











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCTION
Chia (Salvia hispanica L.) is a pseudo-cereal known for its elevated nutritional value, with high content of α -linolenic and linolenic acids, fibers, proteins, minerals and anti-oxidant, and cardioprotective properties. Its consumption can be of whole seeds, oils, or flours (Ixtaina et al., 2011; Capitani et al., 2012). However, although the Rules for Seed Analysis (Ministério da Agricultura, Pecuária e Abastecimento, 2009) establish parameters for germination tests and quality control, few studies focused on chia seed technology (Possenti et al., 2016), e.g., the imbibition curve behavior and the physiological performance of seeds submitted to the physiological conditioning.
This technique is a pre-germination treatment that may directly affect seeds' performance, influencing the speed of their hydration; in turn, it may help in germination, as well as fast and uniform plantlet emergence (Santos et al., 2011). However, the rate and speed of water absorption respond in distinct ways, depending on the physiological conditions of the seed, among other factors (Popinigis, 1985). Among the methods of physiological conditioning, the hydro-conditioning (exclusive use of water to seed hydration); osmoconditioning (use of salts, and polyethylene glycol, mannitol solutions), and matriconditioning (involves the use of materials like clay, vermiculite, sand, and peat) can be mentioned (Marcos-Filho, 2015).
Matriconditioning is one of the most recent technologies in seed pretreatments (Queiroga et al., 2011). The objective of this method is slow hydration until the desired seed water level is achieved. For that to happen, the material must have a high capacity for retaining water, be insoluble in water and physically stable during the conditioning period, inert chemically, non-toxic to the seed, have a high surface area, and ease in the management and separation or removal of the seeds after the treatment (Marcos-Filho, 2015).
This study evaluated the effect of matriconditioning, under the imbibition curve and the physiological performance of two lots of chia seeds of different quality.
MATERIAL AND METHODS
The study was conducted in the Seed Analysis Laboratory of the Federal University of Technology - Paraná, Dois Vizinhos-Paraná state Campus, with two lots (L1 and L2) of chia seeds, produced by small-scale local farmers from the region of Vacaria, Rio Grande do Sul, Brazil, with initial germination of 42 and 83% respectively.
To the matriconditioning, initially, a water retention test was made on vermiculite, as suggested by the Rules for Seed Analysis - RAS (Ministério da Agricultura, Pecuária e Abastecimento, 2009), testing the retention capacity in 25, 50, 75 and 100% of moisture, to determine which one would be the most indicated to do the imbibition process. After the preliminary tests, the capacity of 100% of vermiculite water retention was selected because it presented an imbibition curve with a better stable pattern, similar to the three-phase pattern of seed imbibition proposed by Bewley & Black (1994).
This way, seeds were imbibed on a stainless steel screen covered with a germitest paper sheet inside plastic boxes from the gerbox type, containing 55.5g of sterilized vermiculite and moisturized with distilled water in 100% of the retention capacity. After the box was closed, they were sealed with masking tape and placed in a regulated germination chamber at 10ºC, in an attempt to reduce the embryo's physiological activity during the process. Seed imbibition was made from a systematic weighting of four replications of 100 seeds at 0, 2, 4, 8, 16 and 32 hours after the process had started.
To the monitoring of the imbibition and the expression of its results in graphic form, to obtain a curve, initially, the water level of the samples was determined by the hothouse method at 105 ± 3.0ºC, according to RAS rules (Ministério da Agricultura, Pecuária e Abastecimento, 2009). With the weighting of samples during the process and the final determination of moisture by the same described method, the moisture gains were determined in each imbibition period. The initial and final mass of each sample allowed for the calculation of the imbibition percentage, using the following formula: %E= (Pf−Pi) Pi. 100 where Pf, final mass of samples; Pi, initial mass of samples.
