea una pulgada o dos por sobre los otros, serían frecuentemente preservados [...]. Que los individuos de la misma especie muchas veces difieren un poco en la longitud relativa de todas sus partes puede comprobarse en muchas obras de historia natural en las que se dan medidas cuidadosas. Estas pequeñas diferencias en las proporciones, debidas a las leyes de crecimiento o variación, no tienen la menor importancia ni utilidad en la mayor parte de las especies. Pero en el origen de la jirafa debe haber sido diferente, considerando sus probables hábitos de vida; pues aquellos individuos que tuviesen alguna parte o varias partes de su cuerpo un poco más alargadas de lo corriente hubieron en general de sobrevivir. Se habrán cruzado y habrán dejado descendencia que habrá heredado las mismas peculiaridades corpóreas, o la tendencia a variar de nuevo en la misma manera, mientras que los individuos menos favorecidos en los mismos aspectos habrán sido más propensos a perecer. (1872 178)
Si consideramos, con los estructuralistas, que la ley fundamental de TSN es aquel enunciado en el que aparecen los conceptos fundamentales de TSN relacionados (Balzer, Moulines & Sneed 19), podemos extraer una instanciación de TSN a partir de la explicación citada:
• Las jirafas con cuello, patas delanteras, cabeza y lengua de mayor longitud son más efectivas al alimentarse de las ramas más altas de los árboles, con lo que mejoran su supervivencia y, en consecuencia, su éxito reproductivo diferencial.
Nada en este enunciado parece superfluo. Si quitamos alguna de sus partes, la explicación brindada quedaría trunca. Si se quita la función dada al rasgo por el organismo, la de alcanzar las ramas altas de los árboles, no sabríamos por qué tal rasgo podría mejorar la supervivencia. Podría mejorarla, por ejemplo, permitiendo asustar a posibles predadores, lo que constituiría una explicación alternativa y competidora de la ofrecida por Darwin. Si quitamos la mejora en la supervivencia, quedaría indeterminada la relación que hay entre el rasgo y el éxito reproductivo. No sabríamos si se trata de un caso de selección sexual, por ejemplo, siendo el rasgo y su capacidad atractivos en algún sentido a las hembras de la misma especie.
Por abstracción de este enunciado, nos vamos acercando a lo que considero que es la ley fundamental de TSN:
• Los individuos con rasgos que cumplen con mayor efectividad cierta función2 mejoran su supervivencia y mejoran su éxito en la reproducción diferencial.
La ley fundamental de TSN tendría al menos tres componentes:
a. El rasgo que cumple de manera más adecuada una función.
b. El éxito reproductivo diferencial, que tiene que ver con alguna medida de la cantidad de descendencia fértil dejada por los organismos.
c. La conexión entre el rasgo adecuado y el éxito reproductivo, que llamaré "aptitud". En este caso se debe a una mejora en la supervivencia. Como veremos, y esto es central en este trabajo, no siempre es así.
2. Diferentes interpretaciones de aptitud
Según Darwin, la razón fundamental para aceptar la teoría de la selección natural es su capacidad unificadora. Autores contemporáneos, como Kitcher (1993), están de acuerdo con esta idea. Sin embargo, esta capacidad unificadora no es tan patente como, por ejemplo, sí lo es en el caso de la mecánica clásica. Son conocidas las diferentes leyes que fueron subsumidas por Newton bajo el segundo principio de su mecánica.
Mi intención es mostrar ahora la diversidad de explicaciones que surgen al interpretar de manera diferente el concepto de aptitud. Esto permitirá, por un lado, entender mejor la extensión y naturaleza de tal concepto, y, por el otro, entender la capacidad unificadora de TSN.
Creo que, del mismo modo en que Endler trata la cuestión al respecto de la teoría de la selección natural actual (1986 9, 1992), TSN tiene dos grandes ramas de especificación: la ley especial de la selección sexual y la ley especial de la selección no sexual o de la selección natural en sentido restringido. Esta distinción es planteada por Darwin del siguiente modo:
Esto me lleva a decir algunas palabras acerca de lo que yo he llamado Selección Sexual. Esta forma de selección depende, no de la lucha por la existencia en relación con otros seres orgánicos o las condiciones externas, sino de la lucha entre individuos de un mismo sexo, generalmente los machos, por la posesión del otro sexo. (1872 69)
2.1. Selección sexual
En la selección sexual, la aptitud consiste en la capacidad de conseguir pareja. La ley especial de la selección sexual surgiría, entonces, de la especificación del concepto de aptitud como capacidad de conseguir organismos del otro sexo para cruzarse, lo que llamaré "capacidad de emparejarse".
