










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Geología
Print version ISSN 0120-0283On-line version ISSN 2145-8553
bol.geol. vol.34 no.1 Bucaramanga Jan./June 2012
ESPECTROSCOPIA FTIR Y CRITERIOS PARA
LA IDENTIFICACIÓN DE ESTRUCTURAS ÓSEAS
FOSILIZADAS
Javier H. Jerez-Jaimes1; Silvia Elizabeth García-Gómez1; Luis Enrique Cruz-Guevara1.
1 Universidad Industrial de Santander, Escuela de Geología.
javjerez@gmail.com, silvis689@hotmail.com, lecruz@uis.edu.co
El registro fósil de vertebrados marinos es muy importante para la reconstrucción de filogenias sistemáticas, estudios ecológicos y evolutivos, por tal motivo, cualquier elemento tafonómico debe ser analizado con técnicas rápidas y sencillas antes de ser descartado. En este estudio se evaluó mediante análisis de espectroscopía de infrarrojos FTIR la procedencia biológica de una estructura similar a vértebras. Esta muestra fue colectada en la Formación Paja (Cretácico: Aptiano) en el municipio de La Mesa de los Santos (Santander, Colombia). Muestras de la capa externa y de la matriz de relleno del ejemplar fueron extraídas utilizando un microtaladro. Un hueso largo de vaca (Bos taurus) se usó como muestra control para la determinación del espectro del fosfato de calcio. Los espectros de FTIR permitieron identificar diferencias en la composición de la capa externa y la estructura interna del espécimen, encontrándose una respuesta en los rangos de absorción para el fosfato de calcio en la capa externa. La capa interna mostró una mayor composición en carbonato de calcio. La técnica de FTIR permitió la identificación de fosfato de calcio en esta entidad tafonómica. Los criterios paleontológicos actuales determinan que la morfología y los espectros de infrarrojos obtenidos no soportan el reconocimiento de este elemento como una estructura osificada fósil debido a la ausencia de la microestructura característica.
Palabras Claves: Aptiano, Formación Paja, Pseudofósil, Reptiles marinos.
¿FOSSIL BONE OR SIMPLE ROCK? SPECTROSCOPY FTIR AND CRITERIA
TO FOSSILIZED BONE STRUCTURES IDENTIFICATION
ABSTRACT
Fossil record of marine vertebrates is very important to the reconstruction of systematic phylogenies, evolutionary and ecological studies, for this reason, any taphonomic element must be analyzed with quick and simple technical before it is discarded. In this study, was evaluated the biological origin of a similar structure to the vertebrae with Infrared spectroscopy analysis FTIR. The structure was collected in the Paja Formation (Cretaceous: Aptian) of the municipality of La Mesa de los Santos (Santander, Colombia). Samples of the outer layer and the filling matrix were extracted using a microdrill. Along cow's bone (Bos taurus) was used as a control sample to determine the spectrum of calcium phosphate. FTIR spectra allowed identification of differences in the composition of the outer layer and the internal structure of the specimen. A response was found in the absorption ranges to calcium phosphate in the outer layer. The internal layer showed a greater composition of calcium carbonate. The FTIR technique allowed the recognition of calcium phosphate in this taphonomic entity. Current paleontological criteria determine that the morphology and the spectra IR not support the recognition of this element like fossil ossified structure due to the absence of the characteristic microstructure.
Keywords: Aptian, Marine Reptile, Pseudofossil, Paja Formation.
La identificación de huesos fósiles está determinada por el grado de conservación de la textura y microestructura ósea (Páramo, M. E., comunicación verbal), es decir, la observación de trabéculas, osteonas y microestructuras asociadas. Los huesos fósiles de los vertebrados contienen en su interior cavidades de distinto tamaño donde los minerales diagenéticos se depositan. La alteración en la composición original de los huesos fósiles ofrece un registro suplementario de la evolución de las condiciones físico-químicas de los fluidos intersticiales (Merino y Morales, 2006). Las concentraciones de ciertos elementos en huesos fósiles pueden ser muy diferentes de las originales. Estos elementos son incorporados dentro del hueso fósil principalmente por tres mecanismos: a) sustituciones isomorfas del apatito. Pueden ocurrir cambios por sustitución iónica dentro de la estructura cristalina del apatito sin llegar a afectarla; b) formación de minerales secundarios por precipitación en los poros y cavidades medulares; c) acumulación de minerales por procesos mecánicos en los poros y fracturas de los huesos por compactación de los sedimentos adyacentes (Merino y Morales, 2006).
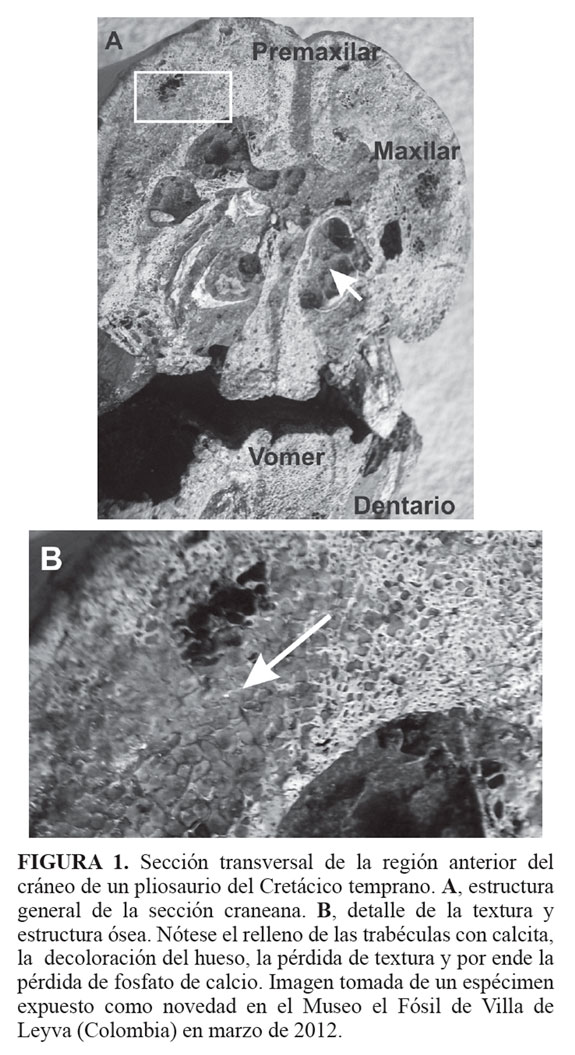
Algunos materiales fósiles conservan la morfología externa del hueso permitiendo su reconocimiento y consisten casi por completo de calcita (Holtz and Schultz, 1998). En secciones delgadas de este tipo de hueso se observa que el apatito original es reemplazado siendo casi ausente, sugiriendo no sólo la destrucción y dislocación si no también disolución y reemplazo de fosfato por carbonato de calcio (Holtz andSchultz, 1998). Huesos con algún signo macroscópico de expansión muestran un reemplazo total por calcita y hematita, así como ausencia de alguna estructura ósea original reconocible, aunque la morfología externa no está muy alterada es un indicativo que la fuerza de cristalización fue suficiente para romper y dislocar la estructura natural del hueso desde el interior (Holtz and Schultz, 1998). La FIGURA 1 permite comprender el proceso de relleno de la estructura trabecular por calcita y la pérdida de la textura característica del hueso.