After each imbibition reading, the following physiological tests were made:
Germination. Made according to RAS (Ministério da Agricultura, Pecuária e Abastecimento, 2009), with four replications of 100 seeds, distributed in plastic boxes (gerbox), in two filter paper sheets moisturized with distilled water in the proportion of 2.5 times the papers’ mass. After seeding, the plastic boxes were kept at a temperature of 25°C for 14 days (Possenti et al., 2016). Simultaneously, the first germination count was evaluated seven days after the installation of the test. Results were expressed in the percentage of normal plants. b) Plantlet Length: Conducted according to the procedure described by Nakagawa (1999) with four replications of 25 seeds in paper roll substrate. After seedling, the paper roll stayed in a germination chamber for 14 days, at a temperature of 25°C, when the total length of normal plantlets in each replication was measured, as well as of the aerial part and root, with a millimeter paper, being the results expressed in cm plantlet-1. c) Emergence velocity index: Conducted in germination chamber, adapting the methodology proposed by Nakagawa (1999). Plastic trays were used, with dimensions of 23 x 23 x 35cm, containing sieved soil by wire threads sieve of 4 millimeters gratings. Four replications of 100 seeds were sowed at 2 centimeters of depth and covered with a fine soil layer. Emerged plantlet counts were made daily, always at the same time. The criterion of emergence was the complete emission of the cotyledons above the soil. The emergence velocity index was calculated with the formula of Maguire (1962): 𝐸𝑉𝐼 𝐸1 𝑁1 + 𝐸2 𝑁2 +…= 𝐸𝑛 𝑁𝑛 , where: EVI= emergence velocity index; E1, E2, En= number of plantlets emerged in the first counting, second counting, and last counting; N1, N2, Nn = number of days from seedling to the first, second and last counting. e) Accumulated emergence: Also evaluated with the emergence test, by the daily counting of emerged plantlets, calculating the cumulative percentage. Results were expressed by adjusted curves to a mathematical model that presented a higher representation. f) Emergence average time: Calculated using the data from the variable EVI, by the formula: = ( 𝑛𝑖𝑡𝑖 ) Σ𝑛𝑖 , where: ni= number of seeds emerged per day; ti = day in which the plantlet emerged. The results were expressed in number of days.
The experimental design was completely randomized, in a 2 x 6 factorial (two lots x six periods of imbibition), with four replications. After compilation, data were submitted to the Lilliefors test, to check for homoscedasticity. After the model assumptions were fit, an analysis of variance was performed to check the effect of the tested factors and their interactions by the F test. In case of a significant effect, Tukey's test with 5% probability was performed on the lot factor and regression analysis to the periods of imbibition.
RESULTS AND DISCUSSION
The ANOVA showed that the tested factors presented independent behavior for the measured variables. However, the lots showed a similar pattern in the water absorption by the seeds. In Figure 1, the imbibition curve of chia seeds submitted to the matriconditioning can be observed. In the first two and four hours of hydration, the biggest humidity gains occurred, coming from 7.5% to 14.57 and 20.13%, respectively in L1. In L2, initial moisture was 6.24% and went to 14.63 and 19.48 after two and four hours, respectively.
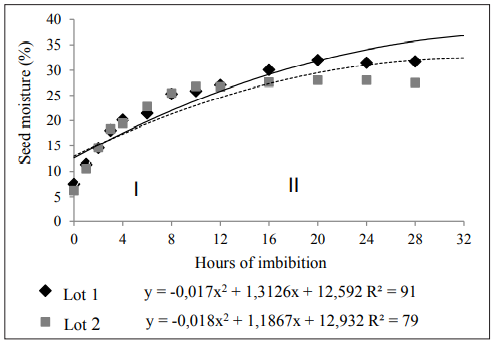
Phase I of imbibition has a duration of two to four hours (Carvalho & Nakagawa, 2012); however, in chia seeds, it lasted around eight to 16 hours, depending on the lot. According to Coll et al. (2001), some factors may interfere with the speed of absorption and with the quantity of imbibed water, like the nature and the composition of the tegument, increasing the duration of the phase. Furthermore, there is variation according to species, water availability, temperature, hydrostatic pressure, seed/water contact area, intermolecular forces, chemical composition, and seed physiologic conditions (Popinigis, 1985). Guimarães et al. (2011), studying imbibition in seeds of horsebush (Peltophorum dubium), observed that the speed of imbibition occurred slowly, at an average of 36 hours, in phase I. This fact was explained by the tegument dormancy in the seeds.
Phase II initiated from eight hours of imbibition to L2, whereas for L1, it initiated only after 16 hours. In this phase, the reduction in the water absorption speed and respiratory intensity takes place (Marcos-Filho, 2015), and the curve tends to stabilize. Still, according to this author, in phase II particular events occur like DNA repair, a new synthesis of enzymes, repair and multiplication of mitochondria, mobilization and assimilation of reserves, synthesis and duplication of DNA, and root cellular elongation.
Table 1, has the average values for the variables germination, first counting of germination, and total plantlet length. It can be observed that L1 showed lower germination and vigor, evaluated by the test of the first count of germination, than L2, which could explain the difference between them concerning the time to initiate phase II, having slower metabolic events, probably by its physiological conditions (Popinigis, 1985). Shoot and root length of plantlets did not show significant differences from the studied factors.