La selección sexual puede funcionar, o bien con base en la capacidad de los organismos del mismo sexo para pelear literalmente o de manera ritualizada por organismos del otro sexo, y esta selección promueve la existencia de rasgos como los espolones del gallo, cuya función es vencer a los contendientes, o bien con base en la capacidad de los organismos de un sexo para atraer a los del otro, y en este caso promueve rasgos que pueden resultar atrayentes a las hembras, como la cola del pavo real.
2.1.1. Capacidad de emparejarse luchando con organismos del mismo sexo
Darwin sostiene:
Generalmente, los machos más vigorosos, los que están mejor adecuados a su situación en la naturaleza, dejarán más descendencia, pero en muchos casos la victoria depende no tanto del vigor natural, como de la posesión de armas especiales limitadas al sexo masculino. Un ciervo sin cuernos, un gallo sin espolones, habrían de tener pocas probabilidades de dejar numerosa descendencia. La selección sexual, dejando siempre criar al vencedor, pudo, seguramente, dar valor indomable, longitud a los espolones, fuerza al ala para empujar la pata armada de espolón. (1859 88)
El enunciado legaliforme supuesto sería:
• Los gallos de espolones más efectivos para luchar con gallos del mismo sexo tienden a emparejarse más, con lo cual mejoran, en consecuencia, su éxito en la reproducción diferencial.
2.1.2. Capacidad de emparejarse atrayendo a organismos del otro sexo
En la siguiente cita podemos encontrar el segundo tipo de selección sexual:
Entre las aves, la contienda es muchas veces de carácter más pacífico. [...] los machos sucesivamente despliegan con el más minucioso cuidado y exhiben de la mejor manera su espléndido plumaje; además ejecutan extraños movimientos ante las hembras, que, asistiendo como espectadoras, escogen al fin al compañero más atractivo. (Darwin 1859 88-89)
El enunciado legaliforme supuesto ahora es:
• Las aves de plumaje más atractivo a las hembras tienden a emparejarse más, con lo cual mejoran, en consecuencia, su éxito en la reproducción diferencial.
2.2. Selección no sexual o restringida
Darwin suele contraponer la selección sexual a la selección natural basada en la lucha por la existencia. Podríamos entonces obtener la otra rama de especialización determinando la aptitud como la capacidad de supervivencia. Pero recordemos que la expresión "lucha por la existencia" es utilizada por Darwin de manera metafórica, pues no sólo incluye la lucha por la supervivencia, sino también, por ejemplo, el incremento de fecundidad en las flores debido a la mayor atracción que estas ejercen sobre ciertos insectos (Darwin 1859 62-63). Si entendemos "lucha por la existencia" de este modo, la caracterización de la selección no sexual como "la que depende de la lucha por la existencia con otros seres orgánicos o las condiciones de vida", dado que los seres orgánicos del mismo sexo también son seres orgánicos con los que se compite en la lucha por la existencia, parece referirse a cualquier tipo de lucha que no sea por conseguir pareja. Esto puede dar cierto aire de artificialidad a la agrupación de diferentes leyes especiales bajo la ley especial de selección no sexual, pues bajo esta se agruparían, como veremos, el incremento de la aptitud como incremento en la fecundidad, el incremento de aptitud debido al incremento en la capacidad de supervivencia, etc. ¿Por qué no poner estas leyes a la misma altura de la selección sexual y eliminar la ley especial de selección no sexual por artificial? Voy a mantener la ley de selección no sexual para poder conservar la expresión "selección natural restringida", útil a la hora de interpretar ciertos pasajes del Origen. La selección restringida o no sexual es a lo que muchas veces Darwin se refiere con "selección natural".
2.2.1. Supervivencia
La primera especialización de esta línea es la que surge de determinar la aptitud como la capacidad de supervivencia. Este es el mecanismo utilizado en el Origen. Cada vez que Darwin tiene que exponer el funcionamiento de la selección natural, ofrece como ejemplar este caso. Veamos un caso particular de este tipo de aplicación:
Cuando vemos [...] blanca en invierno la perdiz alpina y del color de los brezos la perdiz de Escocia, hemos de creer que estos colores son de utilidad a estas [...] aves para librarse de peligros. Los lagopus, especie de perdices, si no fuesen destruidos en algún periodo de su vida, aumentarían hasta ser innumerables, pero se sabe que sufren mucho por las aves de rapiña, y los halcones se dirigen a sus presas por el sentido de la vista [...]. Por consiguiente, la selección natural pudo ser eficaz para dar el color conveniente a cada especie de lagopus, y en conservar este color justo o constante una vez adquirido. (Darwin 1859 84-85)
El enunciado legaliforme supuesto sería:
• Las perdices cuyo color se mimetiza con su entorno a los ojos de sus predadores tienden a mejorar su supervivencia y, en consecuencia, su éxito reproductivo diferencial.