La espectroscopía de infrarrojos es una técnica útil para el estudio de estructuras moleculares en diferentes productos industriales (Bhargava et al, 2003), minerales (García y Campos, 2000) o biológicos (Jerez-Jaimes y Bernal-Pérez, 2008, 2009). Un espectro de infrarrojo ocurre porque cada grupo funcional en una molécula tiene una frecuencia consistente en la cual absorbe radiación. La intensidad de la absorción es proporcional a la concentración, de este modo, si la concentración se incrementa la absorción se incrementa también y la transmisión disminuye (Hill and Rendell, 1975). La ventaja de la técnica tiene que ver con la mínima cantidad de material requerido y por lo tanto la conservación de los especímenes. En paleontología se ha utilizado la espectroscopia de infrarrojos (FTIR) en el estudio de coprolitos fósiles (Hallgren, 1987); De Figueredo-Souto (2006) estudió Crocodilimorfos (huesos, cáscaras de huevos y coprolitos) de la Formación Adamantina (Cretácico superior) en Sao Paulo (Brasil); Edwards et al. (en imprenta) utilizó la técnica para determinar la composición y tafonomía de tejidos blandos epidérmicos fosilizados de reptiles con una edad de 50 millones de años. De igual forma Alvarez-Lloret et al. (2006) cuantificaron los minerales en tejido óseo de reptiles actuales con este método.
En este estudio se evaluó la condición de un ejemplar colectado en 1998 que se asume corresponde a estructuras vertebrales de un reptil marino en la Formación Paja, parte superior (Cretácico: Aptiano temprano; Royero y Clavijo, 2001) del municipio de Los Santos (Santander, Colombia) utilizando espectroscopía de Infarrojos FTIR y planteando las siguientes preguntas: ¿Puede la resolución de la FTIR permitir la identificación de estructuras consideradas como huesos fósiles aunque no conserven la microestructura característica (osteonas, trabéculas)? ¿Existen diferencias en los espectros de FTIR de la estructura interna y externa del espécimen analizado?. Así mismo, con base en los criterios paleontológicos actuales, ¿se puede determinar si el elemento de estudio fue una estructura osificada?. Para poder resolver estos interrogantes se tomó como criterio para la determinación de estructuras óseas la presencia de fosfato de calcio en las muestras. En esta investigación se considera al ejemplar como una entidad tafonómica hipotética, conceptualizando la entidad tafonómica según Fernández-López (2000) como cualquier evidencia de una entidad biológica del pasado, siendo el ejemplar a analizar un elemento conservado.
Formación Paja (Kip)
Inicialmente descrita por Wheeler (en Morales et al., 1958); su localidad tipo es en la quebrada La Paja, afluente del río Sogamoso. Esta formación aflora al SW de La Mesa de Los Santos; hacia la base se compone de limolitas con concreciones de caliza y láminas de yeso muy fino, hacia el techo está conformada por limolitas micáceas muy físiles con concreciones de caliza intercaladas con capas de arenisca arcillosa y limolitas calcáreas (Díaz et al., 2009). De igual forma Royero y Clavijo (2001) describen esta unidad conformada por lutitas y shales gris oscuros a azulosos, fosilíferos, con intercalaciones de areniscas gris amarillentas, de grano fino, con algunas intercalaciones de shales grises, localmente arenosos, calcáreos, fosilíferos, localmente limosos a arenosos, con intercalaciones de areniscas gris amarillentas, de grano fino, también pequeñas intercalaciones de calizas grises, localmente arenosas, fosilíferas. Se estima que su depósito tuvo lugar en un ambiente epicontinental. El espesor varía entre 125 y 625 m. El límite estratigráfico de esta unidad con la suprayacente Formación Tablazo es concordante. La edad ha sido determinada del Barremiano inferior al Aptiano inferior. Se compara en parte con la Formación Tibú-Mercedes (Royero y Clavijo, 2001).
Reptiles Marinos del Cretácico
Los Sauropterygia fueron el grupo más grande de reptiles marinos del Mesozoico y comprendía dos linajes: Placodontos y Eosauropterigios (Rieppel, 2000). Los dos grupos tenían diseños corporales muy diferentes. Los placodontos fueron animales de cuello corto, cráneos pequeños y robustos. Los eosauropterigios tenían cuellos largos, cráneos angostos y muy pequeños en relación con la longitud del cuerpo. Los Placodontos estuvieron geográficamente restringidos al Mar de Tethys y cronológicamente al Triásico. Los Eosauropterigios estuvieron ampliamente dispersos tanto geográfica como cronológicamente. Los plesiosaurios son el clado más derivado de los eosauropterigios, son esencialmente diferentes de todos los demás miembros del grupo. Una de las diferencias radica en que el registro fósil de los plesiosaurios se inicia a principios del Jurásico mientras que los demás eosauropterigios aparecen desde el Triásico. Los miembros han sido modificados en aletas, la muñeca y el tobillo son bien osificados mientras que en los otros eosauropterigios son cartilaginosos (Motani, 2009). En Colombia se reconocen como reptiles marinos del Cretácico (Aptiano-Barremiano) al plesiosaurio Callawayasaurus colombiensis, el pliosaurio Kronosaurus boyacensis (Jerez-Jaimes y Narváez-Parra, 2001) y al ictiosaurio Platypterygius sachicarum (Páramo, 1997). De la parte superior de la Formación Rosablanca se ha descrito un caparazón de Testudine: Notoemys zapatocaensis por Cadena-Rueda and Gaffney (2005) quienes se refieren a esta formación como del Valanginiano tardío.
