










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Geología
Print version ISSN 0120-0283
bol.geol. vol.38 no.4 Bucaramanga Oct./Dec. 2016
https://doi.org/10.18273/revbol.v38n4-2016002
DOI: http://dx.doi.org/10.18273/revbol.v38n4-2016002
DEL CRETÁCICO INFERIOR EN LA CUENCA DEL
VALLE SUPERIOR DEL MAGDALENA, COLOMBIA
Camila Monje Dussán1,2,5; Camila Martínez3; Ignacio Escapa4; Santiago Madriñán1
1 Laboratorio de Botánica, Universidad de los Andes, Bogotá, Colombia. camila.monje.dussan@gmail.com
2 Smithsonian Tropical Research Institute, Panamá, República de Panamá.
3 L. H. Bailey Hortorium, Department of Plant Biology, Cornell University, Ithaca, New York 14850 USA.
4 CONICET, Museo Paleontológico Egidio Feruglio, Trelew, Patagonia, Argentina.
5 Corporación Geológica ARES, Bogotá, Colombia.
Forma de citar: Monje-Dussan, C., Martínez, C., Escapa, I., y Madriñán, S. 2016. Nuevos registros de helechos y coníferas del Cretácico Inferior en la cuenca del Valle Superior del Magdalena, Colombia. Boletín de Geología, 38(4): 29-42.
En este trabajo se describen nuevos registros de la macroflora del Cretácico Inferior (Aptiano-Albiano) de Colombia. El material estudiado fue colectado en la Cuenca del Valle Superior del Magdalena en tres localidades del departamento del Tolima, en las formaciones Yaví y Caballos. Los restos fósiles comprenden especímenes de filicopsidas y coníferas. Entre los helechos se encuentra la familia Marsileaceae, un grupo importante identificado en la flora fósil ya que se propone como el primer registro de un helecho acuático para el Cretácico Inferior en Colombia. Adicionalmente, abundantes restos de frondes estériles al menos bipinnadas con pínulas lanceoladas y venación pecopteridea son asignadas al morfogénero Cladophlebis sp. Las coníferas por su parte, presentan estructuras vegetativas asociadas a estructuras reproductivas. Los ejes con hojas más frecuentes fueron asignados al morfogénero Brachyphyllum sp. mientras que las escamas ovulíferas aisladas poseen características morfológicas de tipo Araucarites sp. (Araucariaceae). En conjunto, estos hallazgos constituyen un punto clave para entender la biogeografía y evolución de estos grupos y así mismo contribuyen a ampliar el conocimiento de la flora del Cretácico en Colombia.
Palabras clave: Cretácico, Colombia, Valle Superior del Magdalena, pteridofitas, gimnospermas
NEW RECORDS OF FERNS AND CONIFERS FROM THE LOWER
CRETACEOUS IN THE UPPER MAGDALENA VALLEY BASIN,
COLOMBIA
ABSTRACT
Descriptions of new plant macrofossils from the Lower Cretaceous (Aptian-Albian) of Colombia are presented. The studied material was collected in the Upper Magdalena Valley Basin in three exposing levels from the Yaví and Caballos formations. The fossil remains are associated to filicopsids and conifers. Within the ferns, the family Marsileaceae, constitutes an important group identified in the fossil flora because it is proposed as the first record of aquatic ferns for the Lower Cretaceous of Colombia. Additionally, abundant sterile bipinnate fronds with lanceolate pinnule and pecopteridea venation remains were included in the morphogenus Cladophlebis sp. The studied conifers presented vegetative structures associated to reproductive organs. The most abundant sterile fragments were included within the morphogenus Brachyphyllum sp. and ovuliferous scales of Araucarites sp. (Araucariaceae) type. Together these new findings represent a key aspect to understand the biogeography of the mentioned groups and, simultaneously, contribute to the knowledge of the Cretaceous plant fossils from Colombia.
Keywords: Cretaceous, Colombia, Upper Magdalena Valley, pteridophytes, gymnospermophyta
Las regiones neotropicales poseen la más alta diversidad de plantas del planeta. Esta diversidad está dominada en gran parte por las angiospermas. Sin embargo, a comienzos del Cretácico los bosques neotropicales eran totalmente dominados por pteridofitas y gimnospermas. El registro fósil documenta estos cambios en la diversidad y permite entender la historia de estos grupos y las dinámicas de los ecosistemas a lo largo del tiempo geológico. En regiones tropicales, el registro fósil es desafortunadamente muy escaso en comparación con el de las zonas templadas, especialmente cuando lo comparamos con el del hemisferio norte (Burnham and Johnson, 2004; Miller and Hickey, 2008; Archangelsky and Del Fueyo, 2010; Jaramillo, 2012). No obstante, en los últimos años, en Colombia, los estudios referentes a macrofloras fósiles se han ampliado considerablemente (Doria et al., 2008; Herrera et al., 2008; Gómez- Navarro et al., 2009; Wing et al., 2009; Correa et al., 2010; Carvalho et al., 2011; Martínez et al., 2015). Sin embargo, la gran mayoría de estos estudios se han centrado en familias de angiospermas (Araceae, Arecaceae, Malvaceae, Menispermaceae, Piperaceae, Rhamnaceae). El estudio de los helechos y coníferas, por el contrario es considerablemente menor. Actualmente, la cuenca del Valle Medio del Magdalena en Villa de Leyva y la cuenca del Valle Superior del Magdalena al sur del Tolima son los principales depósitos cretácicos de donde se ha realizado un mayor estudio de estos grupos (Huertas, 1967; Moreno, 1994; Van Wavener et al., 2002; Moreno et al., 2007).