2.2.2. Fecundidad
Por el carácter informal de mi presentación, y el hecho de usar conceptos en su sentido habitual, no siempre unívoco, en las presentaciones de las leyes no queda claro qué ocurre con el caso de las flores que atraen insectos para polinizarse. ¿Podemos considerar esto un caso de selección sexual o no? Podría ser un caso de selección sexual, si entendemos "la capacidad entre distintos tipos de organismos de entablar exitosamente luchas con los organismos del mismo sexo para emparejarse" de manera lo suficientemente general. De todos modos, prefiero entender la expresión anterior del modo más literal posible; además, el mismo Darwin considera a este un caso de lo que yo llamé "selección natural restringida" y no de selección sexual (cf. Darwin 1859 91-95). Así, podríamos incluir una nueva ley especial que surgiera de especificar aptitud como fecundidad. Darwin aplica la selección natural también de este modo:
Las plantas que produjesen flores con las glándulas y nectarios mayores y que segregasen más néctar serían las visitadas con mayor frecuencia por insectos y las más frecuentemente cruzadas, y de este modo, a la larga, adquirirían ventaja y formarían una variedad local. (Darwin 1859 92)
El enunciado legaliforme supuesto sería:
• Las plantas que producen flores más atractivas para los insectos tienden a mejorar su fecundidad y, en consecuencia, su éxito en la reproducción diferencial.
2.2.3. Selección artificial
En un texto de próxima aparición he defendido que la selección artificial se trata de un caso de la selección natural y, más particularmente, que se trata de una contrastación experimental de TSN. Puesto que parece posible realizar experimentos controlados acerca de cualquiera de las leyes arriba señaladas, carecería de sentido introducir una ley especial de la selección artificial. Sin embargo, creo que Darwin no sólo considera la selección artificial como una versión experimental de la selección natural, sino como un mecanismo particular que funciona en la naturaleza. En cuanto tal, debería ser incluido en la red teórica. La selección artificial es dividida por Darwin en dos procesos: la selección metodológica, que es la realizada por los criadores que de manera intencional buscan la modificación de las razas (Darwin 1859 30), y la selección inconsciente, que es la selección realizada por individuos que no son conscientes de que están modificando las razas en una dirección, y que no necesariamente se encargan de realizar los cruces entre los organismos seleccionados, sino que simplemente los cuidan más que a los otros (id. 39).
Por dudar de la distinción entre lo artificial y lo natural, y por rechazar las consecuencias prácticas de esta distinción, yo tendería a disgregar los casos que caen bajo estos procesos en las leyes ya establecidas. Así, podríamos incluir la selección metódica bajo la ley especial de mayor fecundidad (la paloma seleccionada por el criador mejora su aptitud al portar un rasgo que resultó atrayente al cuidador, del mismo modo que la flor resultó atrayente para el insecto que la poliniza); la selección inconsciente en la que se escogen las parejas, pero sin tener la meta de mejorar la raza, se podría incluir también bajo esta ley, y la selección inconsciente en la que simplemente se cuida de los peligros a las variedades más útiles se podría incluir bajo la ley especial de la supervivencia. Sin embargo, considero que, por el lugar que ocupan tales procesos en la argumentación de Darwin, es mejor introducir leyes especiales que los representen. Así, podemos incluir la selección artificial como una especialización de la selección natural restringida.
2.2.4. Selección de grupo
Un tipo de aplicación de TSN particularmente interesante en el Origen es el de la selección de grupo, ampliamente debatida en los textos actuales, pero que Darwin aceptaba sin titubear:
En ciertas circunstancias, diferencias individuales en la curvatura o longitud de la lengua, etc., demasiado ligeras para ser apreciadas por nosotros, podrían servirle a una abeja u otro insecto de modo que ciertos individuos fuesen capaces de obtener su alimento más rápidamente que otros, y así, las comunidades a que ellos perteneciesen prosperarían y darían muchos enjambres que heredarían las mismas cualidades. (Darwin 1872 74-75)
Podría expresarse el enunciado legaliforme supuesto de la siguiente forma:
• Las abejas con la curvatura o longitud de la lengua más efectiva para recolectar néctar de ciertas flores mejoran el rendimiento de la comunidad a la que pertenecen y, en consecuencia, el éxito reproductivo diferencial de tal comunidad.3
2.2.5. Otras posibles especializaciones
De ningún modo estas pretenden agotar el tipo de especializaciones posibles de TSN, ni siquiera aquellas que podemos encontrar en el mismo Darwin.