Sobre el espécimen
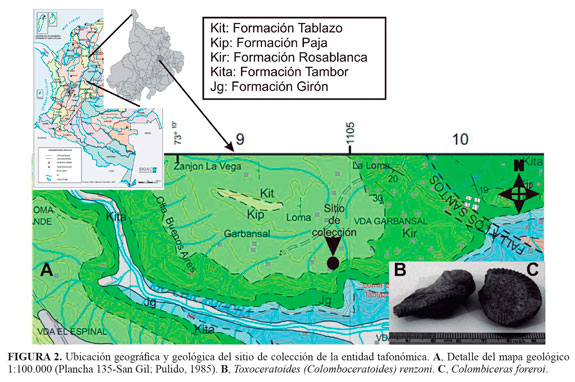
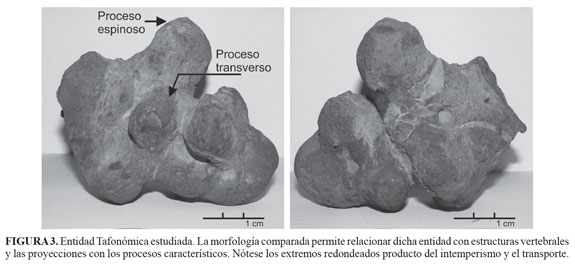
El elemento tafonómico fue colectado en 1998 en el municipio de Los Santos (Santander, Colombia) sobre una colina ubicada a dos horas de camino del municipio, en la vía que conduce hacia las minas de yeso y a unos 50 m de la carretera (FIGURA 2A), correspondiendo con el nivel superior de la Formación Paja. La presencia de las especies de amonites Toxoceratoides (Colomboceratoides) renzoni y Colombiceras foreroi (FIGURA 2B y FIGURA 2c) en el lugar del hallazgo, permite sugerir una datación Aptiano temprano para el ejemplar. Este presenta un alto grado de erosión y efectos del transporte mecánico e hídrico, observables en la reducción de la longitud y redondez de las proyecciones laterales y verticales que semejan los procesos vertebrales (apófisis espinosa y transversa). No se aprecian textura y estructuras macroscópicas (trabéculas), la cementación y reemplazo son aparentemente de calcita. Algunos sectores muestran los delineamientos de las estructuras vertebrales, así como una textura distinta a la del interior, probable relicto de estructura ósea (FIGURA 3). El espécimen se encontraba desprendido (rolledbone) del estrato, pero con un transporte pendiente abajo, se interpreta su posición original hacia la parte superior de la Formación Paja, ya que la secuencia sedimentaria se presenta normal y su contacto con la unidad suprayacente (Formación Tablazo) alcanza a aflorar hacia el NW del sitio de recolección (FIGURA 2), (Pulido, 1985; Díaz et al., 2009).
Debido a la ausencia de museos encargados de la conservación de ejemplares paleontológicos en la región, dicho material ha permanecido bajo la custodia del profesor Javier Jerez-Jaimes desde la fecha del hallazgo.
Espectroscopia de Infrarrojos FTIR
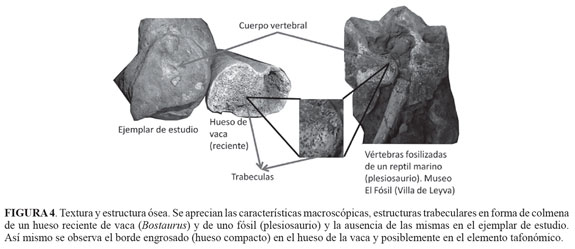
Para determinar la presencia de fosfato de calcio como marcador de estructuras óseas y de carbonato de calcio, se tomaron muestras de un hueso largo de un mamífero herbívoro (Bos taurus), colectado en el lecho de una quebrada, el ejemplar mostraba los efectos del intemperismo pues parte de la estructura esponjosa se había perdido (FIGURA 4). Se tomaron muestras de la parte exterior (hueso compacto) utilizando un microtaladro, para ser utilizado como espectro de fosfato de calcio de comparación. Del elemento tafonómico se tomaron muestras con el microtaladro, perforando hasta dos mm para minimizar los daños de la región interna que correspondería con el hueso esponjoso y de la capa externa -hueso compacto- (FIGURA 5), dicho material se mezcló con KBr y se procedió a analizarlo en un equipo Shimadzu FTIR 8400S de la Escuela de Ingeniería Química de la Universidad Industrial de Santander. Los espectros obtenidos se compararon entre sí. Se utilizaron como referencia para este estudio los picos para el fosfato de calcio y el carbonato de calcio puro, publicado por De Figueredo-Souto (2006).
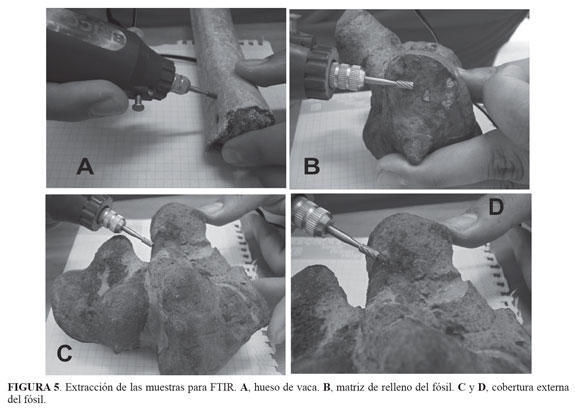
Observación macroscópica
El elemento tafonómico fue cubierto de yeso para evitar las fracturas al momento de cortarlo con la sierra (FIGURA 6A). El corte se realizó de manera longitudinal para obtener dos partes similares, una de estas se utilizó para elaborar una sección delgadas y secciones pulidas.