Las coníferas son un grupo de gimnospermas que han dominado los ambientes boscosos del planeta desde el Pérmico (Taylor et al., 2009; Farjon, 2010). Hoy en día, se reconocen ocho familias con alrededor de 70 géneros y 615 especies, de las cuales 200 se encuentran en los trópicos (Farjon, 2010). De esas ocho familias, la única que posee distribución cosmopolita es la Cupressaceae Bartling, mientras familias como las Araucariaceae Henkel y Podocarpaceae Endlicher, se encuentran prácticamente restringidas al hemisferio sur (Dutra e Stranz, 2007; Farjon, 2010). No obstante, esto es sólo una pequeña representación de la gran diversidad de coníferas que existieron durante el Mesozoico a nivel global (Kunzmann et al., 2004; Dutra e Stranz, 2007; Archangelsky and Del Fueyo, 2010; Farjon, 2010). Durante el Cretácico inferior, la flora gondwánica estaba constituida principalmente por las familias Cheirolepidiaceae, Taxodiaceae (hoy Cupressaceae s.l), Podocarpaceae y Araucariaceae (Kunzmann et al., 2004; Archangelsky and Del Fueyo, 2010). Sin embargo, es poco lo que se conoce de esta flora gondwanica en la región norte, donde los estudios son menores comparados con los de las floras de La Patagonia y de la Península Antárctica (Dutra, 2004; Archangelsky and Del Fueyo, 2010; Cantrill and Poole, 2012).
Los helechos, son el segundo grupo de plantas vasculares más diverso del planeta, después de las angiospermas (Schneider et al., 2004; Rothwell and Stockey, 2008). Así mismo, el registro fósil documenta que fueron un grupo abundante y diverso desde el inicio del Carbonífero hasta el final del Jurásico (Rothwell and Stockey, 2008). En el Cretácico, se registra un cambio en la diversidad, generalmente atribuido como consecuencia de la radiación de las angiospermas. No obstante, Schneider et al. (2004) y Mohr et al. (2015) sugieren que al final de este periodo ocurre la diversificación de la mayoría de los grupos de helechos que existen actualmente.
El objetivo de este estudio es describir la flora fósil de la cuenca del Valle Superior del Magdalena de las formaciones Yaví y Caballos a nivel morfológico y taxonómico, con el fin de ampliar el conocimiento de helechos y coníferas del Cretácico en Colombia y evaluar las implicaciones de estos hallazgos en un contexto biogeográfico y evolutivo.
Un total de 93 macrofósiles de coníferas y helechos fueron colectados en tres localidades en las formaciones Yaví (loc: 110017) y Caballos (loc: 110011 y 110014), ambas aflorantes en la cuenca del Valle Superior del Magdalena al sur del departamento del Tolima en Colombia (FIGURA 1).
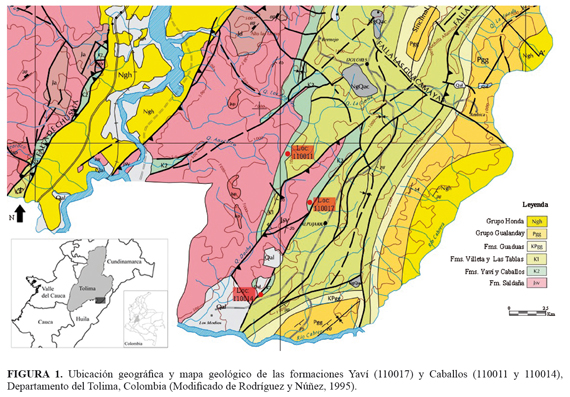
Las formaciones Yaví y Caballos constituyen las unidades estratigráficas más basales de la cuenca Cretácica del Valle Superior del Magdalena. La Formación Yaví, limita en la base con rocas jurásicas de la Formación Saldaña y en el tope con la Formación Caballos. Este límite es considerado ambiguo dado que a lo largo de la secuencia los espesores de ambas formaciones varían y el cambio litológico no es evidente (Guerrero et al., 2000; Velandia et al., 2001; Fuquen y Osorno, 2002). No obstante, hacia el sur de la cuenca, en el municipio de Alpujarra los afloramientos de ambas formaciones son continuos, por lo tanto Guerrero et al. (2000) esclarecen que cerca de Alpujarra se puede establecer el contacto transicional entre las formaciones Yaví y Caballos, con base a la primera aparición de cuarzoarenitas con intercalaciones conglomeráticas de la Formación Caballos (Guerrero et al., 2000); Fuquen y Osorno, 2002).
La Formación Yaví, ha sido datada como Aptiano inferior-medio a partir de polen, esporas, dinoflagelados y amonites (Vergara and Prössl, 1994). La Formación Caballos, por su parte, ha sido datada como Aptiano superior-Albiano, en base a polen y foraminíferos (Vergara and Prössl, 1994; Mejía-Velásquez et al., 2012).
Los restos fósiles de helechos fueron colectados en la Formación Yaví y los de coníferas provienen del segmento 2 de la Formación Caballos e incluyen impresiones tanto estériles como fértiles, cuya preservación no incluye caracteres cuticulares. La observación de los restos macrofósiles se realizó en el Museo Paleontológico Egidio Feruglio (Trelew, Patagonia, Argentina) con un estereoscopio Zeiss Stemi SV 11 con cámara clara incorporada. Para la toma de medidas se usó un calibre digital de 0-150 mm Mitutoyo y para mediciones sobre fotografías se usó el programa Image J. Para las fotografías se utilizó una cámara Canon Eos Rebel T2i/550D de 18 megapíxeles y para el procesamiento y ensamblaje de láminas se usó el programa Adobe Photoshop CS5. El material estudiado se encuentra depositado en la colección paleobotánica del Museo de Historia Natural de la Universidad de los Andes, Bogotá, Colombia (ANDES-Paleo 181-274) y la información de cada espécimen se encuentra en la base de datos del Instituto Smithsonian de Investigaciones Tropicales (STRI).
División - Pteridophyta
Filicopsida Insertae sedis
Morfogénero - Cladophlebis (Brongniart, 1849) FIGURA (2, A-D)
Especie tipo - Cladophlebis albertsii (Dunker) (Brongniart, 1849)
Especímenes: ANDES-Paleo 181-223 (STRI 31289, 31290, 31292, 31294, 31295, 31297-99, 31305-18, 31320-24, 34042-44, 34046, 34048, 34054, 34646, 36759-61, 38993-97).