Con las leyes propuestas, por ejemplo, no se puede explicar la capacidad de ciertos árboles de esparcir sus semillas a través de frutos que atraen a ciertos pájaros. Creo que podríamos introducir una nueva ley especial que incluya este caso y también las diferentes adecuaciones de semillas que permiten, por ejemplo, aprovechar al viento como medio de dispersión. Estos ejemplos son muy trabajados por Darwin a lo largo del Origen, sobre todo en el capítulo dedicado a la distribución geográfica. Esta ley surgiría al especificar la aptitud como 'la capacidad de dispersar sus simientes'.
Otra posible especialización podría constituirla el principio de divergencia (Darwin 1859 112): aptitud, en este caso, podría interpretarse a partir de la divergencia del resto de la población.
3. La red teórica de la teoría de la selección natural
La ley fundamental de TSN podría tener la siguiente forma:
• Los organismos que portan un rasgo que cumple más efectivamente una funcións tienden a mejorar su capacidad de emparejarse, su supervivencia, su fecundidad, su utilidad a humanos y/o el rendimiento de la comunidad a la que pertenecen, y tienden a mejorar, en consecuencia, su éxito reproductivo diferencial o el éxito reproductivo diferencial de la comunidad a la que pertenecen.
Si llamamos "aptitud" a la conexión entre el rasgo que cumple más efectivamente cierta función y el éxito reproductivo diferencial, y "adecuados" a los organismos que portan un rasgo que cumple una función más efectivamente, quedan explicitados los tres componentes fundamentales de la ley fundamental:
• Los organismos más adecuados tienden a mejorar su aptitud y, en consecuencia, su éxito reproductivo diferencial.4
La red teórica que incluye todas las diferentes especializaciones de TSN ya es compleja y unificadora en el texto fundacional de la teoría evolutiva (figura 1). Es un hecho realmente curioso que raramente las reconstrucciones ofrecidas de TSN no atrapen la heterogeneidad de sus diferentes aplicaciones. Esto debería exigirse a cualquier reconstrucción de TSN ofrecida.
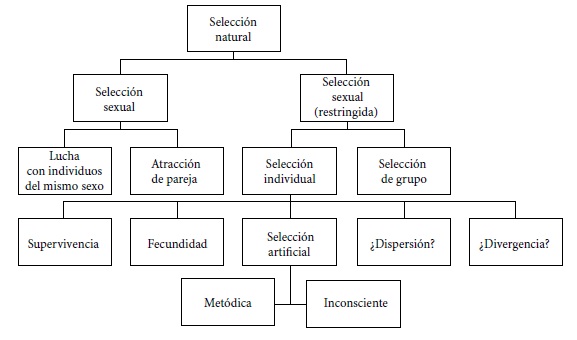
FIGURA 1. Red teórica de la teoría de la selección natural
4. Explicación como subsunción teórica ampliativa
La idea de que los conceptos teóricos en una teoría se introducen para explicar el comportamiento de las entidades descritas a través de términos no teóricos de la teoría está supuesta en algunos escritos estructuralistas. Así, por ejemplo, siguiendo líneas de trabajo planteadas por Bartelborth (1996, 2002), Diez (2002) y Forge (2002), Moulines intenta utilizar el estructuralismo metateórico para ofrecer una síntesis de los enfoques acerca de la explicación de los unificacionistas y causalistas. La estructura en red de las teorías, tal como son presentadas por los estructuralistas, permitiría entender mejor el unificacionismo, mientras que el componente causalista estaría dado, justamente, por la idea de que la explicación supone el enriquecimiento teórico, es decir, la introducción de nuevos conceptos, y que estos conceptos son T-teóricos en la teoría que los propone (y esto es lo que consideramos discutible):
Si aceptamos que tiene sentido hacer una distinción entre conceptos T-teóricos y conceptos T-no-teóricos dentro de una teoría T, y si admitimos una interpretación realista de los conceptos T-teóricos, entonces es plausible considerar las entidades a las que se refieren los conceptos T-teóricos como causas del comportamiento específico de SECI. Las entidades teóricas a las que se refieren los conceptos T-teóricos se introducirían, así, para proporcionar una explicación causal de los procesos fenoménicos codificados en nuestros modelos de datos. Ellas serían parte de la estructura causal "oculta" del mundo que es responsable de los fenómenos que observamos y codificamos por medios T-no-teóricos. (Moulines 2005)
Lorenzano, en su comentario al artículo de Moulines, pone reparos a la interpretación causal de los conceptos T-teóricos (Lorenzano 2005). Esto, según él, no sería siempre factible. De esta discusión no estoy interesado, al menos en esta oportunidad, en la cuestión de la causalidad, sino en la clara referencia a que los conceptos T-teóricos serían explicativos en las teorías en las que aparecen, y una teoría sería explicativa, por lo tanto, si propone nuevos conceptos teóricos. Esto elucidaría la intuición, por ejemplo, de que la teoría propuesta por Newton es explicativa en un sentido que no lo es la cinemática galileana, pues la primera propone nuevos conceptos teóricos (fuerza y masa) en la mecánica clásica newtoniana. Mi intención es mostrar, a partir del caso que nos ocupa, que, si bien podría afirmarse que toda explicación satisfactoria es ampliativa, en el senti-do de enriquecer conceptualmente la descripción previa con la que se conceptualiza el fenómeno que hay que explicar, no necesariamente esta ampliación debe llevarse a cabo por medio de conceptos teóricos en la teoría utilizada para explicar.