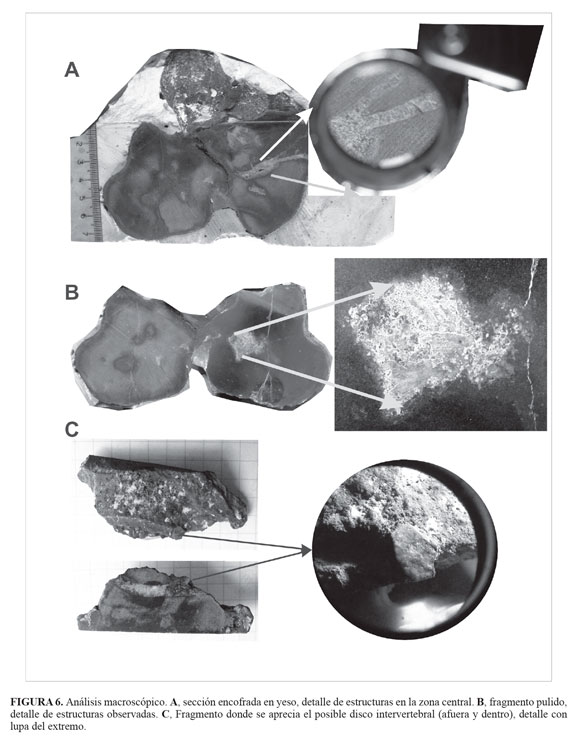
Observación microscópica
De la zona central de uno de los bulbos -centro vertebral hipotético, hueso de tipo canceloso- (FIGURA 6A) se extrajo un segmento longitudinal que presentaba una banda de color blanco y pequeños cristales, dicho segmento se utilizó para la elaboración de una sección delgada.
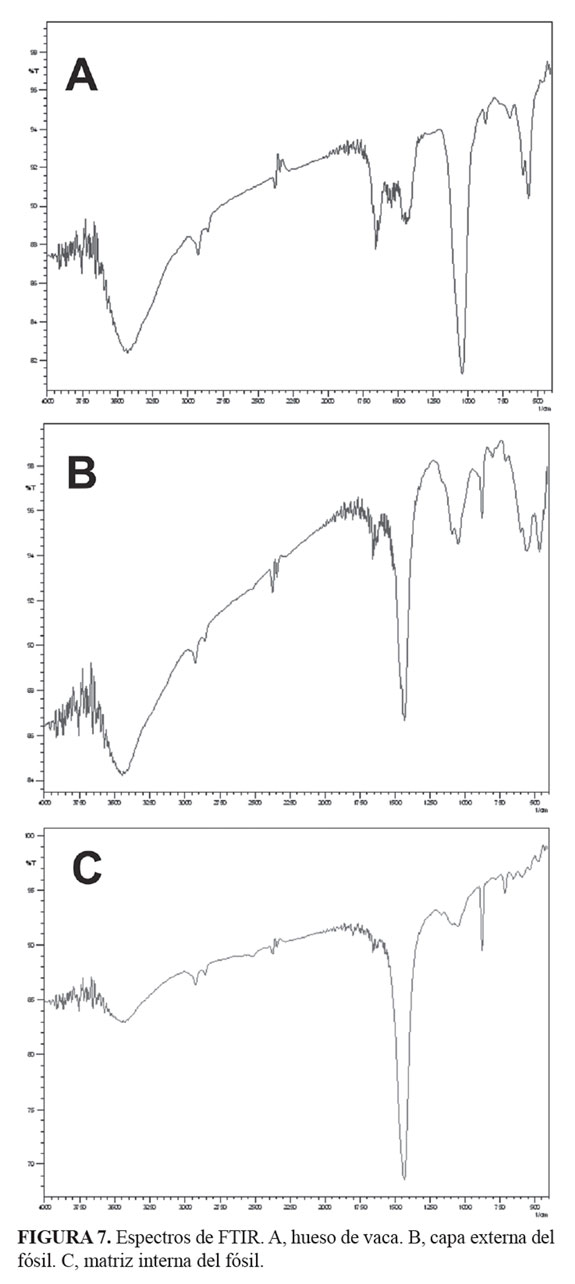
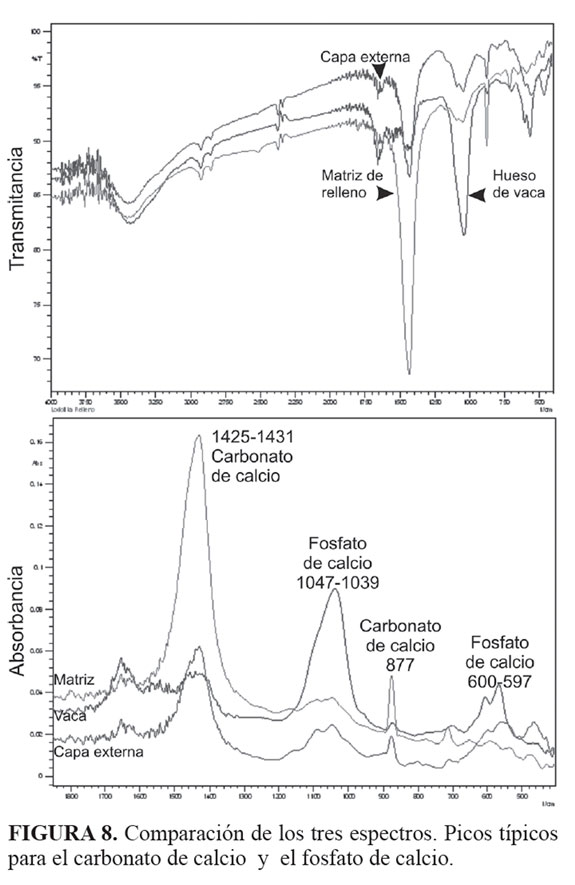
El espectro para el hueso de Bos taurus (control) mostró tres picos característicos del fosfato de calcio (1.039 nm, 600 nm, 566 nm), la capa externa del espécimen (fósil) presentó los picos característicos del apatito (1.091 nm, 1.047 nm, 597 nm, 561 nm) así como los picos del carbonato de calcio (1.425 nm, 877 nm), el material de relleno interno presentó picos con gran intensidad para la calcita (1.431 nm, 877 nm, 713 nm) y de baja intensidad para el fosfato de calcio (1.092 nm, 1.047 nm, 592 nm) FIGURAS (7 y 8). El pico en los 600 nm característico del fosfato de calcio no está resuelto en la matriz rica en carbonato de calcio (calcita). La TABLA 1 presenta los valores característicos para el carbonato de calcio (CaCO3), el fosfato de calcio (Ca2P2O3) y los obtenidos para el hueso de vaca (fosfato de calcio-apatito), la matriz de relleno y la capa externa del ejemplar.
