











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google 
 Permalink
Permalink
INTRODUCCIÓN
Los ictiosaurios fueron los primeros amniotas secundariamente acuáticos en desarrollar un cuerpo en forma de pez, con extremidades consideradas como características únicas del grupo, adaptadas a una forma eficiente de locomoción en el agua y representadas por fenómenos de hiperfalangia e hiperdactilia (Motani, 2005; Maisch y Matzke, 2000; Maxwell et al., 2014).
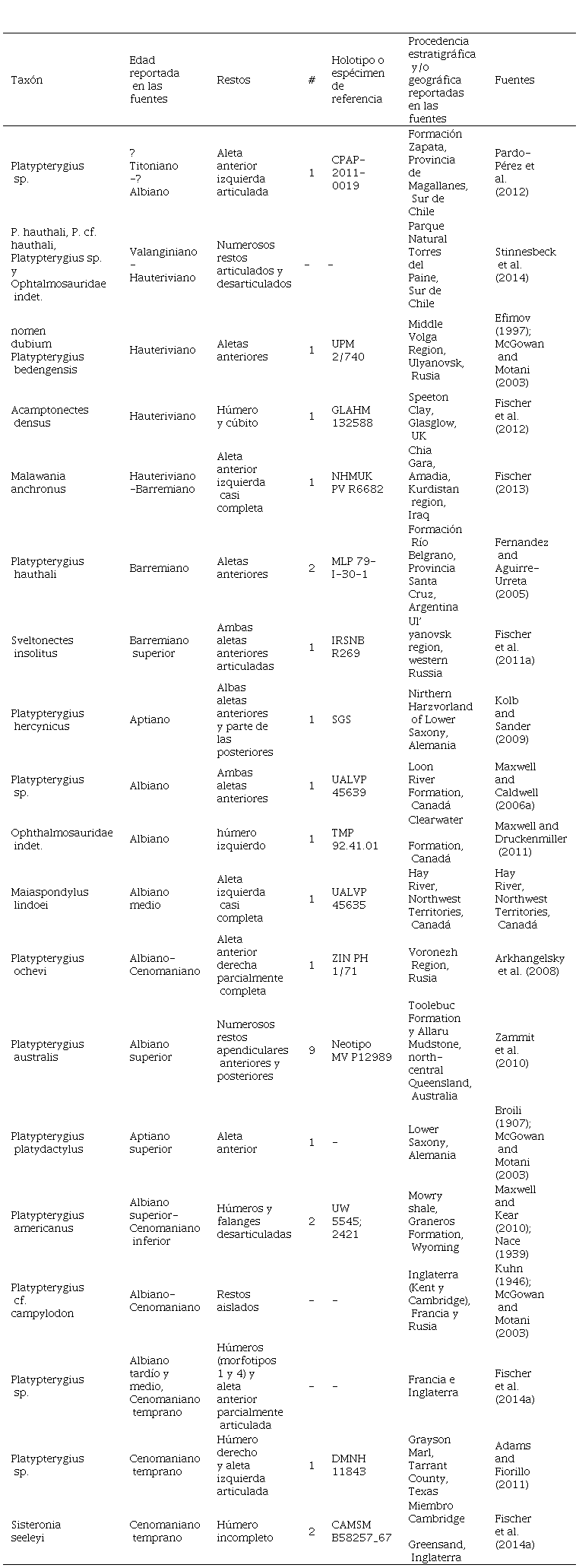
El registro fósil muestra que la preservación de los restos apendiculares de los ictiosaurios cretácicos no es muy frecuente y en especial la de los miembros posteriores (Motani, 1999a). Las publicaciones sobre restos de apéndices anteriores procedentes de rocas cretácicas reportan algunos especímenes articulados y muchos restos aislados, todos ellos incluidos en Ophthalmosauridae (ver TABLA 1).
TABLA 1 Listado de especímenes con restos del esqueleto apendicular preservados reportados para el Cretácico Inferior. P. ochevi y P. campylodon son considerados como species inquirenda por Fischer et al. (2016).
El nombre Ophthalmosauridae, inicialmente propuesto por Baur (1887) para incluir a OphthalmosaurusSeeley, 1874 (Baptanodon en la versión inglesa de su publicación según McGowan y Motani, 2003), fue utilizado por Andrews (1910) en su descripción detallada de Ophthalmosaurus. En trabajos de revisión posteriores (Motani, 1999b; Maisch y Matzke, 2000; McGowan y Motani, 2003) se proponen definiciones o diagnosis de Ophthalmosauridae (u Ophthalmosauria) que incluyen rasgos craneanos y apendiculares.