5. El estatus de teoricidad de aptitud
En una primera instancia, parece que el concepto explicativo introducido por la teoría de Darwin es el de aptitud. Porque a partir de tal concepto es posible dar una explicación tanto de cómo se ha fijado una adecuación en una población de organismos como de por qué un tipo de organismos dado está teniendo éxito reproductivo diferencial.
Podemos esperar entonces que el criterio de teoricidad informal estructuralista deje del lado de los no teóricos en esa teoría los conceptos de adecuación y éxito reproductivo, y del lado de los teóricos el concepto de aptitud. Veamos si esto es así.
Los tres conceptos, el grado de adecuación, el grado de aptitud y el grado de éxito en la reproducción diferencial son comparativos en una población dada en un ambiente dado. Son conceptos cuantitativos, pero que meramente establecen un orden de más y menos en la población de organismos en cuestión. Según ese criterio, serán teóricos en TSN si es necesaria TSN para aplicarlos, y no teóricos en TSN si no.
Veamos que ocurre en cada caso:
Rasgo adecuado
Que un organismo posea un rasgo que permite realizar una función con cierta efectividad puede ser establecido con independencia de TSN. Que las jirafas utilizan el cuello para alcanzar las ramas altas de los árboles puede ser determinado por el estudio del comportamiento de las jirafas, sin necesidad de la postulación de ningún mecanismo evolutivo y con independencia del origen histórico filogenético del rasgo. El grado de efectividad correspondiente a cada rasgo también puede ser determinado en ambientes particulares.
Éxito reproductivo diferencial
También puede ser determinado con independencia de TSN el éxito reproductivo de los organismos. Sin importar cómo se realice esta medición, evidentemente será de manera independiente de TSN, puesto que surgirá mediata o inmediatamente del conteo de descendencia fértil o viable de estos organismos. La genética de poblaciones, por ejemplo, permite actualmente y en ciertas condiciones ideales determinar este concepto con bastante precisión.
Aptitud
Si bien esperábamos que este fuese el concepto teórico en TSN, por su papel aparentemente explicativo, según el criterio informal, también es no teórico en TSN. Fuerza es concepto teórico en la mecánica clásica (MC), porque cuando se quiere aplicar el concepto se debe apelar a la ley fundamental de MC. Esto quiere decir que cuando se quiere medir una fuerza particular que actúa sobre cierta partícula, como la fuerza de gravedad, la fuerza de rozamiento, etc., se debe acudir, necesariamente, a la ley fundamental de MC. Cuando se quiere medir la aptitud de un individuo, tal como aparece en alguna ley especial, el caso es distinto, pues se puede medir la supervivencia, la capacidad para atraer parejas y/o la fecundidad, independiente de TSN.
Nos vemos en una disyuntiva: o abandonamos la idea de que este es un concepto explicativo en algún sentido en esta teoría, o abandonamos la pretensión de que el criterio informal estructuralista puede distinguir imbatiblemente entre conceptos explicativos y no explicativos. O, dicho de otro modo, que la distinción entre conceptos T-teóricos y T-no teóricos sea coextensiva con la distinción entre conceptos T-explicativos y T-no explicativos. O tal vez, que la distinción entre conceptos T-teóricos y T-no teóricos pueda elucidar de manera completamente satisfactoria la distinción intuitiva entre conceptos explicativos y no explicativos.
Creo, sin embargo, que hay que optar por alguna de las versiones del segundo de los disyuntos, pues hay varias razones para considerar que el concepto de aptitud tiene un estatus diferente al de los otros dos conceptos que el criterio de T-teoricidad informal no alcanza a atrapar:
1. Puede considerarse que es el concepto históricamente introducido con TSN. Darwin se dio cuenta de la relación que hay entre las funciones inmediatas de los rasgos con la aptitud, que podría ser caracterizada como la función evolutiva. Sin embargo, como ocurre con la aplicación del criterio informal estructuralista, las instancias del concepto de aptitud, como, por ejemplo, la supervivencia, obviamente no fueron propuestas con TSN. Por supuesto, como señala Hempel (1970), para detectar si un concepto se encuentra disponible con anterioridad a una teoría dada se presupone algún criterio semántico que permita distinguir conceptos novedosos de no novedosos. En qué sentido puede ser novedoso el concepto en cuestión, aunque sus instancias en leyes especiales no lo sean, se discutirá en el siguiente punto acerca de la dependencia semántica de aptitud con respecto a TSN.