Table 1 Mean germination values (GER), first count of germination (FCG), and total plantlet length (TPL) of two lots (L1 and L2) of chia submitted to different imbibition periods.
| Imbibition hours | GER (%) | FCG (%) | TPL (cm) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| L1 | L2 | L1 | L2 | L1 | L2 | ||||||
| 0 | 42 b | 83 a | 19 | 78 | 12.37 | 12.19 | |||||
| 2 | 36 b | 88 a | 22 | 78 | 11.83 | 11.25 | |||||
| 4 | 50 b | 82 a | 24 | 76 | 10.63 | 10.45 | |||||
| 8 | 45 b | 83 a | 20 | 78 | 11.55 | 11.02 | |||||
| 16 | 47 b | 83 a | 22 | 80 | 11.64 | 11.43 | |||||
| 32 | 56 b | 76 a | 22 | 74 | 11.01 | 11.68 | |||||
| Average | 46 | 83 | 22 b | 77 a | 11.51 | 11.33 | |||||
| C.V. (%) | 5.1 | 6.8 | 6.1 | ||||||||
*Averages followed by the same lowercase letter in the row, for each variable, do not differ statistically by Tukey’s test with 5% of probability. C.V.: coefficient of variation.
For both lots, the behavior of germination in the function of the imbibition periods was adjusted to a quadratic model, yet, with a tendency to reduce its percentage for L2 and increase for L1, as the imbibition period increased (Figure 2).
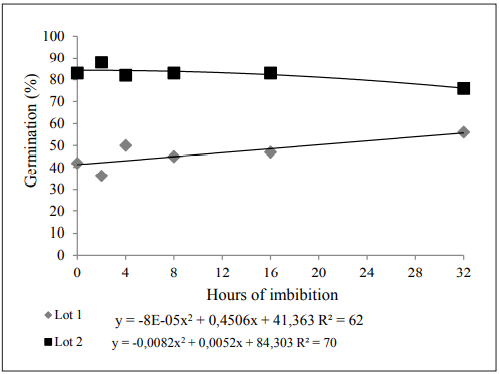
Pill & Kilian (2000) quoted that the matric conditioning might improve seed germination and it may be inferred that this improvement occurs, in general, in seeds that have lower initial germination, justifying the behavior in L1. Some authors report the influence of physiological conditioning on germination percentage from seeds and vigor (Kissmann et al., 2010; Masetto et al., 2013; Masetto et al., 2014). However, this result is linked to the standardization of the metabolic state and not to the vigor of seeds, occurring a “revitalization” because germination is practically instantaneous after the physiological conditioning (Marcos-Filho, 2015).
The periods of imbibition presented a significant difference in the variable total plantlet length, being adjusted to a quadratic model (Figure 3). The lowest length was registered with four hours of matriconditioning, with approximately 10cm. A decrease in length in the first hours of imbibition, followed by an increase and stabilization, was verified.
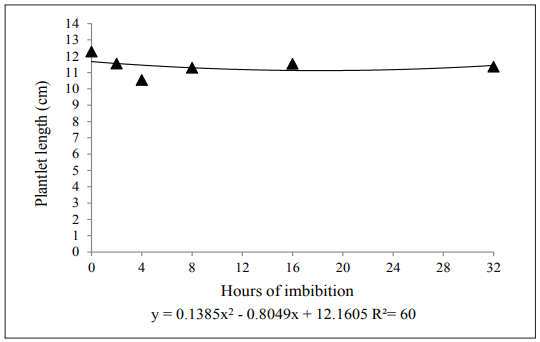
Guimarães et al. (2013) evaluating pre-imbibed watermelon seeds (0, 12, 24, 48, and 72 hours) observed that imbibition promotes a higher shoot, root, and total length. Similarly, Ferraz et al. (2017) found a relative increase in the average shoot length of cotton seeds hydro-conditioned in silicon solution for 24 hours in the concentration of 100mg L-1.
Positive results for plantlet length were also found for onion seeds in a study with hydro-conditioning. An increase in the total and shoot length of plantlets when compared to the seeds without hydro-conditioning was observed (Hölbig et al., 2011). Rabbani et al. (2013), working with horseradish seeds (Moringa oleifera Lam.), obtained the highest average for plantlet length (11.7cm) in seeds imbibed for 24 hours. These variations occur because the plantlet growth is dependent on the physiological potential of seeds, environment conditions, and the characteristics of the lots (Marcos-Filho, 2015).
The average for the variables Accumulated Emergence (EMER), Emergence Velocity Index (EVI), and Emergence Average Time (EAT) are presented in Table 2. It can be observed that the emergence velocity index and the average time of emergence were higher in lot 2 in all imbibition periods.