En la revisión de McGowan y Motani (2003) se consideran válidos 7 géneros para la familia Ophthalmosauridae: Ophthalmosaurus, MollesaurusFernández, 1999; Nannopterygiusvon Huene, 1922; UndorosaurusEfimov, 1999; Brachypterygiusvon Huene, 1922; CaypullisaurusFernández, 1997; AegirosaurusBardet y Fernández, 2000; del Jurásico y Platypterygiusvon Huene, 1922 del Cretácico. Después de la revisión de McGowan y Motani (2003) se han publicado numerosos artículos sobre nuevos especímenes, nuevos taxones, revisiones y análisis filogenéticos de esta familia (Maxwell y Caldwell, 2006a; Maxwell y Caldwell, 2006b; Maxwell, 2010; Druckenmiller y Maxwell, 2010; Fischer et al., 2011a; Fernández y Maxwell, 2012; Druckenmiller et al., 2012; Fischer, 2012; Fischer et al., 2012; Roberts et al., 2014; Fischer et al., 2014a; Fischer et al., 2014b; Ji et al., 2016; Paparella et al., 2017; Maxwell et al., 2016; Fischer et al., 2016). Un panorama de estos aportes, con sus respectivas referencias, se expone sintéticamente en la revisión de las ocurrencias de especímenes de oftalmosáuridos presentada por Fernández y Campos (2015). Dentro de la familia Ophthalmosauridae se reconocen actualmente 19 géneros, muchos de ellos monoespecíficos y representados sólo por su holotipo, de edades que comprenden desde el Jurásico medio (Bajociano) hasta el Cretácico superior (Cenomaniano) (Fernández y Campos, 2015; Fischer et al., 2016). En Ophthalmosauridae se incluyen numerosos especímenes de Europa y otros no tan abundantes de Norteamérica, Centroamérica, Suramérica y Australia (Fernández y Campos, 2015; Fischer et al., 2016).
Las últimas propuestas filogenéticas que incorporan los nuevos hallazgos y dan una definición o una diagnosis de la familia Ophthalmosauridae son las de Fischer et al. (2012) y Ji et al. (2016). En el trabajo de Fischer et al. (2012) se reconocen y se redefinen dos subfamilias previamente sugeridas, Ophtalmosaurinae Baur, 1887 y Platypterygiinae Arkhangelsky, 2001, las cuales, según dichos autores, representan la radiación de los miembros de la familia Ophthalmosauridae ocurrida antes del Cretácico. Estas subfamilias son acogidas en varias publicaciones posteriores (Fischer et al., 2013a; Fischer et al., 2014a; Fischer et al., 2014b; Roberts et al., 2014; Druckenmiller y Maxwell, 2014; Zverkov et al., 2015). Sin embargo, en algunos análisis filogenéticos no se reconocen subgrupos dentro de Ophthalmosauridae (Fernández y Talevi, 2014; Maxwell et al., 2015; Ji et al., 2016). En el trabajo de Ji et al. (2016) se ofrecen las más recientes diagnosis de los géneros de la familia Ophthalmosauridae.
Hasta hace pocos años, el género Platypterygius fue reconocido como el único género de Ophthalmosauridae del Cretácico, con una amplia distribución paleogeográfica (Sander, 2000). Con los recientes y abundantes hallazgos, se ha hecho evidente que la familia alcanzó una alta diversidad genérica en el Cretácico (Fernández y Campos, 2015). Además, como lo resalta Zammit (2012), tres géneros del Jurásico superior alcanzaron el Cretácico: Ophthalmosaurus con registros del Calloviano al Berriasiano (Fischer et al., 2012; Zverkov et al., 2015), Aegirosaurus del Titoniano al Valanginiano superior (Fischer et al., 2011b) y Caypullisaurus del Titoniano al Hauteriviano-Barremiano (Fernández, 2001; Fernández y Aguirre-Urreta, 2005; Pardo-Pérez et al., 2011).
Los restos de ictiosaurios reportados para Colombia son numerosos y corresponden todos a especímenes cretácicos (Páramo-Fonseca, 2015). En su mayoría son restos de cráneos y de esqueletos axiales aún no publicados (Páramo-Fonseca, 2015). El material estudiado hasta el momento incluye dos cráneos parcialmente completos encontrados en capas del Barremiano-Aptiano de Villa de Leiva, Boyacá, que fueron incluidos en dos géneros de Ophthalmosauridae: Platypterygius sachicarumPáramo, 1997 y Muiscasaurus cathetiMaxwell et al., 2015.
Este artículo ofrece el primer reporte de la ocurrencia de restos apendiculares de ictiosaurio en Colombia. Su propósito es describir los restos apendiculares anteriores de un ictiosaurio oftalmosáurido encontrados en rocas del Barremiano inferior de Villa de Leiva, Boyacá, Colombia (FIGURA 1), los cuales se han incluido en Ophthalmosauridae por presentar una faceta extrazeugopodial anterior en el húmero, un elemento extrazeugopodial anterior al radio y un dígito distal a éste y por carecer de muescas en los elementos de las aletas anteriores, rasgos considerado diagnóstico para Ohthalmosauridae por McGowan y Motani (2003), Fischer et al. (2012) y Ji et al. (2016).