Estructuras macroscópicas
Al interior del elemento tafonómico se observaron acumulaciones de color blanco hacia la zona central (FIGURA 6A y FIGURA 6B), sobre dichas acumulaciones se realizó la prueba de Deniges para determinar la presencia de fosfatos, obteniéndose un color amarillo pálido, indicando una respuesta positiva y una baja concentración, confirmando con esto los espectros de infrarrojos obtenidos. No se distinguió claramente el arreglo en forma de colmena (trabéculas) característico del tejido óseo esponjoso en estas áreas donde se conserva fosfato. En una de las secciones bulbosas (centro vertebral hipotético) se observó en la región central un área de color blanco con una retícula diferenciable (FIGURA 6B). En la FIGURA 6C se puede observar un fragmento del elemento tafonómico donde se aprecia una banda (disco intervertebral hipotético) en el exterior y su continuidad en el interior de color blanco, al observar con una lupa de 10 aumentos el extremo de esa banda, se aprecia un arreglo de cavidades circulares muy pequeñas (condrocitos hipotéticos), ya que esta estructura estaría constituida por cartílago y tejido conectivo.
Estructura microscópica
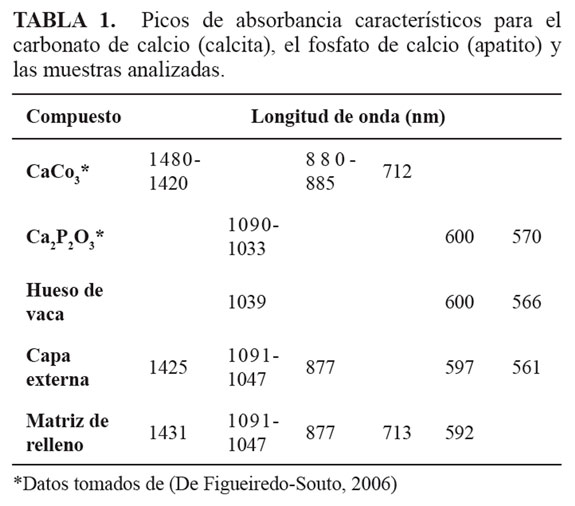
La sección delgada permitió la observación de cristales de calcita que componen la mayor parte del elemento de estudio (FIGURA 9A). Se observaron asociaciones de estructuras peloides rodeados por una matriz de esparita (FIGURA 9B). Dichos peloides poligonales mostraron un arreglo en forma de roseta (FIGURA 9C, FIGURA 9D y FIGURA 9E). No se observaron estructuras que indiquen la conservación de trabéculas o espacios de resorción óseos.
¿Por qué comparar un posible hueso de reptil marino con un hueso de mamífero herbívoro?
Es lógico pensar que los mamíferos y los reptiles no tienen mucho en común, aunque es una idea infundada si lo que se quiere analizar es la composición química de los huesos, siendo ésta similar en ambos grupos, además se debe recordar que los mamíferos descienden de una rama basal de los reptiles Pelicosauria-Therapsidos mamiferoides según Kemp (2006). De igual forma las diferencias que se han encontrado con la estructura ósea de los primeros mamíferos y los Cynodontes y los Therapsidos no mamíferoides tiene que ver con la velocidad de crecimiento y formación del hueso durante el desarrollo ontogénico (Chinsamy and Hurum, 2006), no con su composición química-mineralógica. La composición química y mineralógica de los huesos en los diferentes grupos de vertebrados es básicamente la misma: fosfato de calcio, carbonato de calcio y apatito como mineral. Morgulis (1931) realizó un estudio de las cenizas de los huesos de diferentes grupos de vertebrados actuales incluyendo peces, anfibios, reptiles, aves y mamíferos, mostrando que en todos los grupos los huesos estaban constituidos por fosfato de calcio y carbonato de calcio, así mismo, encontró que había una relación fosfato de calcio/carbonato de calcio mayor en los peces marinos que en los vertebrados terrestres, es decir, había más fosfato de calcio en los huesos de los vertebrados marinos que en los terrestres.
A partir del trabajo de Morgulis (1931) se calculó una media de 6,61 en la relación fosfato de calcio/carbonato de calcio para los vertebrados terrestres y de 11,77 para los peces marinos. Esto permite establecer que los vertebrados terrestres fijan una proporción significativamente mayor de carbonato de calcio en los huesos que los peces marinos. Desde el punto de vista de la textura y estructura las diferencias macroscópicas son mínimas como ya se evidenció en las FIGURAS 1 y 3.
El análisis de los espectros de FTIR confirmó que el ejemplar corresponde a una estructura que conserva una proporción de fosfato de calcio (FIGURAS 7 y 8). La técnica de infrarrojos es muy precisa en esta determinación como lo demostraron Hallgren (1987) y De Figueiredo-Souto (2006) en sus análisis de estructuras biogénicas, coprolitos y cáscaras de huevo. Es claro que se encontraron diferencias en los espectros de las dos capas del elemento tafonómico estudiado.
¿Hueso fósil o simple roca?
Mike Everhart (comunicación verbal) curador adjunto de Paleontología del Museo Sternberg de Historia Natural de la ciudad de Hays en Kansas, revisó imágenes del elemento tafonómico y confirmó la similitud con vértebras, pero consideró, basado en su experiencia de campo, el hecho muy probable de que sea una concreción. María Páramo (comunicación verbal) profesora de paleontología de Vertebrados de la Universidad Nacional de Colombia, se centró en la ausencia de textura y estructura ósea descartando de hecho que el elemento fuese un fósil.