Edad: Aptiano inferior-medio (Formación Yaví)
Descripción: fragmentos de frondes al menos bipinnadas, con medidas de 2,5-6 cm de largo (incompleto) y de 3-12 cm de ancho (FIGURAS 2A-D). Presenta un raquis recto en la mayoría de los casos, con medidas de 4-8 mm de ancho en la base y disminuye hacia el ápice, siendo surcado por finas estrías longitudinales (FIGURA 2A). La disposición de las pinnas es generalmente opuesta o sub-opuesta en la base del fronde y alternas hacia el ápice (FIGURAS 2A, 2D). Se insertan en ángulos de entre 60° y 90° aumentando hacia la base (FIGURA 2A). El ancho del raquis secundario (de 0,32 mm a 1,58 mm), decrece hacia el ápice. La distancia entre pinnas adyacentes es de 0,3 mm a 1 cm. Las pinnas adyacentes se encuentran levemente imbricadas hacia la porción basal del fronde (FIGURA 2A). Las pínulas tienen forma elíptica (distales) u ovada (proximales) y margen entero (FIGURA 2C). Se disponen de forma opuesta o sub-opuesta en la parte proximal de la pinna y alterna en la parte distal y se insertan perpendicularmente al raquis (FIGURA 2C).

Las pínulas con mejor preservación presentaron medidas de 1-6 mm de largo por 0,9-2 mm de ancho. Las pínulas contiguas se encuentran levemente imbricadas. La vena media se extiende de forma recta hasta los tres cuartos de la pínula bifurcándose en el ápice una única vez (FIGURAS 2B, 2C).
Las venas secundarias presentan disposición sub-opuesta con ángulos que varían entre 25° y 60° siendo en algunos casos los más agudos del lado distal de la pínula. La mayor parte de las venas secundarias se bifurcan al menos 2 veces. La primera división ocurre cerca de la vena primaria donde en algunos casos ocurre anastomosis (FIGURA 2C). Las segundas o terceras bifurcaciones -cuando ocurren- se llevan a cabo cerca del margen de la pínula, donde en algunas venas se forma anastomosis (FIGURA 2C).
Discusión y comparaciones: Cladophlebis se asigna sobre la base de restos de frondes estériles al menos bipinnadas con pínulas lanceoladas insertadas totalmente desde la base y con venación cladophleboide, que no pueden ser asignados a un grupo natural debido a la ausencia de caracteres reproductivos (Escapa and Cúneo, 2012).
Cladophlebis fue propuesto por Brongniart (1849) para designar únicamente frondes estériles de aspecto filiforme del Mesozoico. Ha sido alternativamente relacionado con distintas familias, como Osmundaceae, Matoniaceae, Cyatheaceae y Schizaeaeceae (Skog, and Dilcher, 1994; Tidwell and Ash, 1994). La incierta afinidad sistemática de este morfogénero sugiere el desarrollo de morfologías vegetativas homoplásicas en distintas familias de helechos (Escapa and Cúneo, 2012). No obstante, las afinidades de frondes del tipo Cladophlebis comúnmente se sugieren como posibles Osmundaceae puesto que se han registrado frondes filiformes en asociación con esporangios de tipo Osmundaceae en especies de Todites (Cladophlebis) y con restos de Osmundacaulis (Miller, 1971).
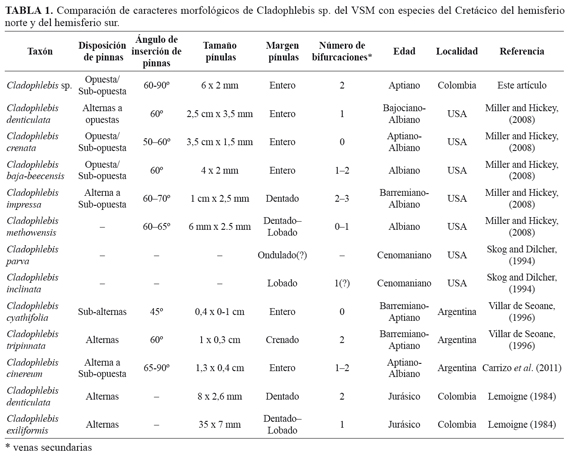
Los materiales descritos de Cladophlebis sp. de la cuenca del Valle Superior del Magdalena son comparados con registros del Cretácico Inferior a nivel global (TABLA 1).
Familia - Marsileaceae Mirbel FIGURA (3, A-C)
Especímenes: ANDES-Paleo 224-231 (STRI 31288, 31296, 34049-53).
Edad: Aptiano inferior-medio (Formación Yaví)
Descripción: fragmentos de foliolos aislados de 1,3 cm a 2 cm de largo y de 1,5 cm a 4 cm de ancho (FIGURAS 3A, 3B). En las muestras mejor preservadas se puede observar que el margen es entero (FIGURAS 3A, 3C). La forma del foliolo es flabelada, de ápice redondo y base truncada (FIGURA 3A). No se demarca una vena principal sino varias venas que se originan de una basal. (FIGURA 3B).

Las venas son sinuosas y se extienden hasta el margen distal. Los ángulos de bifurcación son cerrados hasta rectos. Algunas venas se dividen cerca al margen mientras que otras lo hacen tanto en el sector medio como cerca de la base de la lámina (FIGURA 3B). En todas las muestras estudiadas se han registrado dos o más bifurcaciones en la gran mayoría de las venas (FIGURA 3C). Algunas de las venas forman anastomosis en el tercio distal de la lámina formando areolas elípticas y obovadas (FIGURA 3C). Vena marginal ausente.
Discusión y comparaciones: el nuevo material del VSM, asignado a Marsileaceae se refiere a restos estériles de un foliolo con venación dicotómica, anastomosis parcial, ausencia de vena media y presencia o ausencia de una vena marginal colectora (Nagalingum, 2007; Hu et al., 2008; Hermsen et al., 2014).