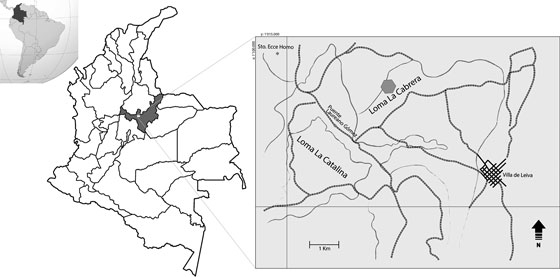
FIGURA 1 Mapa de localización del sector en que se encontró el espécimen FCG-CBP-87El polígono gris indica el sector del hallazgo. Modificado de Etayo-Serna (1968) y Páramo-Fonseca et al. (2016)
Abreviaturas anatómicas
cd2, cd3, carpales distales 2 y 3; cde, cúbito derecho; cdl, cresta deltopectoral del húmero; ci, cúbito izquierdo; co, costillas; dg2, dg3, dg4, dg5, dígitos 2, 3, 4 y 5; e, elemento extrazeugopodial anterior; fc, faceta cubital del húmero; fe, faceta para el elemento extrazeugopodial anterior del húmero; fr, faceta radial del húmero; Fcu, faceta para el cubital; Fh, faceta para el húmero; Fi, faceta para el intermedio; Fp, faceta para el pisiforme; Fr, faceta para el radio; h, húmero; mc2, mc3, metacarpales 2 y 3; m?, posibles metacarpales o carpales distales; p, pisiforme; pd, proceso dorsal del húmero; pr1, pr2, dígitos preaxiales 1 y 2; pt1, pt2, dígitos postaxiales 1 y 2; r, radio; ra, radial; t, tubérculo.
MATERIALES Y MÉTODOS
El espécimen FCG-CBP-87 fue colectado en la región denominada “Loma La Cabrera”, al occidente de la población de Villa de Leiva, Boyacá (FIGURA 1). La ubicación puntual del sitio de hallazgo es desconocida, por lo cual la procedencia estratigráfica del espécimen es imprecisa. En la región del hallazgo afloran rocas del Cretácico Inferior que corresponden a las denominadas “Arcillolitas Abigarradas” de la Formación Paja (Etayo-Serna, 1968; Forero y Sarmiento, 1985), cuya edad ha sido determinada como Barremiano-Aptiano (Etayo-Serna, 1968; Patarroyo, 2000; Patarroyo, 2009). En el proceso de preparación de los restos apendiculares se encontraron dos amonitas junto a los restos óseos, las cuales fueron extraídas y preparadas. Una de ellas (FCG-CBP-87/13-09-14/2) fue conferida a Nicklesia pulchella (D ́Orbigny, 1841) (ver Vermeulen, 2003. P.140, con sinonimia cf. Bürgl 1956, pl VI, fig. 6) (Dr. Fernando Etayo-Serna, comunicación verbal). Su presencia permite concluir que las capas portadoras de los restos apendiculares pertenecen al Barremiano inferior (Dr. Fernando Etayo-Serna, comunicación verbal).
El espécimen comprende los restos apendiculares de las dos aletas anteriores de un ictiosaurio, dispuestas unidas una sobre la otra, junto con algunos fragmentos de costillas (FIGURAS 2, 3). Los restos fueron hallados en una concreción calcárea y exhiben una excelente preservación del material óseo. La muestra fue preparada en el laboratorio del Centro de Investigaciones Paleontológicas (CIP), por el técnico Fredy H. Parra, a través de protocolos mecánicos y químicos. El fósil fue sometido a una tomografía axial computarizada con el fin de rastrear restos óseos en la matriz que ocupa el espacio entre las dos aletas. Esta tomografía fue tomada en los equipos del Hospital Universitario San Ignacio de Bogotá con la colaboración del Dr. Luis Felipe Uriza. Las imágenes de la tomografía se visualizaron en IQ-view y se modelaron en Osirix Lite 2015 e InVesalius 3.1.1. (public license 2017) (FIGURAS 4A, 4B). Con base en las imágenes de la tomografía se orientó la preparación de algunos detalles, los cuales fueron ejecutados por una de las autoras (D. Cortés). Las ilustraciones del espécimen se realizaron sobre la base de fotografías y de las imágenes obtenidas en la tomografía.

FIGURA 2 Espécimen FCG-CBP-87. Fotografía y esquema interpretativo de la aleta izquierda en vista dorsal. Las flechas indican las filas de falanges hexagonales. Las líneas de puntos representan la interpretación de los sectores cubiertos por sedimento u otro hueso y las líneas de trazos indican la reconstrucción de los huesos. Barra de escala: 5 cm.
Los trabajos de McGowan y Motani (2003), Fischer et al. (2012) y Ji et al. (2016), fueron la base principal para abordar la identificación del espécimen. La determinación taxonómica partió de una extensa comparación de la morfología del ejemplar con las descripciones presentadas para los distintos géneros y especies de Ophthalmosauridae y se apoyó en los análisis filogenéticos de los ictiosaurios presentados por Motani (1999a), Maisch y Matzke (2000), McGowan y Motani (2003), Ji et al. (2016) y de los ictiosaurios cretácicos presentada en Fischer et al. (2016). Los estados de los caracteres apendiculares utilizados en este último trabajo fueron establecidos para FCG-CBP-87.
PRESERVACIÓN Y CONSIDERACIONES TAFONÓMICAS
En el espécimen se conservan dos aletas anteriores de un mismo individuo dispuestas unidas, una sobre la otra. Los elementos de la aleta izquierda están en su mayoría articulados o muy poco desplazados de su posición anatómica original, mientras que los de la derecha están desarticulados y desordenados (FIGURAS 2, 3, 4A). La superficie formada por el conjunto de los huesos de la aleta izquierda muestra una convexidad dorsal. El húmero carece de su parte más proximal por ruptura, probablemente en el momento de la extracción, y no se encuentra en relación anatómica articulada con los huesos próximos. La superficie dorsal de este hueso está preservada, a diferencia de una parte de la superficie ventral (hacia el margen posterior) que se encuentra rota. Los elementos zeugopodiales de la aleta izquierda se encuentran ligeramente desplazados de las facetas distales articulares del húmero. La mayor parte de las falanges de la aleta izquierda se encuentra bien articulada, aunque ligeramente desplazada dorsoventralmente, cada una con relación a la siguiente en sentido proximodistal, de manera escalonada.