2. Se puede afirmar que hay cierta dependencia semántica del concepto de aptitud con respecto a TSN, aunque sea TSN-no teórico (e independiente semánticamente en el sentido que permite determinar el criterio informal), pues:
a. Es posible medir la supervivencia de un tipo de organismos independiente de TSN. Pero parece claro que esto no implica que se está determinando su aptitud. Pues la mayor supervivencia de un tipo de organismos infértil, como una mula, no parece implicar una mayor aptitud. Lo mismo se puede decir de un caso en el que se desconoce el rasgo que cumple de manera más efectiva una función. Parece que la supervivencia de un organismo sólo es indicativa de su aptitud si tengo una explicación en donde se encuentran determinados los otros dos conceptos.
b. Lo único que tienen en común las diferentes instanciaciones del concepto de aptitud es, justamente, caer bajo ese concepto: fecundidad, emparejamiento, supervivencia caen bajo una misma clase únicamente por su papel en TSN. Dicho de otro modo, sólo cobran relevancia evolutiva por su papel en TSN.
3. Se da entre aptitud y los otros dos conceptos la asimetría típica que se da entre los conceptos explicativos y los explicados.
Del mismo modo en que estamos dispuestos a explicar cómo se acelera una partícula acudiendo a fuerzas que actúan sobre la partícula y su masa, pero no a aceptar la explicación de la masa de una partícula a partir de la aceleración de la partícula bajo ciertas fuerzas, estamos dispuestos a aceptar la explicación de que cierto tipo de organismos vivos tienen un mayor éxito reproductivo que otros por tener una mejor supervivencia, pero no a que sobrevivan más por tener un mejor éxito reproductivo.
4. Tal vez la razón más importante sea la siguiente: aptitud, como veíamos, es un concepto de naturaleza fuertemente abstracta, de cuya instanciación surge la red teórica de TSN. Su papel es parecido al de fuerza en MC. En ambos casos, la naturaleza abstracta del concepto tiene que ver con su fertilidad -incluso, es posible considerarlo un funcional, y por lo tanto considerar a la ley fundamental de TSN un principio guía, en el sentido utilizado por Moulines (1982)-.
Si bien estas razones me parecen indicativas de que aptitud tiene un estatus distinto al resto de los conceptos que aparecen en TSN, pese a ser TSN-no teórico, no creo, sin embargo, que esto constituya un contraejemplo al criterio de teoricidad informal estructuralista. No creo, por lo tanto, que haya que reformular ese criterio en ningún sentido. Su función principal e indiscutible es distinguir los conceptos que pueden ser determinados a partir de otras teorías científicas, y que, en consecuencia, sirven para contrastarla:
La distinción entre conceptos específicos para una teoría dada T, y los conceptos no específicos para T ha jugado un papel importante en el desarrollo de la filosofía de la ciencia en los últimos 40 años. La distinción está fuertemente conectada con la pregunta de cómo podemos proveer una "base empírica" para la teoría, es decir, un marco de conceptos por medio de los cuales la teoría pueda ser controlada, o por medio de los cuales podamos averiguar cuándo la teoría funciona correctamente y cuándo no. La "base empírica" de una teoría dada T debe incluir esos conceptos que, en un sentido que se precisará [con el criterio informal estructuralista], no son específicos de T. Los otros conceptos de T pertenecerán a la "superestructura" teorética de T. (Balzer, Moulines & Sneed 48)
Efectivamente, si se está intentando contrastar una explicación particular dada con TSN, por ejemplo: la cola del pavo real que, por su estructura, cumple mejor la función de atraer a las hembras mejora la capacidad de emparejarse de sus portadores, y mejora, en consecuencia, su éxito reproductivo diferencial, cada una de las partes del enunciado es determinable con independencia de las otras y de TSN, y, por lo tan-to, pueden ser determinadas para su contrastación. Podemos realizar experimentos y observaciones para determinar que efectivamente las colas de los pavos reales atraen a las hembras, o que son usadas por los pavos reales para eso, para determinar que los que portan ese rasgo tienen éxito en la reproducción diferencial, pero también podemos idear experimentos o realizar observaciones para determinar si se da la mejora en la aptitud señalada, es decir, si los pavos que portan ciertos rasgos logran conseguir más parejas reproductivas que los que portan otros (cf. Takahashi et al. 2008).
Parece, por consiguiente, que, en cuanto permite distinguir con claridad la base de contrastación, la aplicación del criterio informal estructuralista tiene éxito también en su aplicación a TSN. Sin embargo, creo que aptitud tiene un estatus distinto al de los otros conceptos de TSN que el criterio en cuestión no alcanza a resaltar. Creo, por las razones ya señaladas, que aptitud tiene un rol explicativo.