Las evidencias obtenidas en el estudio macroscópico confirman una concentración mínima de fosfato de calcio en una estructura de orificios pequeños no definida claramente (FIGURA 6B). El fragmento que conserva un segmento del posible disco intervertebral mostró una retícula de círculos muy pequeños en un extremo de la banda, similar al arreglo del cartílago y el tejido conectivo (FIGURA 6C). El análisis microscópico no aportó información sobre la conservación de una estructura ósea, los peloides observados presentan bordes angulares, característica que los desvía de la descripción general como formas esféricas o elípticas, aunque se hace interesante su distribución en roseta en algunos sectores de la sección que no pudimos asociar con alguna organización tisular. Clarke and Barker (1995) habían demostrado la posibilidad de identificar el deterioro óseo patológico y mecánico en vertebras fosilizadas de Iguanodon spp mediante el estudio de secciones delgadas de cortes longitudinales y transversales, nada de lo encontrado en la sección delgada estudiada en este trabajo es similar a lo reportado por estos autores.
¿Qué indican los espectros de infrarrojos?
La presencia del fosfato de calcio en el elemento tafonómico no fue considerado por los dos expertos citados anteriormente. Se debe comprender que la altura de los picos está en estrecha relación con la concentración de los compuestos presentes y que los picos aparecerán corridos de su posición con base en el control, debido a que se estudian mezclas de compuestos y no compuestos puros. En las FIGURAS 7 y 8 se observa que los picos de baja intensidad para el fosfato de calcio en 600 nm y 570 nm (compuesto puro) se correlacionan en el hueso de vaca en 600 nm y 566 nm reconociendo la mezcla con carbonato de calcio típica de todos los huesos, y en 597 nm y 561 nm para la capa externa del elemento tafonómico (TABLA 1). La capa interna no produjo una respuesta en esa longitud de onda ya que prácticamente está constituida por calcita.
Las diferencias en los espectros de la matriz de relleno y la capa externa del elemento tafonómico se deben principalmente a cambios diagenéticos que afectaron la estructura y composición del elemento tafonómico.
Las inferencias son válidas sólo si el elemento es un hueso fosilizado, ¿ficción o realidad?
El hueso cortical (capa externa) es mucho más resistente y menos esponjoso que la región interna trabecular, por lo que el relleno y posterior reemplazo es mucho más rápido en esta última donde el fosfato de calcio se encuentra en baja proporción, debido al tamaño de las cavidades o trabéculas. Gran parte de la capa externa del hueso cortical se perdió por el transporte y la erosión. También se puede establecer que el elemento sufrió un reemplazo del fosfato de calcio por carbonato de calcio pudiéndose asumir que la calcita fue transportada en solución a través del tejido óseo, precipitada en los espacios vacíos del interior y durante la cristalización reaccionó con la red del apatito original perturbándola, causando dislocación de la estructura ósea original. Esta explicación se basa en el modelo de Holtz and Schultz (1998) creado para explicar las diferencias morfológicas y volumétricas en cráneos y extremidades de reptiles del Triásico medio de Brasil, donde la cementación inicial con calcita fue la responsable de la pérdida de textura y estructura, además de la deformación de los huesos. Para que el modelo funcione se debe considerar un enterramiento somero y estaciones bien marcadas de humedad y sequía. Se debe aclarar que el proceso de disolución del apatito y reemplazo por calcita no está entendido completamente.
¿Algo mordió al elemento tafonómico?
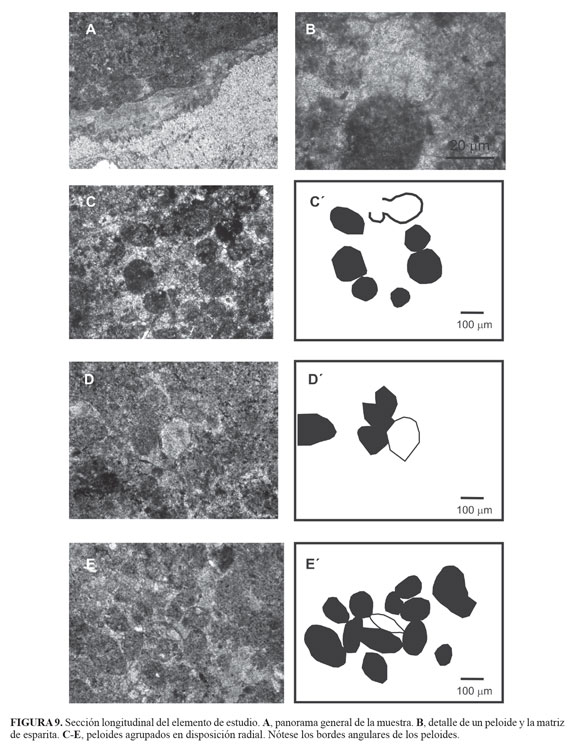
Al revisar con detenimiento la textura y estructura relictual del elemento analizado se puede observar que la capa exterior de una de las proyecciones verticales (proceso espinoso) está fracturada en pequeñas piezas alrededor de un segmento faltante de forma triangular (FIGURA 10), ¿Es ésto acaso la evidencia de una mordida? o ¿es el producto de la diagénesis secundaria de la roca?.