Table 2 Average values of accumulated emergence (EMER), emergence velocity index (EVI), and emergence average time (EAT) of two lots (L1 and L2) of chia submitted to different imbibition periods.
| Hours of imbibition | EMER (%) | EVI | EAT (days) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| L1 | L2 | L1 | L2 | L1 | L2 | ||||
| 0 | 21 | 67 | 3.67 b | 14.11 a | 6.1 b | 4.9 a | |||
| 2 | 23 | 77 | 5.21 b | 24.50 a | 4.7 b | 3.3 a | |||
| 4 | 37 | 79 | 9.72 b | 23.66 a | 4.1 b | 3.5 a | |||
| 8 | 39 | 90 | 9.29 b | 29.26 a | 4.4 b | 3.3 a | |||
| 16 | 46 | 87 | 13.28 b | 33.99 a | 4.0 b | 2.8 a | |||
| 32 | 51 | 99 | 13.87 b | 33.63 a | 4.1 b | 3.1 a | |||
| Average | 36 | 83 | 917 | 26.53 | 4.56 | 3.48 | |||
| C.V. (%) | 10 | 11.1 | 5.6 | ||||||
*Averages followed by the same lowercase letter in the row do not differ statistically by Tukey’s test with 5% of probability. C.V: coefficient of variation.
Curves for each imbibition period were adjusted for Accumulated Emergence. In both lots, the maximum emergence of seedlings occurred five days after sowing (Figure 4).
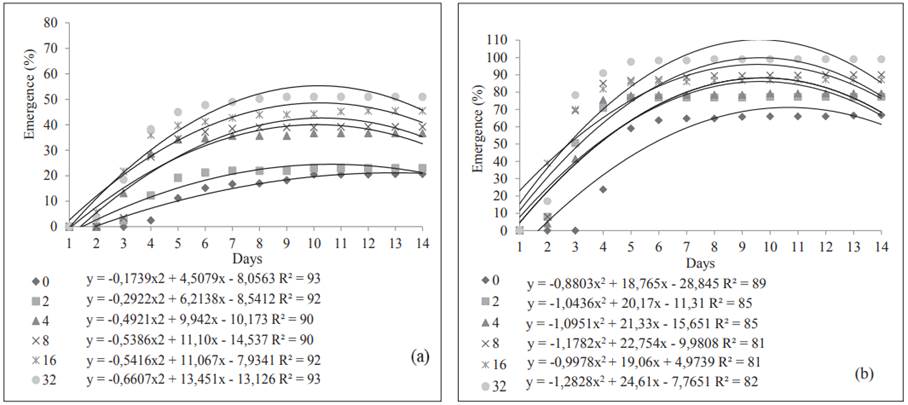
Figure 4 Accumulated emergence of plantlets originated from two lots, L1 (a) and L2 (b), of chia seeds submitted to six imbibition periods.
In the studied lots, the imbibition for 32 hours benefitted emergence, while non-conditioned seeds presented poorer performance, as well as those submitted for 2 hours of imbibition, especially in L1. The results from the present study differ from Armondes et al. (2016) who investigated cabbage seeds and verified no effect of osmotic conditioning in three of the four tested lots when they were submitted to osmoconditioning in PEG 6000 at -1,0 MPa for six days when compared to the control.
The emergence velocity index (Figure 5) was adjusted to a positive slope linear model in function of the imbibition periods, in both evaluated lots. For the average emergence time, lots were adjusted to a quadratic model (Figure 6).
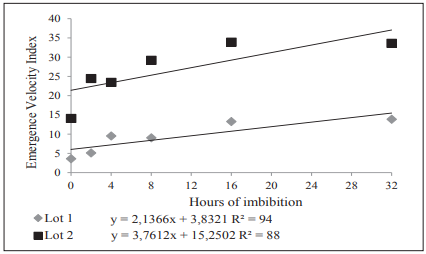
Figure 5 Emergence velocity index of chia plantlets originated from two seed lots submitted to six imbibition periods.
Figure 6 Emergence average time of chia plantlets originated from two seed lots submitted to six imbibition periods.
According to the results, matriconditioning positively influences plantlet emergence of chia, following the observed by Hölbig et al. (2011) who found superior results with the hydro-conditioning of onion seeds. When studying seeds of Crambe abyssinica, Hochst, Rocha et al. (2017) obtained a higher emergence velocity index in at least three of the four lots, when they were matriconditioned after three months of storage, but did not differ from the control.
Rabbani et al. (2013) found similar results when they observed a higher emergence velocity index in horseradish tree seeds (Moringa oleifera Lam.) when imbibed for 24 hours, and originated from three months of storage. The control of corn seeds also showed a lower emergence velocity index when compared to matriconditioned seeds (Andreoli & Andrade, 2003). In the case of cauliflower, the matriconditioning between four and six paper towels demonstrated benefits to the velocity of germination and emergence of plantlets, being that response dependent on the cultivar or the background of the lots (Kikuti & Marcos-Filho, 2009).