El criterio informal estructuralista brinda condiciones necesarias y suficientes con respecto a la cuestión de qué conceptos forman parte de la base de contrastación de una teoría, pero no lo hace (no al menos necesariamente) para determinar qué conceptos son explicativos en esa teoría. El concepto de aptitud sería explicativo, aunque no teórico en TSN.
6. Conclusiones
He intentado mostrar en este trabajo que, si bien es cierto que TSN es fenomenológica -en el sentido modificado del utilizado en textos clásicos de que ninguno de sus conceptos fundamentales es teórico, según el criterio estructuralista-, propone un nuevo concepto explicativo, el concepto de aptitud.
Tal concepto, dado que sus instanciaciones en leyes especiales pueden ser determinadas con independencia de TSN, debe ser considerado no teórico en TSN. Sin embargo, he dado razones por las que, de todos modos, su estatus es diferente al de los otros conceptos de TSN, ya que es introducido con TSN; depende semánticamente de TSN en el sentido antes señalado, estamos dispuestos a aceptar que la mayor aptitud de un organismo puede causar su mayor éxito reproductivo, pero no lo inverso, y, finalmente, su instanciación permite la obtención de las leyes especiales a través de las cuales TSN se aplica. Este concepto, por lo tanto, puede ser considerado no teórico pero explicativo en TSN y, por lo tanto, puede considerarse a TSN fenomenológica pero explicativa.
Este puede ser considerado un éxito más de la metateoría estructuralista, al pensar problemas metateóricos y elucidar teorías científicas, e incluso puede considerarse un éxito más de la distinción, acerca de la teoricidad estructuralista, de los conceptos que se determinan con independencia de las teorías y los que no, pero pone límites a la capacidad de tal distinción para diferenciar entre conceptos explicativos y no explicativos, y, en consecuencia, a su capacidad de distinguir entre teorías explicativas y no explicativas.
* Trabajo realizado con la ayuda del proyecto de investigación PICT Redes 2006 n.º 2007 de la Agencia Nacional de Promoción Científica y Tecnológica. Agradezco a SADAF (Sociedad Argentina de Análisis Filosófico), por otorgar a este trabajo el Premio Eduardo Rabossi - 2009 en la categoría de investigadores en formación. Así mismo los comentarios a versiones previas de este trabajo de Pablo Lorenzano, Mario Casanueva y Daniel Blanco, en especial a Christián Carman, con quien vengo reflexionando sobre los alcances de la distinción de la T-teoricidad del estructuralismo, cuestión central en este trabajo.
1 La elección de este término para nombrar a este concepto no carece de problemas. Aunque el término fitness de la genética de poblaciones suele traducirse al castellano con "aptitud", no deben confundirse ambos conceptos. La genética de poblaciones y la teoría de la selección natural son teorías distintas, aunque relacionadas. Tal confusión ha sido señalada por otros autores (Glymour 388, Moya 37-55, Rosenberg 110-111) y no pretendo tratar aquí la cuestión. Aunque la elección terminológica que propongo es problemática, otras elecciones no lo son menos. Espero entonces atención por parte del lector, y pido que intente aceptar mi terminología en pos de la comprensión de este trabajo.
2 Que en la ley fundamental de TSN aparezca el concepto de función, puede parecer relativamente controvertido. Mario Casanueva, en una comunicación personal, me ha sugerido reemplazar "la efectividad con la que se cumple una función", por la "efectividad con la que se soluciona un problema ambiental". Además de señalar que utilizar el término función me parece más natural (con respecto a la práctica habitual de los biólogos), no puedo aquí tratar el tema con mayor profundidad. Para un desarrollo más extenso sobre esta cuestión, véase Ginnobili (2009).
3 Una cuestión interesante que se puede resaltar es que, si se toma en cuenta que los conceptos que aparecen en TSN son más de los que normalmente se señalan en las reconstrucciones habituales, la polémica acerca de la unidad de selección parece hacerse más compleja. En esta explicación, el grupo aparece en el lugar de la aptitud y en el de éxito reproductivo. Aunque es posible imaginar instanciaciones de TSN en donde el grupo aparezca en otros lugares y combinaciones. El rasgo que cumple mejor la función podría ser un rasgo del grupo, y esto podría mejorar el éxito reproductivo de los individuos, por ejemplo. Pero no pretendo tratar esta cuestión aquí, pues excedería los objetivos de este trabajo.
