










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488On-line version ISSN 2665-4385
Rev. Colomb. Entomol. vol.29 no.1 Bogotá Jan./June 2003
Caracterización de genotipos de fríjol ( Phaseolus vulgaris) por su resistencia a Thrips palmi (Thysanoptera: Thripidae)
Characterization of dry bean genotypes ( Phaseolus vulgaris)for resistance to Thrips palmi (Thysanoptera: Thripidae)
JHON DIAZ M.1, CESAR CARDONA M.2, JUAN MIGUEL BUENO1
1 Centro Internacional de Agricultura Tropical, CIAT. Cali.
2 Autor para correspondencia: Centro Internacional de Agricultura Tropical, CIAT. A. A. 6713, Cali. E-mail: c.cardona@cgiar.org
Resumen. Con el fin de identificar fuentes de resistencia a Thrips palmi Karny en fríjol ( Phaseolus vulgaris L.) se hicieron tres experimentos en condiciones de campo e infestación natural en el municipio de Pradera (Valle del Cauca, Colombia). En el primero, se caracterizaron por resistencia al insecto 38 genotipos en un diseño de bloques al azar. En los otros dos experimentos, se utilizaron nueve genotipos en diseños de parcelas divididas; en un caso la parcela mayor fue niveles de fertilización, y en el otro niveles de protección química. Se detectó variabilidad al ataque de T. palmi en los genotipos evaluados. La distribución del insecto en el campo fue agregada. Las poblaciones estuvieron directamente relacionadas con el daño causado a la planta y, éste incidió negativamente en la adaptación reproductiva y en los rendimientos. Los genotipos Brunca, BH 21134-5, EMP 486, BAT 477, FEB 115 y BH 21134-60 mostraron resistencia al insecto a través de los experimentos. Sin embargo, es necesario hacer otros estudios para conocer cuál o cuáles mecanismos de resistencia (antixenosis, antibiosis o tolerancia) están presentes en ellos. No hubo efecto de las condiciones nutricionales en la expresión de resistencia. Las pérdidas causadas por T. palmi oscilaron entre 11,9 y 65,1%. FEB 115, BH 21134-5, EMP 486, BH 21134-60 y Brunca estuvieron por debajo del índice de susceptibilidad promedio del experimento; genotipos con índices bajos tienen mayor tolerancia al daño y menores pérdidas. Es necesario realizar más trabajos de investigación con el fin de descifrar los mecanismos de resistencia a los trips en genotipos resistentes.
Palabras clave: Adultos. Larvas. Infestación natural. Daño. Fertilización. Pérdidas. Trips.
Summary. To identify sources of resistance to Thrips palmi Karny in dry beans ( Phaseolus vulgaris L.), three field trials were conducted in Pradera (Cauca Valley, Colombia) under heavy natural infestation. In the first trial, 38 genotypes were characterized for resistance to T. palmi in a completely randomized block design. In the other two trials, nine genotypes were used in a split-plot design. In one case, the main plot was fertilization level; while in the other, chemical protection was the main plot. Variation in T. palmi attack was detected in the genotypes tested. Thrip populations showed an aggregate pattern of distribution and had a significant effect on damage levels to plants. Damage negatively affected reproductive adaptation and yield plants. Genotypes Brunca, BH 21134-5, EMP 486, BAT 477, FEB 115 y BH 21134-60 showed resistance to Thrips in the experiments. Fertilization levels did not have an effect on resistance expression. Losses due to Thrips damage ranged from 11,9% to 65,1%. FEB 115, BH 21134-5, EMP 486, BH 21134-60 and Brunca were below the average susceptibility index of the experiment; genotypes with low susceptibility indexes may have tolerante to damage and suffer lower losses. Further work is needed in order to elucidate the mechanisms of resistance to Thrips in resistant genotypes.
Key words: Adults. Larvae. Natural infestation. Damage. Fertilization. Losses. Thrips.
Introducción
El trips del melón, Thrips palmi Karny, es una plaga relativamente nueva en Colombia; este insecto tiene una amplia gama de hospederos,es muy agresivo y presenta alta capacidad de reproducción. Su control es muy difícil y requiere aplicaciones repetitivas de insecticidas. Por 10 tanto, entidades privadas y del Estado desde su aparición han comenzado a investigar las características generales de T. palmi y posibles métodos de manejo.
T. palmi tiene una amplia distribución geográfica. Inicialmente fue encontrado en tabaco en las islas de Sumatra Y Java en Indonesia (Karny 1925 citado Por Cermeli Y Montagne 1993). A finales de la década de los años 70 comenzó a ser una plaga seria en regiones tropicales y subtropicales como el sudeste de Asia, las Islas del Pacífico y del Caribe (Kawai 1990b). Fue registrado por primera vez en la Florida (Estados Unidos) en 1990 (FA0 1991). Posteriormente apareció en Venezuela (Cermeli y Montagne 1990) y en Cuba (Jiménez 1998). En colombia se señaló su aparición en marzo de 1997 ( Mesa y Zuluaga1997) .