De la aleta derecha se conserva el cúbito y numerosos elementos autopodiales desarticulados. El cúbito se ubica en posición anterior y distal con relación a la ubicación del cúbito izquierdo (FIGURAS 2, 3, 4A). Además del cúbito, se pueden identificar algunos elementos probablemente del carpo o del metacarpo y abundantes falanges de la aleta derecha. La mayoría de estos elementos se sitúan desordenadamente, agrupados en la superficie ventral de la parte media de la aleta izquierda y separados del cúbito (FIGURA 3).
Se preservan once fragmentos de costillas, la mayoría agrupados en la región posterior y ventral de la aleta izquierda (FIGURAS 2, 3, 4). Los ejes longitudinales de los fragmentos se disponen cerca a los dígitos postaxiales de la aleta izquierda y contactan el pisiforme y varios elementos ovoidales de la aleta derecha. Los tres fragmentos más largos, se encuentran agrupados y ubicados en el margen posterior de la aleta izquierda. Su disposición ordenada sugiere que se trata de fragmentos de tres costillas contiguas conservadas en relación anatómica (FIGURA 4B).
La disposición de los huesos en la muestra evidencia una preservación desigual entre la aleta izquierda y la aleta derecha. La relación espacial de las dos aletas una al lado de la otra soporta la hipótesis de que el cadáver del ictiosaurio se depositó de medio lado. La disposición de los fragmentos de costillas apoya esta hipótesis. Los tres fragmentos más largos representan costillas contiguas y se ubican en posición paralela con la aleta izquierda por lo que parecen corresponder a costillas del lado izquierdo del animal, mientras que los fragmentos más cortos y de disposición desordenada, ubicados sobre la superficie ventral de la aleta izquierda, indican su pertenencia al costado derecho del animal.
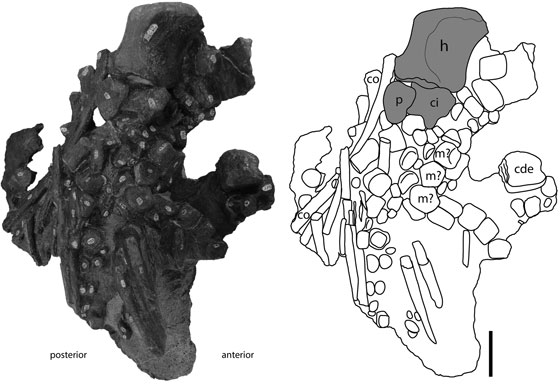
FIGURA 3 Espécimen FCG-CBP-87. Fotografía y esquema interpretativo de la aleta derecha. Barra de escala: 5 cm.
La condición articulada de los elementos de la aleta izquierda, en posición muy próxima a la anatómica, sugiere que ésta aleta fue la que quedó en contacto con el fondo, mientras que la aleta derecha, cuyos elementos se encuentran totalmente desarticulados, fue la que ocupó una posición alta en el cadáver del ictiosaurio. La disposición escalonada que presenta el conjunto de elementos de la aleta izquierda, sin evidencia de deformación tectónica, sugiere que los huesos de esta aleta se acomodaron en esta posición estando ya avanzado el proceso de descomposición de los tejidos blandos y antes del enterramiento. La desarticulación y el leve desplazamiento de los elementos proximales estarían relacionados con la posición ligeramente más alta de esta parte de la aleta, unida al cuerpo. Bajo esta interpretación, el hecho de que los elementos de la aleta derecha se encuentren en contacto con los de la aleta izquierda sugiere la caída por gravedad de los primeros sobre los segundos sin la intervención de corrientes fuertes o depredadores que los disgregaran. La ausencia de corrientes fuertes y de depredadores indica condiciones del fondo tranquilas y anóxicas, lo cual es concordante con las características litológicas de la roca que rodea los restos apendiculares del ictiosaurio (sedimento lodoso gris y ausencia de fauna bentónica). Estas condiciones del fondo favorecieron la buena preservación que exhiben los restos óseos. Un endurecimiento rápido de los sedimentos por precipitación de carbonato de calcio, formando la concreción en que se encontraron los restos óseos facilitó posiblemente la preservación.
SISTEMÁTICA
ICHTHYOSAURIA Blainville, 1835
OPHTHALMOSAURIDAE Baur, 1887
Platypterygiusvon Huene, 1922
Platypterygius sp.
Material referido
FCG-CBP-87. Elementos de dos aletas anteriores de un mismo individuo. Húmero incompleto, zeugopodio y la mayor parte del autopodio izquierdos, cúbito y elementos autopodiales derechos y fragmentos de costillas (FIGURAS 2, 3, 4A, 4B). Muestra depositada en la colección paleontológica del Centro de investigaciones Paleontológicas (CIP), Villa de Leiva, Colombia.