4 Por un lado, no sostengo que estos sean todos los conceptos de la ley fundamental de TSN. Por ejemplo, puede ser necesario, en una reconstrucción más completa, incluir el concepto de herencia o ambiente. Esa presentación incompleta de la ley, sin embargo, es suficiente para seguir adelante con los objetivos de este trabajo. Por otro lado, no relacionaré mi propuesta reconstructiva con la que otros autores han hecho. Pero en este nivel informal de reconstrucción puede notarse que hay más conceptos fundamentales de los que normalmente son tomados en cuenta (cf. Brandon 1990, Kitcher 1993, Williams 1970, 1973). Para una comparación con otros enfoques, véase Ginnobili (2006).
Bibliografía
Balzer, W., Moulines, C. & Sneed, J. An Architectonic for Science: The Structuralist Program. Dordrecht & Lancaster: Reidel, 1987.
[ Links ]Bartelborth, T. "Scientific Explanation". Structuralist Theory of Science, Balzer, W. & Moulines, C. (eds.). Berlin: Walter de Gruyter, 1996.
[ Links ]Bartelborth, T. "Explanatory Unification", Synthese 130/1 (2002): 91-107.
[ Links ]Brandon, R. Adaptation and Enviroment. Princeton, NJ: Princeton University Press, 1990.
[ Links ]Darwin, C. On the Origin of Species by Means of Natural Selection. London: John Murray, 1859.
[ Links ]Darwin, C. The Origin of Species, 6th ed. London: John Murray, 1872.
[ Links ]Diez, J. A. "Explicación, unificación y subsunción". Pluralidad de la explicación científica, González, W. (ed.). Barcelona: Ariel, 2002. 73-93.
[ Links ]Endler, J. Natural Selection in the Wild. Princeton, NJ: Princeton University Press, 1986.
[ Links ]Endler, J. "Natural Selection: Current Usages". Keywords in Evolutionary Biology, Keller, F., Lloyd, E. & Lloyd, E. A. (eds.). Cambridge, MA: Harvard University Press, 1992. 220-224.
[ Links ]Forge, J. "Reflections on Structuralism and Scientific Explanation", Synthese 130/1 (2002): 109-121.
[ Links ]Ginnobili, S. "La teoría de la selección natural darwiniana". Tesis. Universidad de Buenos Aires, 2006.
[ Links ]Ginnobili, S. "Adaptación y función", Ludus Vitalis XVII/31 (2009): 3-24.
[ Links ]Ginnobili, S. "Selección artificial, selección sexual, selección natural" (por aparecer).
[ Links ]Glymour, B. "Wayward Modeling: Population Genetics and Natural Selection", Philosophy of Science 73/4 (2006): 369-389.
[ Links ]Hempel, C. G. "On the 'Standard Conception' of Scientific Theories". Minnesota Studies in the Philosophy of Science, vol. IV, Radner, M. & Winokur, S. (eds.). Minneapolis, MN: University of Minnesota Press, 1970. 142-163.
[ Links ]Kitcher, P. The Advancement of Science: Science without Legend, Objectivity without Illusions. New York / Oxford: Oxford University Press, 1993.
[ Links ]Klimovsky, G. Las desventuras del conocimiento científico. Buenos Aires: A-Z Editora, 1994.
[ Links ]Lorenzano, P. "Comentarios a 'Explicación teórica y compromisos ontológicos: un modelo estructuralista' de C. U. Moulines", Enrahonar: Quaderns de Filosofia 37 (2005): 55-59.
[ Links ]Moulines, C. U. Exploraciones metacientíficas. Madrid: Alianza Editorial, 1982.
[ Links ]Moulines, C. U. "Explicación teórica y compromisos ontológicos: un modelo estructuralista", Enrahonar: Quaderns de Filosofia 37 (2005): 37-53.
[ Links ]Moya, A. Sobre la estructura de la teoría de la evolución. Barcelona: Anthropos, 1989.
[ Links ]Putnam, H. "What Theories Are Not". Logic, Methodology and Philosophy of Science, Nagel, E., Suppes, P. & Tarski, A. (eds.). Stanford, CA: Stanford University Press, 1962.
[ Links ]Rosenberg, A. Instrumental Biology or the Disunity of Science. Chicago/London: The University of Chicago Press, 1994.
[ Links ]Takahashi, M., Arita, H., Hiraiwa-Hasegawa, M. & Hasegawa, T. "Peahens Do Not Prefer Peacocks with More Elaborate Trains", Animal Behaviour 75/4 (2008): 1209-1219.
[ Links ]Williams, M. B. "Deducing the Consequences of Evolution: A Mathematical Model", Journal of Theoretical Biology 29/3 (1970): 343-385.
[ Links ]Williams, M. B. "The Logical Status of the Theory of Natural Selection and Other Evolutionary Controversies". The Methodological Unity of Science, Bunge, M. (ed.). Dordrecht: Reidel, 1973. 343-385.
[ Links ]