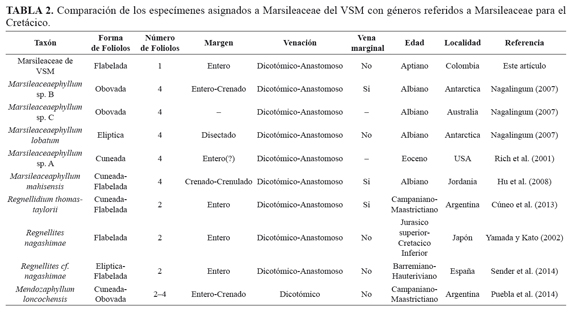
Marsilea johnhallii del Cretácico de USA fue descrito inicialmente por Skog and Dilcher (1992) para designar restos de foliolos de afinidad a Marsileaceae a partir al hallazgo de rizomas y peciolos con cuatro foliolos terminales de margen crenado a entero, venación dicotómica, formación de anastomosis, presencia de una vena marginal y sin estructuras fértiles en conexión orgánica. Posteriormente Nagalingum (2007), sugirió que restos vegetativos de tipo Marsileaceae que no han sido hallados en asociación repetida con esporocarpos deben ser incluidos en el morfogénero Marsileaceaephyllum (Hu et al., 2008; Taylor et al., 2009). Sin embargo Hermsen et al. (2014), proponen que muchos de los caracteres diagnósticos para asignar especímenes a Marsileaceaphyllum son muy amplios y se utilizan de manera ambigua, por lo que limitan la diagnosis para Marsileaceaephyllum únicamente a frondes estériles con cuatro foliolos de forma obovada a obdeltoide, y especímenes que sean hallados con menor número de foliolos y patrones de venación con poca preservación es recomendable designarlos como cf. Marsileaceae (Hermsen et al., 2014). Los patrones de venación observados en el nuevo material del VSM y en conjunto con comparaciones con otros foliolos de géneros referidos a Marsileaceae para el Cretácico (TABLA 2) permiten proponerlo como un representante fósil de esta familia en Colombia.
División - Gimnospermae
Coniferales Insertae sedis
Morfogénero - Brachyphyllum Brongiart, 1828 emend. (Harris 1979) FIGURA (4, A-C)
Especie tipo - Brachyphyllum mamillare (Braun) Linder et Hutton
Especímenes: ANDES-Paleo 232-259 (STRI 27196- 99, 27200, 27202, 27209, 27211, 27213, 27219, 27223- 25, 27230-34, 29947-49, 29951-54, 29957, 29958).
Edad: Albiano inferior (Segmento 2 de la Formación Caballos)
Descripción: fragmentos vegetativos de al menos tres ordenes de ramificación (FIGURAS 4A, 4B). Eje primario de 2-8 cm de largo y 1,5-3 mm de ancho (con hojas), con crecimiento aparentemente ortotrópico. Eje secundario (incompleto) de 2-2,5 cm de largo y 2-4 mm de ancho (con hojas) se inserta en su mayoría en ángulos de 60° y con disposición alterna (FIGURA 4A). La distancia entre ramas adyacentes es de hasta 2 cm.

El tercer orden de ramificación de 0,6-1,5 cm de largo y 2-3 mm de ancho se inserta en ángulos rectos (hacia la base) y agudos (hacia el ápice) con disposición entre opuesta a sub opuesta (FIGURA 4A).
Hojas persistentes, de margen entero, imbricadas, con disposición helicoidal y generalmente adpresas al eje primario, secundario y terciario (FIGURAS 4B, 4C). De apariencia plana y contorno generalmente romboide (las de la base suelen ser de aspecto circular), levemente más anchas (hasta 1,6 mm) que largas (hasta 1,3 mm) en los ejes secundario y terciario, y más largas (2,19 mm) que anchas (1,23 mm) en el eje primario (FIGURA 4C). Parastiquias de contacto 5/7 en la rama de primer orden y 5/6 en la de segundo y tercer orden (FIGURA 4C). Las hojas del eje primario tienen ápice acuminado y base redonda, las del segundo y tercer eje, presentan ápice y base convexas y obtusas (FIGURAS 4A, 4B, 4C).
Discusión y comparaciones: Brachyphyllum es un morfogénero que se asigna a restos estériles de coníferas del mesozoico que presentan hojas romboides a ovaladas, imbricadas y dispuestas en espiral. La distinción con el morfogénero Pagiophyllum radica en la relación entre el ancho y el largo de la hoja, que en Brachyphyllum es más ancha que larga, mientras que en Pagiophyllum es más larga que ancha (Harris, 1979).
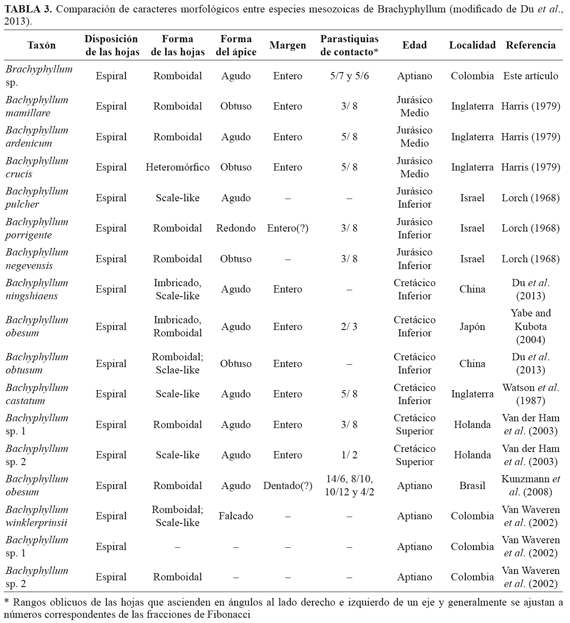
Brachyphyllum fue definido por Brongniart inicialmente en 1828, posteriormente Linder and Hutton (1836) describieron la especie tipo Brachyphyllum mamillare, haciendo énfasis en caracteres cuticulares de las estructuras vegetativas (Harris, 1979). Restos vegetativos de tipo Brachyphyllum han sido alternativamente relacionadas con varias familias de coníferas como Cupressaceae, Taxodiaceae (hoy Cupressaceae s.l), Podocarpaceae, Cheirolepidiaceae y Araucariaceae (Van der Hammen and Van Konijnenburg-Van Cittert, 2003; Du et al., 2013) siendo comúnmente asignado por varios autores a las dos últimas familias (Harris, 1979; Kunzmann et al., 2004). El material de Brachyphyllum sp. del VSM es comparado con varias especies mesozoicas a escala global (TABLA 3).