DESCRIPCIÓN
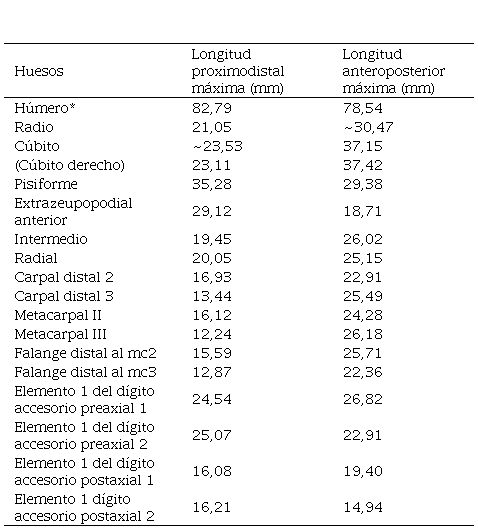
El espécimen FCG-CBP-87 comprende las dos aletas anteriores de un individuo, dispuestas una sobre la cara interna de la otra (FIGURAS 2, 3, 4A). La aleta izquierda, que es la mejor preservada, mide 32 cm a lo largo del eje del dígito IV, sin incluir el húmero. Las dimensiones de los distintos elementos se presentan en la TABLA 2. La lateralidad de las aletas se estableció a partir de la morfología del húmero y su ubicación con respecto a los elementos zeugopodiales adyacentes. La descripción que se presenta a continuación se basa en la morfología de la aleta izquierda, por ser esta la mejor preservada y se complementa con rasgos la aleta derecha cuándo sólo en ésta son visibles.
Húmero- El húmero carece de su parte proximal (FIGURA 5). El extremo distal es robusto, con una longitud anteroposterior de 70 mm y dorsoventral de 46 mm y presenta tres facetas articulares distales: faceta para un elemento extrazeugopodial anterior, faceta radial y faceta cubital (FIGURA 5A). La faceta para el elemento extrazeugopodial anterior es pequeña pero marcadamente cóncava, desviándose anteriormente con respecto a las facetas adyacentes. El borde dorsal de esta faceta es pronunciado (FIGURA 5F). Las facetas radial y cubital son ligeramente cóncavas y se disponen con sus planos de articulación formando un ángulo cercano a los 180º (FIGURA 5A). La faceta cubital es ligeramente más ancha anteroposteriormente que la faceta radial y presenta un borde más amplio que las facetas radial y extrazeugopodial. Posterodistalmente al borde de la faceta cubital se encuentra un tubérculo prominente y cónico (FIGURAS 5B, 5C, 5D). Pese a que el húmero está roto se observa el inicio del proceso dorsal en posición ligeramente anterior al límite entre las facetas radial y cubital (FIGURAS 5A, 5D, 5E). En vista ventral se observa una protuberancia débil próxima al margen anterior del húmero que casi alcanza el borde distal del hueso que corresponde a la cresta deltopectoral (FIGURAS 5B, 5D, 5E).
TABLA 2 Medidas de los huesos diagnósticos de las aletas del espécimen FCG-CBP-87.*Medición del húmero estimada desde el extremo más distal hasta el punto más proximal de la superficie fragmentada. Datos tomados de la aleta izquierda exceptuando el cúbito derecho.
Zeugopodio- Cuatro elementos contactan con el extremo distal del húmero: elemento extrazeugopodial anterior, radio, cúbito y pisiforme (FIGURAS 2, 3). El extrazeugopodial anterior, el radio y el cúbito se encuentran ligeramente girados y desplazados de las facetas articulares del húmero (FIGURAS 2, 3, 4C), pero las longitudes anteroposteriores de los huesos son proporcionales a las de las facetas humerales (20,8 mm, 30,92 mm, 38,16 mm respectivamente).
El radio es ligeramente más pequeño que el cúbito (TABLA 2) y tiene forma pentagonal. Su superficie articular proximal es ligeramente curva y rugosa. Los bordes anterior y posterior son casi rectos y anteriormente presenta dos facetas para la articulación con el radial y con el intermedio. La superficie dorsal del radio es suavemente cóncava, mientras que la superficie ventral es plana. El cúbito derecho, desplazado de su posición original, se observa completo. Tiene contorno pentagonal en vista dorsal y semirectangular en vista ventral (FIGURAS 6A, 6B). Es evidente la presencia de un surco compuesto por dos canalillos que forman una onda bilobular a lo largo de la superficie anterior (FIGURA 6C). La superficie distal está ocupada mayormente por la faceta articular para el cubital (FIGURA 6D). Presenta además una hendidura pronunciada en la superficie posterior (FIGURA 6E).
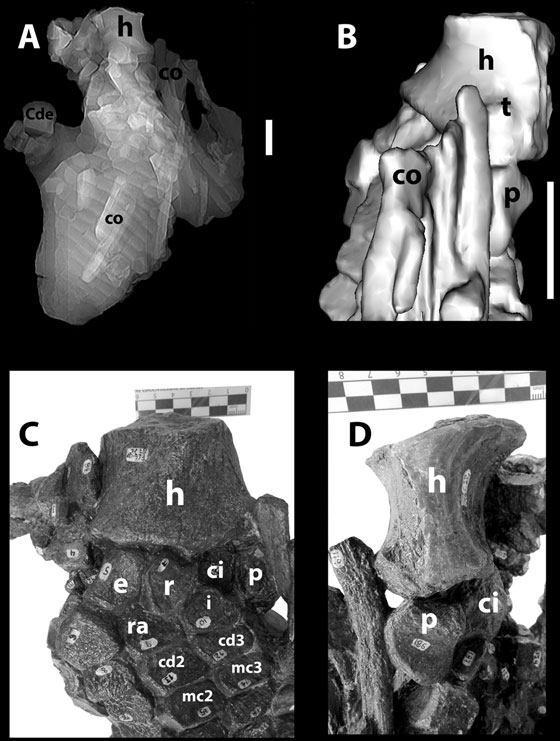
FIGURA 4 Espécimen FCG-CBP-87A. Tomografía Axial Computarizada sin valores de volumen, mostrando la disposición de los elementos de las dos aletas. B. Acercamiento de modelo tridimensional del espécimen en vista posterior, generado en Osirix Lite (public license 2015). C. Fotografía del húmero y elementos próximos en vista anterodorsal, en donde se observa el elemento extrazeugopodial anterior. D. Fotografía del húmero y elementos próximos en vista posteroventral en donde se observa el pisiforme. Barra de escala: 5 cm.