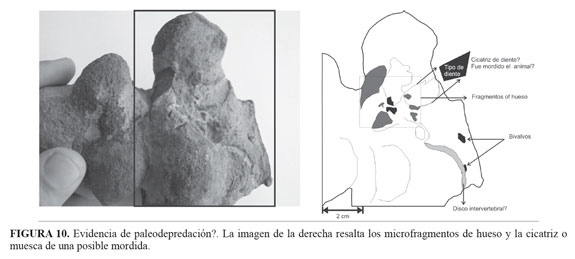
Método de razonamiento lógico para identificar un hueso fosilizado
Los espectros de infrarrojos respondieron a las preguntas planteadas, detectaron el fosfato de calcio y determinaron las diferencias en composición de las capas del elemento tafonómico analizado. Pero no tenemos la suficiente evidencia para determinar dicho elemento como una estructura ósea fosilizada. Tres criterios determinan o valoran la identidad de una estructura ósea fosilizada: morfología, composición y la estructura (TABLA 2). Cada criterio aporta una cantidad de información valorada por los investigadores. El criterio con mayor puntuación es la estructura ósea, básicamente porque es el más evidente y fácil de confirmar, con el inconveniente de que se debe sacrificar una buena parte del elemento tafonómico al momento de elaborar secciones delgadas. El segundo criterio es la morfología, si el elemento es similar a uno ya conocido es muy probable que ese sea, aunque si carece de la microestructura ósea muy seguramente será desechado. Por esta razón se debe reconocer que un hueso en su proceso diagenético puede perder toda su microestructura característica y conservar la forma e inclusive pudiese ser reemplazado en otro mineral muy diferente al apatito.
Finalmente, se debe considerar la composición química, todos los huesos fósiles desde el punto de vista mineralógico y en términos generales están conformados por carbonato de calcio (calcita) y fosfato de calcio (apatito), pero también es claro que la calcita puede reemplazar parcial o completamente al apatito. De igual forma el fosfato de calcio puede precipitarse en apatito por autigénesis y actividad microbiana. Los fosfatos pueden formarse en ambientes no upwelling como lagos someros, estuarios o deltas, donde el fosfato es aportado por los sistemas fluviales que contienen detritos orgánicos ricos en fósforo y azufre (Ruttenberg and Bemer, 1993), proceso que debe considerarse en los ambientes someros del cretácico inferior de Colombia. Los niveles de fosfato disuelto se incrementan en los sedimentos en condiciones de anoxia y de actividad microbiana. Una vez alcanzada la concentración de fosfatos requerida en los sedimentos, la formación de apatito autigénico comienza (Lamboy, 1993).
Si otorgamos un valor de 60 puntos sobre 100 al criterio "estructura" basados en la tendencia paleontológica actual y 20 puntos para cada una: composición y morfología, tendríamos que, en una matriz simple de valoración y selección de tres por tres, sólo serían seleccionados el 55,5 % de los elementos evaluados y aceptados como estructuras óseas fosilizadas (TABLA 3). En este caso en particular, el elemento analizado sólo consiguió 20 puntos por su morfología (similar a vértebras), 20 puntos por la composición (presencia de fosfato de calcio) y sumaremos 20 puntos (de 60) por la información estructural, totalizaremos 60 puntos. ¿Qué decisión debemos tomar con base en este modelo arbitrario? ¿Aceptamos el elemento tafonómico como un fósil, a pesar que el puntaje mínimo requerido sea superior a 60?, o ¿lo rechazamos porque el puntaje alcanzado genera incertidumbre?.
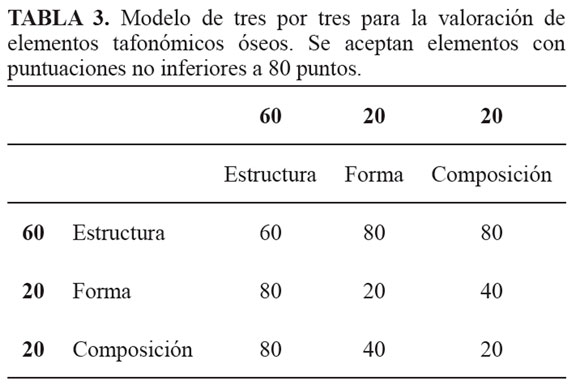
Con base en este análisis se puede inferir que el elemento analizado posee fosfato de calcio y carbonato de calcio en su estructura externa y sólo carbonato de calcio en su estructura interna. De igual forma se demostró que la técnica de FTIR puede ser una alternativa rápida y sencilla para aquellas entidades tafonómicas que no conservan la microestructura característica del tejido óseo y se quiera confirmar la composición química y mineralógica. De igual forma, consideramos que rechazar sin evaluar un elemento tafonómico que carece de estructura y textura, pero que presenta una morfología comparable, implica un desconocimiento y negación de los procesos diagenéticos primarios y secundarios.
Concluimos entonces, que nos encontramos en un limbo conceptual con este tipo de "elementos tafonómicos dudosos" que cuestionan nuestra experticia en los tres criterios de razonamiento lógico para identificar estructuras óseas fosilizadas. La decisión de optar por una prueba destructiva cuando se tiene un sólo elemento tafonómico para confirmar el criterio de la estructura microscópica implica una decisión extrema y no acorde con la conservación de un patrimonio cultural irrepetible -aunque en este trabajo se asumió el riesgo-, pues si el proceso confirma a la entidad, ésta dejará de existir o será seccionada.
Para finalizar, consideramos importante llevar a cabo una síntesis profunda de la información disponible sobre los procesos diagenéticos y los efectos de estos en la estructura, textura, forma y composición de entidades tafonómicas óseas.
Los autores agradecen a Guillermo Acero Medina del laboratorio de análisis térmico de la Escuela de Ingeniería Química de la Universidad Industrial de Santander (UIS) por la elaboración de las pruebas de FTIR. De igual forma agradecemos a los estudiantes Julián David De Bedout (Geología) por suministrar el material óseo reciente (Bos taurus) y a Claudio Antonio Mendoza Jaimes (Ingeniería Química) por su colaboración en la extracción de las muestras para los análisis de FTIR. A Luis Garrido de la Escuela de Geología de la (UIS) por la elaboración de la sección delgada. Al biólogo Richard Méndez Torres (AMGEN PR) por la revisión del Abstract y sus comentarios. Se agradecen también los comentarios y sugerencias de los evaluadores anónimos. Así mismo, se reconocen los aportes del profesor Francisco Velandia (UIS) en la posición estratigráfica y a la profesora Eliana X. Narváez Parra (UDES) en la datación con fósiles de amonites del espécimen analizado.