Familia Araucariaceae Henkel y Hochst, 1865
Morfogénero - Araucarites Presl, 1838 FIGURA (5, A-C)
Especie tipo - Araucarites goeppertii Presl, en Sternberg, 1838
Especímenes: ANDES-Paleo 260-274 (STRI 27189, 27193, 27201, 27204, 27211, 27212, 27215, 27217, 27221, 27228, 27235, 29947, 29951, 29958). Edad: Albiano inferior (Segmento 2 de la Formación Caballos)
Descripción: Complejos ovulíferos aislados de 8,5 a 17 mm de longitud. La porción más amplia se encuentra en el lado distal de entre 8 a 13 mm, disminuyendo gradualmente hacia la parte proximal, de 4 a 6 mm de ancho (FIGURA 5A-C). De forma obovada, ápice mucronado y base truncada. Un único ovulo obovado se encuentra embebido en la región media de los complejos con medidas variables de entre 6 a 12 mm de largo y 4 a 6 mm de ancho en la parte distal (FIGURA 5A-C). Las alas del complejo ovulífero, de 2-2,5 mm de ancho, se encuentran a cada lado del óvulo (FIGURAS 5-C).
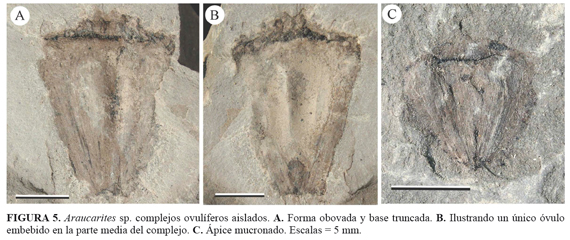
Discusión y comparaciones: el morfogénero Araucarites incluye complejos ovulíferos aislados de afinidad Araucariaceae (Zijlstra and Van Konijnenburg- Van Cittert, 2000; Rees and Cleal, 2004). Varios autores han propuesto incluir únicamente complejos aislados que no presenten lígula como Araucarites y dentro de Araucaria cuando la lígula es visible (Rees and Cleal, 2004). Sin embargo, la presencia o ausencia de lígula radica principalmente tanto de la cara (abaxial/adaxial) que se esté observando como de la preservación del espécimen (Rees and Cleal, 2004). En este trabajo se hace uso del nombre Araucarites como sugiere Rees and Cleal (2004) para designar únicamente complejos ovulíferos aislados de afinidad Araucariaceae y que no presentan una lígula visible.
La morfología estudiada aquí de Araucarites sp. es semejante a otros especímenes del mesozoico para ambos hemisferios. Con la especie Araucarites phillipsii del Jurásico de Inglaterra (Harris, 1979) se asemeja en cuanto a tamaño y proporciones de las alas que se disponen a cada lado de la semilla y se diferencia sutilmente en tamaño del complejo ovulífero y de la semilla. Con la especie Araucarites cf. cutchensis (Rees and Cleal, 2004) del Jurásico de la Antárctica comparte caracteres como dimensiones y proporciones generales de la semilla, forma mucronada del ápice y forma ovada-obovada de la semilla. Con la especie cretácica de Brasil cf. Araucaria sp. (Kunzmann et al., 2004) es similar en forma y tamaño del complejo ovulífero, y se diferencia principalmente en que la especie de Brasil presenta un conjunto de 20 estrías características del complejo, las cuales no están presentes en la especie de Colombia, adicionalmente no hay evidencia de un único óvulo embebido en el complejo, lo cual es el carácter más importante de las especie del VSM.
Los restos fósiles hallados en los depósitos de la cuenca del Valle Superior del Magdalena comprenden 93 fragmentos de helechos y coníferas, asignables a tres morfogéneros: Cladophlebis, Brachyphyllum, y Araucarites, y uno a nivel de familia (Marsileaceae).
Los únicos registros previos de macrofósiles en las formaciones Yaví y Caballos fueron descritos por Moreno (1994). Estos restos corresponden a frondes estériles de Pteridophyta, asignadas al género Weichselia Stiehler, ramos y hojas de Cycadophytas con afinidad al género Pseudoctenis Seward y órganos vegetativos de coníferas con afinidad a los géneros Podozamites Braun y Cupressinocladus Seward. Para los restos de angiospermas no se realiza ninguna identificación formal dada la mala preservación del material. Adicionalmente, los autores hacen referencia a abundantes troncos de coníferas asociados a restos vegetativos. Todos los especímenes descritos por Moreno (1994) fueron colectados en las formaciones Caballos (antes Fm. Alpujarra) y El Ocal. Estudios palinológicos también han sido realizados para las formaciones Yaví y Caballos, principalmente se resaltan aquellos por Vergara and Prössl (1994) y Mejía-Velásquez et al. (2012), en los que se ha reportado mayor variedad de helechos, coníferas, gnetales y algunas angiospermas.
El hallazgo de foliolos de tipo Marsileaceae en la Formación Yaví representaría el registro más antiguo conocido para la familia en Colombia. Adicionalmente, los restos acá asignados a Cladophlebis, Brachyphyllum y Araucarites constituyen nuevos ocurrencias para la macroflora de la cuenca del Valle Superior del Magdalena.
Implicaciones biogeográficas y evolutivas
El registro de macrofósiles de la familia Marsileaceae es bastante escaso comparado con el de microfósiles. El registro más antiguo pertenece a Regnellites nagashimae Yamada et Kato del Jurásico Superior de Japón (Yamada and Kato, 2002). No obstante, los registros de Marsileaceae aumentan en frecuencia hacia el Cretácico Inferior (Nagalingum, 2007; Hu et al., 2008; Cúneo et al., 2013; Hermsen et al., 2014; Puebla et al., 2014; Sender et al., 2014). En Suramérica, se tiene registro de la familia para el Cretácico Superior de Argentina con Regnellidium thomas-taylorii Cúneo, Gandolfo et Hermen (Cúneo et al., 2013), Mirasolita irupensis Hermsen, Gandolfo et Cúneo (Hermsen et al., 2014) y Mendozaphyllum loncochense Puebla, Prámparo et Gandolfo gen. nov. (Puebla et al., 2014). El nuevo material del VSM, asignado a Marsileaceae difiere en cuanto a caracteres morfológicos del foliolo con las especies Argentinas. Por el contrario, con las especies Regnellites nagashimae de Japón (Yamada and Kato, 2002) y Regnellites cf. nagashimae de España (Sender et al., 2014) presenta mayores similitudes.