El elemento extrazeugopodial anterior es el más pequeño de los zeugopodiales (FIGURAS 2, 4C). Es alargado proximodistalmente, la superficie anterior tiene forma de “silla de montar” mientras que la posterior es ligeramente convexa. El borde proximal es más largo que el distal. La superficie dorsal es convexa. Presenta cuatro facetas de articulación para unirse proximalmente con el húmero, posteriormente con el radio y el radial y distalmente con el primer elemento del dígito accesorio preaxial I. El pisiforme es más largo que ancho. En su vista ventral tiene proximalmente una elevación aguda ligeramente alargada (FIGURAS 3, 4D, 4E). Su forma desde la vista ventral es subtriangular con ligeras depresiones posteriores y un borde anterior agudo y fuertemente convexo.
Autopodio- El radial tiene forma de ladrillo con margen proximal curvo (FIGURAS 2, 4C). Las facetas posterior y distal son casi planas. La faceta anterior tiene una pequeña hendidura distal, perceptible con el tacto. El intermedio tiene forma pentagonal, siendo más ancho que largo y estrechándose proximalmente sin terminar completamente en punta (FIGURAS 2, 4C). Su superficie dorsal es plana. El intermedio hace contacto proximal con el radio y el cúbito. Las superficies anterior y posterior son más o menos planas y forman ángulo recto con la faceta distal. La anterior hace contacto con el radial y la posterior debió hacerlo con el cubital, no observable. La faceta distal del intermedio es amplia y contacta en su totalidad con el carpal distal 3 (FIGURAS 2, 4C).

FIGURA 5 Fotografías y esquemas interpretativos del húmero izquierdo del espécimen FCG-CBP-87A. Vista dorsal. B. Vista ventral. C. Vista anterior. D. Vista proximal. E. Vista posterior. F. Faceta para el elemento extrazeugopodial anterior con borde protuberante marcado con una flecha. Barra de escala: 2 cm.
Tanto el carpal distal 2 como el carpal distal 3 tienen forma romboidal (FIGURAS 2, 4C). El carpal distal 2 tiene 6 facetas articulares: cuatro amplias (anterior, posterior, proximal y distal) y dos muy pequeñas (anteroproximal y posterodistal), haciendo contacto proximalmente con el radial, posteriormente con el carpal distal 3 y distalmente con el metacarpal 2. Su superficie anterior tiene hendiduras de articulación (FIGURA 2). El carpal distal 3 tiene 5 facetas de articulación (FIGURA 4C). Proximalmente hace contacto con el intermedio, anteriormente con el carpal distal 2 y distalmente con el metacarpal 3. Los carpales distales 2 y 3 continúan las filas (en sentido proximodistal) del radial y del intermedio (FIGURAS 2, 4C).
Los metacarpales tienen contornos menos rectangulares que los carpales (FIGURAS 2, 4C). El metacarpal 2 tiene un margen anterior marcadamente convexo y la superficie anterior es rugosa con hendiduras pronunciadas longitudinalmente. Los márgenes proximal y distal son casi rectos, mientras que el posterior es curvado. El metacarpal 2 es marcadamente masivo y hace contacto proximalmente con el carpal distal 2 y posteriormente con el metacarpal 3. El metacarpal 3 se ve menos masivo que los elementos que lo circundan, tiene 6 facetas de articulación, dos grandes (proximal y distal) y cuatro pequeñas (dos anteriores y dos posteriores). La faceta articular posteroproximal es más larga que la posterodistal. Los metacarpales 2 y 3 siguen contiguamente las filas de los carpales distales 2 y 3. En la aleta derecha, se encuentran tres elementos de tamaño superior al del resto de los adyacentes (FIGURA 3, señalados como m?) y de forma poligonal también diferente que se consideran posibles metacarpales o carpales distales.
La aleta izquierda presenta al menos ocho dígitos: cuatro primarios, dos preaxiales, y dos postaxiales (FIGURA 2). Los dígitos primarios corresponden a los dígitos II, III, IV y V, de acuerdo con su posición topológica (Motani, 1999a). Tanto el IV como el V perdieron su secuencia proximal que articula con los matacarpales. El dígito más largo es el IV con 20 falanges conservadas en secuencia continua, seguido del III con 18 falanges en secuencia continua. El dígito II conserva 14 falanges en secuencia continua y del V sólo se pueden contar 13 en su parte distal. El primer elemento del dígito accesorio anterior es más o menos pentagonal con 4 facetas articulares (FIGURAS 2, 4C). Es un elemento grande en comparación con los carpales, metacarpales y las falanges y presenta una hendidura posterodistal muy pronunciada. A partir de este elemento se originan dos dígitos preaxiales. Posteriormente al dígito V se ubican al menos dos dígitos postaxiales (FIGURA 2).