Alvarez-Lloret, P., Rodriguez-Navarro, A.B., Romanek, Ch.S., Gaines, K.S., and Congdon, J. 2006. Quantitative Analysis of Bone Mineral Using FTIR. Macla, 6:45-47. [ Links ]
Bhargava, R.,Wang, S., and Koenig, L. 2003.FTIR Microspectroscopy of Polymeric Systems.Advances in Polymer Science, 163: 93-96. [ Links ]
Cadena-Rueda, E.A., and Gaffney, E. S. 2005. Notoemyszapatocaensis, a New Side-Necked Turtle (Pleurodira: Platychelyidae) from the EarlyCretaceous of Colombia. American Museum Novitates.American Museum of Natural History.No. 3470. [ Links ]
Chinsamy, A., and Hurum, J.H. 2006. Bone microstructure and growth patterns of early mammals. ActaPalaeontologicaPolonica,51 (2): 325–338. [ Links ]
Clarke, J., and Barker, M. 1995. Abnormal histology in an Iguanodon caudal centrum from the lower Cretaceous of the Isle of Wight. Paleontology, 38(4): 905-914. [ Links ]
De Figueiredo-Souto, P.R. 2006. Use of Infrared Spectroscopy Analysis to Fossil Structures of Baru Basin, Brazil.Geociencias, 25 (1): 99-104. [ Links ]
Díaz, J. E., Contreras, N. M., Pinto, J. E., Velandia, F., Morales, C. J., e Hincapié, G. 2009. Evaluación Hidrogeológica Preliminar de las Unidades Geológicas de La Mesa de los Santos, Santander. Boletín de Geología, UIS, 31(1): 61-70. [ Links ]
Edwards, N.P., Barden H.E., van Dongen, B.E., Manning, P.L., Larson P.L., Bergmann, U., Sellers, W.I., and Wogelius, R.A. 2011. Infrared mapping resolves soft tissue preservation in 50 million year-old reptile skin. Proceedings of the Royal Society B., 278(1722): 3209-3218. Manuscrito en imprenta.doi:10.1098/rspb.2011.0135. [ Links ]
Fernández López, S.R. 2000. Temas de Tafonomía. Departamento de Paleontología. Facultad de CienciasGeológicas. Universidad Complutense de Madrid, 167p. [ Links ]
García, R.C.A., y Campos, A.N.O. 2000. Composición Química y Mineralógica de las Biotitas Metamórficas del Sector Central del Macizo de Santander, Colombia. Boletín de Geología, UIS, 22(37): 18-27. [ Links ]
Hallgren, L. 1987. Infrared spectroscopy analysis of fossil coprolites. Scientific Publications of the Science Museum of Minnesota, 6(2): 1-31. [ Links ]
Hill, R.R., and Rendel, D.A. E. 1975.The Interpretation of Infrared Spectra.London: Heyden, 375 pp. [ Links ]
Holtz, M., and Schultz, C.L. 1998.Taphonomy of the South Brazilian Triassic Herpetofauna: fossilization mode and implications for morphological studies. Lethaia, 31: 335-345. [ Links ]
Jerez-Jaimes, J.H., y Bernal-Perez, M.C. 2009. Taxonomía de onicóforos de Santander, Colombia y termogravimetría,calorimetría de barrido diferencial y espectroscopía infrarroja de lasecreción adhesiva (Onychophora: Peripatidae). Rev. Biol. Trop. 57 (3): 567-588. [ Links ]
Jerez-Jaimes, J.H., y Bernal-Pérez, M.C. 2008. Análisis de la seda del refugio de la araña Plexippuspaikulli (Audouin 1826) (Salticidae) mediante termogravimetría (TGA) espectroscopia de Infrarrojos (FT-IR). Memorias del XXXV Congreso SOCOLEN Sociedad Colombiana de Entomología, 59p. [ Links ]
Jerez-Jaimes, J.H., y Narváez-Parra, E.X. 2001. Callawayasauruscolombiensis (Welles) Carpenter 1999. El Plesiosaurio de Villa de Leyva (Boyacá, Colombia) ¿Un Nuevo Espécimen? Boletín de Geología, UIS, 23(38): 9-19. [ Links ]
Julivert, M. 1968. Léxico Estratigráfico Internacional. Unión Internationale des SciencesGeologiques. Paris. 415p. [ Links ]
Kemp, T.S. 2006. The origin and early radiation of the therapsid mammal-like reptiles: a palaeobiological hypothesis. European Society for Evolutionary Biology, 19:1231-1247. [ Links ]
Lamboy, M. 1993. Phosphatization of calcium carbonate in phosphorites: microstructure and importance. Sedimentology, 40(1):53-62. [ Links ]
Merino, J., y Morales, J. 2006. Mineralogía y Geoquímica del esqueleto de los mastodontes de los yacimientos Batallones 1, 2 y 5. Implicaciones tafonómicas. Estudios Geológicos, 62 (1): 53-64. [ Links ]
Motani, R. 2009. The Evolution of Marine Reptiles.EvoEdu Outreach. 2:224-235. [ Links ]
Morales, J.L. 1958.General Geology and Oil Ocurrences of Middle Magdalena Valley, Colombia.AAPG habitat of oil symposium, p. 641-695. [ Links ]
Morgulis, S. 1931. Studies on the Chemical Composition of Bone Ash.TheJournal of BiologicalChemistry, 93:455-466. [ Links ]
Páramo, M. E. 1997. Platypterygiussachicarum (Reptilia, Ichthyosauria) nueva especie del Cretácico de Colombia». Revista INGEOMINAS,6: 1–12. [ Links ]
Pulido, O. 1985. Geología de la Plancha 135-San Gil. Escala 1:100.000. INGEOMINAS. [ Links ]
Rieppel, O. 2000. Sauropterygia I. Handbuch der Paläoherpetologie 12A. München: Verlag Dr. Friedrich Pfeil, 134p. [ Links ]
Royero, J.M., y Clavijo, J. 2001. Mapa Geológico Generalizado Departamento de Santander Escala 1.400.000. Memoria Explicativa. INGEOMINAS. 92p. [ Links ]
Ruttenberg, K. C., and Bemer, R. A. 1993. Authigenic Apatite Formation and Burialin sediments from non-upwelling, continental margin environments.Geochim. Cosmochim. Acta. 57:991-1007. [ Links ]
Trabajo recibido: Julio 26 de 2011
Trabajo aceptado: Junio 15 de 2012