Cladophlebis, contrario a Marsileaceae, posee un registro fósil abundante para el Mesozoico a nivel global, siendo bastante frecuente en casi todo Gondwana (Carrizo et al., 2011). Cladophlebis ha sido atribuido a distintas familias de helechos como Cyatheaceae, Osmundaceae y Dennstaetiaceae (Villar de Seoane, 1996; Phipps et al., 1998) a partir del hallazgo de órganos reproductivos en conexión orgánica. Sin embargo, las frondes y pinnas descritas para Cladophlebis sp. del VSM podrían pertenecer a cualquiera de estas familias, dada la ausencia de órganos reproductivos en las formaciones de estudio.
Cladophlebis sp. del VSM presenta caracteres que se asemejan a la especie Cladophlebis baja-beecensis del Albiano de USA (Miller and Hickey, 2008) y difiere notablemente de las especies descritas para el Cretácico Inferior en Argentina (Villar de Seoane, 1996; Carrizo et al., 2011). De igual manera, es muy distinta de las especies descritas por Lemoigne (1984) para el Jurásico de Colombia.
Los especímenes de Brachyphyllum sp. del VSM presentan caracteres morfológicos semejantes con las especies Brachyphyllum ardenicum (Harris, 1979) y Brachyphyllum castatum (Watson et al., 1987) del Jurásico y Cretácico de Inglaterra, respectivamente. Adicionalmente, es similar con la especie aptiana Brachyphyllum obesum descrita para Brasil (Kunzmann et al., 2004) y para Japón (Yabe and Kubota, 2004). Esto podría indicar que el material del VSM podría ser atribuido como Brachyphyllum obesum, sin embargo Kunzmann et al. (2004) y Yabe and Kubota (2004) utilizan caracteres epidérmicos preservados para la asignación de esta especie y dichos caracteres no están presentes en el material del Valle Superior del Magdalena. Con respecto a las especies aptianas descritas por Van Waveren et al. (2002) de Villa de Leyva, no se puede establecer una comparación clara, dado que los caracteres descritos por los autores no son suficientes.
Araucarites ha sido registrado para ambos hemisferios durante el Mesozoico, principalmente en sedimentos Jurásicos de Inglaterra (Harris, 1979) y la Antárctica (Rees and Cleal, 2004). Para el Cretácico se encuentra ampliamente distribuido en el hemisferios sur, especialmente en floras de Argentina (Archangelsky, 1966), Antártica (Halle, 1913) y Brasil (Kunzmann et al., 2004). En Colombia, restos de tipo Araucarites no han sido reportados para sedimentos mesozoicos. La semejanza entre los morfogéneros descritos de la cuenca del Valle Superior del Magdalena con otros morfogéneros a nivel global pueden ser interpretadas como circunstanciales dada la amplia distribución y gran abundancia de estas formas durante el Mesozoico. Adicionalmente, en Colombia, son pocos los hallazgos y las descripciones de estos morfogéneros, lo que impide establecer comparaciones y relaciones de estos grupos con mayor precisión. No obstante, el material de la cuenca del Valle Superior del Magdalena suma una mejor colección de referencia y descripciones más detalladas de las formas vegetales existente durante el Cretácico Inferior en Colombia.
Los autores agradecen al Museo Paleontológico Egidio Feruglio, a la Corporación Geológica ARES, al Instituto Smithsonian de Investigaciones Tropicales y al Servicio Geológico Colombiano, especialmente a José Arenas por su ayuda con los permisos de exportación. A Jorge Moreno, Javier Luque y Catalina Suarez por colectar los restos fósiles estudiados. Camila Monje agradece a COLCIENCIAS por la beca de jóvenes investigadores e innovadores 2014-2015. A Nicolás Pérez, Aura Cuervo y Helbert García por sus valiosos aportes de geología y a Rosario Romero por la ilustración de Cladophlebis. Finalmente, a Carlos Jaramillo por sus comentarios y a los dos evaluadores, Dr. Ari Iglesias y un evaluador anónimo, por nutrir este estudio con sus críticas y sugerencias.
Archangelsky, S. 1966. New Gimnosperms from Ticó, Santa Cruz Province, Argentina. British Museum Bulletin of Geology, 13: 259-195. [ Links ]
Archangelsky, S., and Del Fueyo, G.M. 2010. Endemism of Early Cretaceous conifers in western Gondwana. In: Plants in Mesozoic time. Morphological Innovation, Phylogeny, Ecosystem. Indiana University Press, Bloomington, pp. 247-268 [ Links ]
Brongniart, A.1849. Tableau des genres de végétaux fossiles considérés sous le point de vue de leur classification botanique et de leur classification. Dictionnaire Universal de Histoire Natural. Imprimerie de L. Martinet. Paris, Vol.13, 127p. [ Links ]
Burnham, R. J., and Johnson, K.R. 2004. South American palaeobotany and the origins of neotropical rainforests. Philosophical Transactions of the Royal Society of London Series B, 359: 1595-1610. [ Links ]
Cantrill, D.J and Poole, I. 2012. The vegetation of Antarctica through geological time. Cambridge University Press. United Kingdom, 189p. [ Links ]
Carrizo, M.A., Del Fueyo, G.M., y Archangelsky, S. 2011. Morfología y Anatomía de un helecho creciendo bajo condiciones de estrés en el Aptiano de Santa Cruz, Argentina. Ameghiniana, 48(4): 605-617. [ Links ]
Carvalho, M.R., Herrera, F.A., Jaramillo, C.A., Wing, S.L., and Callejas, R. 2011. Paleocene Malvaceae from northern South America and their biogeographical implications. American Journal of Botany, 98(8): 1337- 1355. [ Links ]