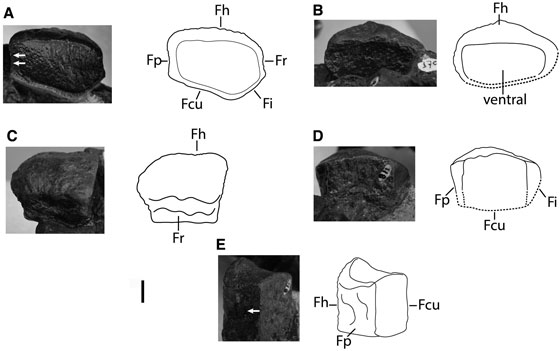
FIGURA 6 Espécimen FCG-CBP-87. Fotografías e interpretaciones del cúbito derechoA. Vista dorsal. B. Vista ventral. C. Vista anteroproximal. D. Vista posterodistal. E. Vista posterodistal. Las flechas indican hendiduras posterior y anterior. Las líneas de puntos representan la interpretación de los sectores cubiertos por sedimento u otro hueso. Barra de escala: 1 cm.
Todas las falanges se encuentran estrechamente empaquetadas (FIGURA 2). La mayoría de las falanges proximales son más altas dorsoventralmente que anchas proximodistalmente mientras que las más distales presentan una forma general hexagonal aplanada. Las falanges decrecen en tamaño distalmente de manera regular. Las falanges de los dígitos primarios son más cuadrangulares que los de los dígitos accesorios. Las proximales de los dígitos accesorios son poligonales con suaves bordes redondeados, y las falanges distales varían de forma poligonal a ovalada, conservando una organización fuertemente empaquetada. Tanto los dígitos preaxiales como los postaxiales presentan superficies dorsales más o menos planas. Los elementos de los dígitos preaxial 1 y preaxial 2 son tan largos como anchos, mientras que los de los dígitos postaxial 1 y postaxial 2 son más largos que anchos. El número de falanges preservadas en los dígitos accesorios es: Pr2=8, Pr1=12, Pt1>9, Pt2>4.
Costillas- Se conservan once fragmentos de costillas (FIGURAS 2, 3, 4A). Contiguos al húmero de la aleta izquierda hay tres fragmentos que son los más largos y corresponden a extremos proximales. Estos fragmentos son ligeramente curvados, tienen cabeza articular bifurcada y son comprimidos anteroposteriormente (FIGURA 4E). A lo largo de su eje estos fragmentos presentan una depresión de tal forma que en corte transversal, estas costillas tienen forma de 8 con la región dorsal más robusta que la ventral.
DISCUSIÓN
Los rasgos diagnósticos establecidos por Fischer et al. (2012) para definir las subfamilias de Ophthalmosauridae incluyen algunas características de los miembros anteriores. En la diagnosis para Ophthalmosaurinae Baur, 1887, se incluye la presencia de una faceta cubital desviada posterodistalmente y un cúbito grande con superficie posterior cóncava. Aunque en FCG-CBP-87 el cúbito tiene superficie posterior cóncava, este hueso es sólo ligeramente más grande que el radio y la faceta cubital no se encuentra desviada posterodistalmente. Estas características separan el espécimen en estudio de la subfamilia Ophthalmosaurinae. Para Platypterygiinae Arkhangelsky, 2001, el rasgo diagnóstico corresponde a la presencia de trocánteres dorsal y ventral prominentes sobre el húmero. Puesto que el extremo proximal del húmero no se preserva, no se cuenta con evidencia diagnóstica para asignarlo a la subfamilia Platypterygiinae.
Al comparar la morfología de FCG-CBP-87 con los caracteres utilizados por Fischer et al. (2016) en su análisis filogenético de los ictiosaurios, se encuentra que 16 de los 21 caracteres relacionados con las aletas anteriores pueden ser establecidos confiablemente en FCG-CBP-87 en los siguientes estados: 57(0), 58(0), 60(2), 61(0), 62(0), 64(1), 65(0), 67(1), 68(1), 69(2), 70(2), 71(0), 72(1), 73(1), 74(1), y 76(1). La comparación de estos caracteres muestra que los rasgos de FCG-CBP-87 corresponden a una morfología apendicular derivada dentro de los oftalmosáuridos y que FCG-CBP-87 comparte caracteres derivados en mayor medida con Platypterygius y Caypullisaurus.
La preservación de huesos diagnósticos en FCG-CBP-87, permite hacer una comparación detallada de su morfología con los caracteres sinapomórficos propuestos para los distintos géneros de la familia Ophthalmosauridae. Todos los caracteres considerados diagnósticos para Platypterygius en las definiciones más recientes de este género (Kolb y Sander, 2009; Adams y Fiorillo, 2010; Ji et al., 2016) están presentes en FCG-CBP-87: húmero con tres facetas distales para radio, cúbito y un elemento extrazeugopodial anterior; faceta radial más pequeña que la cubital y oblicua; presencia de pisiforme; cúbito más largo anteroposteriormente que el radio; intermedio sin contacto con el húmero; más de siete dígitos sin bifurcaciones, incluyendo más de un dígito accesorio preaxial; falanges rectangulares y fuertemente empaquetadas. De acuerdo con la diagnosis presentada por Ji et al. (2016) para Caypullisaurus, FCG-CBP-87 comparte varias de las sinapomorfías de este género, pero difiere en que el húmero no tiene la faceta para el cúbito más pequeña que la del radio ni en dirección oblicua. Con los demás géneros de Ophthalmosauridae, FCG-CBP-87 difiere en mayor medida de sus caracteres sinapomórficos. La inclusión de FCG-CBP-87 en el género Platypterygius resulta bien soportada.
Las especies descritas para el género Platypterygius incluyen: P. campylodonCarter, 1846 (asignado recientemente a Pervushovisaurus campylodonFischer, 2016); P. americanusNace, 1939; P. australisMcCoy, 1867; P. sachicarumPáramo, 1997; P. hercynicusKuhn, 1946; P. hauthalivon Huene, 1927; P. platydactylusBroili, 1907 (especie tipo del género). Hasta el momento, no se conocen restos apendiculares de P. sachicarum.
FCG-CBP-87 comparte con estas especies algunos caracteres, pero también se diferencia de ellas en algunos rasgos. El húmero de FCG-CBP-87 tiene tres facetas distales (diferente de P. platydactylus, P. hercynicus y algunos especímenes de P. australis) (von Huene, 1922; Maisch y Matzke, 2000; Kolb y Sander, 2009; Zammit et al., 2010). El intermedio de FCG-CBP-87 es pentagonal irregular sin contacto con el húmero (diferente de P. hauthali) (Fernández y Aguirre-Urreta, 2005); el pisiforme no tiene forma creciente (diferente de P. campylodon, P. platydactylus, P. americanus) (McGowan, 1972), sino subtriangular, siendo un hueso masivo en relación con los demás huesos del zeugopodio; FCG-CBP-87 tiene más de 1 dígito postaxial (diferente de P. americanus y P. hauthali) (Fernández y Aguirre-Urreta, 2005); el dígito más largo de FCG-CBP-87 tiene 20 elementos (diferente de P. australis y probablemente P. hercynicus) (Kolb y Sander, 2009; Zammit et al., 2010); la aleta de FCG-CBP-87 tiene 8 dígitos (P. hercynicus tiene 8 con dígitos adicionales interaxiales) (Kolb y Sander, 2009); el radio es ligeramente más pequeño que el cúbito (diferente de P. australis) (Wade, 1990).
FCG-CBP-87 presenta rasgos que no se conocen en ninguna de las especies de Platypterygius y que podrían ser considerados de utilidad taxonómica: (1) la faceta para el elemento extrazeugopodial anterior del húmero está dirigida anteriormente, tiene un reborde pronunciado y la superficie de articulación es cóncava significativamente; (2) el cúbito presenta hendiduras pronunciadas en sus superficies anterior y posterior; (3) el extrazeugopodial tiene forma de ‘silla de montar’ con 4 facetas articulares, proximal, distal, posteroproximal y posterodistal; y (4) el pisiforme es masivo y de forma distintivamente subtriangular. Estas diferencias, señalan a FCG-CBP-87 como una especie distinta a todas aquellas de las que se conocen restos de apéndices anteriores.
Teniendo en cuenta que P. Sachicarum procede de la misma región geográfica y de la misma unidad estratigrágfica (miembro “Arcillolitas Abigarradas” de la Formación Paja) de la que procede FCG-CBP-87 y que el tamaño de los dos puede ser correlacionado, existe la posibilidad de que FCG-CBP-87 corresponda a restos apendiculares de P. Sachicarum. Sin embargo, como se desconocen los restos apendiculares P. sachicarum, no es posible establecer ningún tipo de comparación entre FCG-CBP-87 y esta especie. Sólo el hallazgo de material craneano y aletas asociadas con rasgos diagnósticos permitirá establecer o descartar alguna relación entre P. sachicarum y FCG-CBP-87. Sin evidencia diagnóstica se considera aquí adecuado limitar la ubicación sistemática de FCG-CBP-87 a Platypterygius sp.
CONCLUSIONES
El esqueleto apendicular de FCG-CBP-87 es incluído en el género Platypterygius por exhibir una morfologia consistente con las definiciones recientemente publicadas para este género. Sin embargo FCG-CBP-87 se diferencia de las especies incluidas dentro de Platypterygius en varias características relacionadas con el zeugopodio. FCG-CBP-87 se distingue por presentar la faceta para el elemento extrazeugopodial anterior del húmero dirigida anteriormente, con reborde pronunciado y superficie de articulación cóncava; el elemento extrazeugopodial anterior en forma de silla de montar con 4 facetas articulares; el cúbito con hendiduras pronunciadas en sus superficies anterior y posterior y el pisiforme masivo y en forma distintivamente subtriangular. Su relación con P. sachicarum no puede ser establecida por falta de material de comparación. El nuevo espécimen constituye la segunda evidencia de la presencia de Platypterygius en el mar interior del Cretácico temprano de Colombia y el primer reporte de restos apendiculares de Platypterygius procedentes del extremo norte de Suramérica. Además, ofrece nueva evidencia de la excelente preservación alcanzada en los restos de vertebrados del yacimiento paleontológico de la región de Villa de Leiva.