Correa, E., Jaramillo, C.A., Manchester, S., and Gutierrez, M. 2010. A fruit and leaves of Rhamnaceous affinities from the late Cretaceous (Maastrichtian) of Colombia. American Journal of Botany, 97(1): 71-79. [ Links ]
Cúneo, N.R., Hermsen, E.J., and Gandolfo, M.A. 2013. Regnellidium (Salviniales, Marsileaceae) Macrofossils and Associated Spores from the Late Cretaceous of South America. International Journal of Plant Sciences, 174(3): 340-349. [ Links ]
Doria, G., Jaramillo, C.A., and Herrera, F. 2008. Menispermaceae from the Cerrejón Formation, middle to late Paleocene, Colombia. American Journal of Botany, 95(8): 954-973. [ Links ]
Du, B.X., Sun, B.N., Ferguson, D.K., Yan, D.F., Dong, C., and Jin, P.H. 2013. Two Brachyphyllum species from the Lower Cretaceous of Jiuquan Basin, Gansu Province, NW China and their affinities and palaeoenvironmental implications. Cretaceous Research, 41: 242-255. [ Links ]
Dutra, T.L. 2004. Paleofloras da Antárctica e sua relação com os eventos tectônicos e paleoclimâticos nas altas latitudes do sul. Revista Brasileira de Geociências, 34(3): 401-410. [ Links ]
Dutra, T.L., e Stranz, A. 2009. Biogeografia, evolução e ecologia da familia Araucariaceae: o que mostra a Paleontologia. Em: Floresta com Araucária: Ecologia, Conservação e Desenvolvimento Sustentável. (Fonseca, C.R., Souza, A.F., Leal-Zanchet, A.M., Dutra, T.L., Backes, A., e Ganade, G. Eds.), Holos Editora, pp. 15-33. [ Links ]
Escapa, I.H., and Cúneo, N.R. 2012. Fertile Osmundaceae from the Early Jurassic of Patagonia, Argentina. International Journal of Plant Sciences, 173(1): 54-66. [ Links ]
Farjon, A. 2010. A Handbook of the World's Conifers (2 Vols.). Brill Academic Publishers. pp. 13-21. [ Links ]
Fuquen, J. A., y Osorno, J. F. 2002. Geología de la plancha 303 Colombia. Departamentos de Huila, Tolima y Meta, escala 1:100.000. INGEOMINAS, pp.32-37. [ Links ]
Gómez-Navarro, C., Jaramillo, C.A., Herrera, F., Wing, S.L., and Callejas, R. 2009. Palms (Arecaceae) from a Paleocene rainforest of northern Colombia. American Journal of Botany, 96(7): 1300-1312. [ Links ]
Guerrero, J., Sarmiento, G., and Navarrete, R. 2000. The stratrigraphy of the W side of the Cretaceous Colombia Basin in the Upper Magdalena Valley. Reevaluation of Selected Areas and Type localities including Aipe, Guaduas, Ortega and Piedras. Geologia colombiana, 25: 45-110. [ Links ]
Halle, T.G. 1913. Some Mesozoic plant bearing deposits in Patagonia and Tierra del Fuego and their floras. Kunglinga Svenska Vetensakapakademien Handlingar, 51: 1-58. [ Links ]
Harris, T.M. 1979. The Yorkshire Jurassic Flora V. Coniferales. British Museum of Natural History, London, pp. 4-57. [ Links ]
Hermsen, E.J., Gandolfo, M.A., and Cúneo, N.R. 2014. New marsileaceous fossils from the late Cretaceous of South America and a reevaluation of Marsileaceaephyllum. Plant Systematics and Evolution, 299(7): 369-386. [ Links ]
Herrera, F., Jaramillo, C.A., Dilcher, D., Wing, S.L., and Gómez-Navarro, C. 2008. Fossil Araceae from a Paleocene neotropical rainforest in Colombia. American Journal of Botany, 95(12): 1569-1583. [ Links ]
Hu, S., Taylor, D.W., Brenner, G.J., and Basha, S.H. 2008. A new marsilealean fern species from the Early Cretaceous of Jordan. Paleoword, 17: 235-245. [ Links ]
Huertas, G. 1967. Sertum Florilae Fossilis Villae de Leivae. Caldasia, 9(46): 59-75. [ Links ]
Jaramillo, C.A. 2012. Historia geológica del Bosque Húmedo Tropical. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales, 36(138): 57-80. [ Links ]
Kunzmann, L., Mohr, B.A.R., and Bernardes de Olivera, M.E.C. 2004. Gymnosperms from the Lower Cretaceous Crato Formation (Brazil). I. Araucariaceae and Lindleycladus (incertae sedis). Mitteilung aus dem Museum für Naturkunde Berlin, Geowissenschaftliche Reihe, 7: 155-174. [ Links ]
Lemoigne, Y. 1984. Données nouvelles sur la paléoflore de Colombie. Geobios, 17: 667-690. [ Links ]
Lorch, J. 1988. Some Jurassic conifers from Israel. Journal of the Linnean Society, 61:177-188. [ Links ]
Lindley, J., and Hutton, W. 1836. The fossil flora of Great Britain; or, figures and descriptions of the vegetable remains found in a fossil state in this country. James Ridgway and Sons, 208p. [ Links ]
Martínez, C., Carvalho, M.R., Madriñán, S., and Jaramillo, C.A. 2015. A Late Cretaceous Piper (Piperaceae) from Colombia and diversification patterns of the genus. American Journal of Botany, 102(2): 273-289. [ Links ]
Mejía-Velásquez, P.J., Dilcher, D., Jaramillo, C.A., Fortini, L.B., and Manchester, S. 2012. Palynological composition of a Lower Cretaceous South American tropical sequence: climatic implications and diversity comparisons with other latitudes. American Journal of botany, 99(11): 1819-1827. [ Links ]
Miller, C.N. 1971. Evolution of the fern family Osmundaceae based on anatomical studies. Contributions from the Museum of Paleontology, University of Michigan, 23(8): 105-169. [ Links ]
Miller, I.M., and Hickey, L.J. 2008. The fossil flora of the Winthrop formation (Aptian-Early Cretaceous) of Washington State, USA. Part I. Bryophyta and Pteridophytina. Bulletin of the Peabody Museum of Natural History, 49: 135-180. [ Links ]
Moreno, M. 1994. La Paleoflora del Cretácico Inferior de las regiones de San Antonio y Aipe (Huila). En: Estudios del Valle Superior del Magdalena (Etayo-Serna Ed.). W.Taller Editorial, Bogotá, pp. XIV1-XVI12. [ Links ]
Moreno, M., Gómez, A., y Castillo, H. 2007. Frenelopsis y Pseudofrenelopsis (Coniferales: Cheirolepidiaceae) en el Cretácico Temprano de Colombia. Boletín de Geología, 29(2): 13-19. [ Links ]
Mohr, B.A.R., Bernardes de Oliveira, M.E.C., Loveridge, R., Pons, D., Sucerquia, P.A., and Castro Fernandes, M.C. 2015. Ruffordia goeppertii (Schizaeales, Anemiaceae)- A common fern from the Lower Cretaceous Crato Formation of northeast Brazil. Cretaceous Research, 54: 17-26. [ Links ]
Nagalingum, N. S. 2007. Marsileaceaephyllum, a new genus for marsileaceous macrofossils: leaf remains from the Early Cretaceous (Albian) of Southern Gondwana. Plant Systematics and Evolution, 264: 41-55. [ Links ]
Phipps, C.J., Taylor, T.N., Taylor, E.L., Cúneo, N.R., Boucher, L.D., and Yao, X. 1998. Osmunda (Osmundaceae) from the Triassic of Antarctica: An example of evolutionary stasis. American Journal of Botany, 85(6): 888-895. [ Links ]
Puebla, G.G., Prámparo, M. B., and Gandolfo, M.A. 2015. Aquatic ferns from the Upper Cretaceous Loncoche formation, Mendoza, central-western, Argentina. Plant Systematics and Evolution, 301: 577-588. [ Links ]
Rees, P. M., and Cleal, C. J. 2004. Lower Jurassic Floras From Hope Bay And Botany Bay, Antarctica. Special Papers in Paleontology, 72: 5-90. [ Links ]
Rich, F.J., Johnson, D.M., and Durkin, T.V. 2001. Occurrence and Paleoecology of Marsilea from Eocene Wasatch Formation, Johnson County, Wyoming. Palaios, 16: 608-613. [ Links ]
Rodríguez, G., y Núñez, A. 1995. Mapa geológico generalizado del Departamento del Tolima, escala 1:250.000. INGEOMINAS. [ Links ]
Rothwell, G.W., and Stockey, R.A. 2008. Phylogeny and evolution of ferns: A paleontological perspective. In: Biology and Evolution of Ferns and Lycophytes (Ranker, T.A., Haufler, C.H. Ed). Cambridge University Press, Cambridge, pp. 332-366. [ Links ]
Schneider, H., Schuettpelz, E., Pryer, K.M., Cranfill, R., Magallón, S., and Lupia, R. 2004. Ferns diversified in the shadow of angiosperms. Nature, 428: 553-557. [ Links ]
Sender, L.M., Villanueva-Amadoz, U., Diez, J.B., Fernandez-Baldor, F.T., and Ferrer, J. 2014. Earliest Record of semi-aquatic fern leaves (Family Marsileaceae) in the Mesozoic of Eurasia (Lower Cretaceous, Spain). Cretaceous Research, 51: 241-247. [ Links ]
Skog, J.E., and Dilcher, D.L. 1992. A new species of Marsilea from the Dakota Formation in central Kansas. American Journal of Botany, 79(9): 982-988. [ Links ]
Skog, J.E., and Dilcher, D.L. 1994. Lower vascular plants of the Dakota formation in Kansas and Nebraska, USA. Review of Paleobotany and Palynology, 80:1-18. [ Links ]
Taylor, T.N., Taylor, E.L., and Krings, M. 2009. Paleobotany: The biology and evolution of fossil plants. Academic Press. San Diego, California, 1230p. [ Links ]
Tidwell, W.D., and Ash, S.R. 1994. A review of selected Triassic to Early Cretaceous ferns. Journal of Plant Research, 107(4): 417-442. [ Links ]
Van der Hammen, R.W.J.M., and Van Konijnenburg- Van Cittert, J.H.A. 2003. Rare conifers from the type area of the Maastrichtian (Upper Cretaceous, Southeast Netherlands). Scripta Geologica, 126: 111-119. [ Links ]
Van Waveren, I.M., Van Konijnenburg-Van Cittert, J.H.A., Van der Burgh, J., and Dilcher, D.L. 2002. Macrofloral remains from the Lower Cretaceous of the Leiva region (Colombia). Scripta Geologica, 123: 1-39. [ Links ]
Velandia, F., Núñez, A., y Marquínez, G. 2001. Mapa Geológico del Departamento del Huila, escala 1:300.000. INGEOMINAS, 184p. [ Links ]
Vergara, L., and Prössl, K. 1994. Dating The Yavi Formation (Aptian, Upper Magdalena Valley, Colombia) palynological results. In: Etayo-Serna, F., (ed.). Estudios geológicos del Valle Superior del Magdalena. Universidad Nacional de Colombia. Bogotá, 14p. [ Links ]
Villar de Seoane, L. 1996. Estudio Morfológico de Helechos fértiles hallados en la Formación Baquero (Cretácico Inferior), provincia de Santa Cruz, Argentina. Revista Española de Paleontología, 11: 83-90. [ Links ]
Yabe, A., and Kubota, K. 2004. Brachyphyllum obesum newly discovered thermophilic conifer branch from the Lower Cretaceous Kitadani Formation of the Tetori Group, central Japan. Memoir of the Fukui Prefectural Dinosaur Museum, 3: 23-29. [ Links ]
Yamada, T., and Kato, M. 2002. Regnellites nagashimae gen. et sp. nov., the oldest macrofossil of Marsileaceae, from the Upper Jurassic to Lower Cretaceous of western Japan. International Journal of Plant Sciences, 163(5): 715-723. [ Links ]
Watson, J., Fisher, H.L., and Hall, N.A. 1987. A new species of Brachyphyllum from the English Wealden and its probable female cone. Review of Palaeobotany and Palynology 51, 169-187. [ Links ]
Wing, S.L., Herrera F., Jaramillo, C., Gomes-Navarro, C., Wilf, P., and Labandeira, C.C. 2009. Late Paleocene fossils from the Cerrejón Formation, Colombia, are the earliest record of Neotropical rainforest. Proceedings of the National Academy of Sciences, 106(44): 18627- 18632. [ Links ]
Zijlstra, G., and Van Konijnenburg-Van Cittert, H. 2000. Proposal to conserve the name Araucarites C. Presl (Fossil Gymnospermae, Coniferales, Araucariaceae) against Araucarites Endl. (Gymnospermae, Coniferales). Taxon, 49: 279-280. [ Links ]
Trabajo recibido: mayo 11 de 2016
Trabajo aceptado: septiembre 23 de 2016
Manuscrito publicado en internet: septiembre 29 de 2016