T. palmi es un insecto polífago, que en altas poblaciones causa daños severos y pérdidas significativas. En el departamento del Valle del Cauca, Durán et al. (1999) señalaron como hospederos de T. palmi a más de 25 especies vegetales. Los huevos de T. palmi son depositados individualmente en los tejidos de las plantas; el insecto tiene dos estados larvales que se alimentan de las plantas; cuando el segundo ínstar madura, cae al suelo y allí muda a prepupa y pupa. Después de la emergencia los adultos se trasladan a las partes en crecimiento de las plantas tales como hojas jóvenes, flores y frutos jóvenes, donde se alimentan y depositan los huevos. T. palmi puede habitar diferentes partes de los cultivos hospederos. Los adultos se encuentran casi siempre sobre tejidos jóvenes, mientras las larvas están sobre tejidos un poco más viejos, lo cual da lugar a diferencias en la distribución del insecto en la planta así como a la aparición de distintos tipos de daño en las plantas hospedantes (Kawai 1990b). Los hábitos que tiene T. palmi de depositar los huevos en los tejidos de las plantas y de pasar los estados de prepupa y pupa en el suelo, hacen más difícil el control con productos químicos a no ser que se apliquen muchas veces; son muy pocos los productos químicos efectivos para el control de T. palmi (Kawai 1990a).
Una alternativa de control podría ser la resistencia varietal. En estudios preliminares realizados por el equipo de Entomología de Fríjol del Centro Internacional de Agricultura Tropical (CIAT-palmira), se detectaron niveles de variabilidad en la respuesta al insecto que sugirieron la posible existencia de fuentes de resistencia a T. palmi.
Por las razones anteriormente expuestas se realizó este trabajo encaminado a probar la hipótesis de que no existe resistencia varietal T. palmi en fríjol. En consecuencia, el estudio se orientó a conocer las características de infestación de la plaga y su efecto en el cultivo, caracterizar genotipos por su resistencia o susceptibilidad a T. palmi, observar la respuesta al insecto de algunos genotipos de fríjol en parcelas con y sin fertilización y finalmente realizar estudios en ambientes protegidos y no protegidos para conocer los niveles de pérdidas en rendimiento ocasionados por T. palmi en diferentes genotipos.
Materiales y Métodos
El presente estudio se adelantó en condiciones de campo e infestación natural en el municipio de Pradera (Valle del Cauca), vereda La Floresta, finca La Esquina, a una altura de 980 msnm y temperatura promedio de 23°C. Se hicieron tres ensayos, uno en cada uno de los semestres 19998,2000A y 2000B.
En el primer ensayo (Tamizado de genotipos de fríjol por resistencia a T. palmi) se caracterizaron por resistencia o susceptibilidad 38 genotipos (seleccionados en estudios previos por haber mostrado niveles diferenciales de resistencia o susceptibilidad). Cada genotipo se sembró en una parcela de 2 m y tuvo cuatro repeticiones dispuestas en un diseño de bloques al azar. Las distancias de siembra fueron de 0,5 m entre surcos y 0 , l m entre plantas. No se aplicaron fertilizantes y las infestaciones de mosca blanca se controlaron con buprofezin (0.065 kg i.a.1 ha). Este insecticida es selectivo para controlar estados inmaduros de mosca blanca y no afecta a ningún estado de T. palmi.
En el segundo ensayo (Efecto de la fertilidad del suelo en la expresión de resistencia a T. palmi) se estudió el posible efecto del estado nutricional de la planta en la expresión de resistencia. Se seleccionaron cuatro genotipos que mostraron resistencia en el primer experimento (Brunca, EMP 486, BAT 477 y FEB 115) y dos (BH 21134- 5 y BH 21134-60) seleccionados en otros estudios adelantados por el CIAT. Como testigos susceptibles se incluyeron EMP 514, APN 18 y BAT 41. Estos genotipos se dispusieron en un diseño de parcelas divididas con cuatro repeticiones en el cual dos niveles de fertilidad (fertilización óptima determinada mediante previo análisis del suelo y no-fertilización) conformaron las parcelas mayores y los nueve genotipos constituyeron las parcelas menores o subparcelas. Cada subparcela estuvo constituida por cuatro surcos de 3 m de longitud con distancias de 0,5 m entre surcos y 0,l m entre plantas. Los fertilizantes recomendados en el análisis del suelo fueron cloruro de potasio (KCI), urea, óxido de magnesio (MgO) y bórax a dosis de 100, 65, 60 y 6.7 kg/ha, respectivamente.
En el Último ensayo (Medición de pérdidas causadas por T. al mi en aenotipos de fríjol) se calculó la respuesta en rendimiento de los genotipos a dos niveles de infestación: no infestación (protección química completa) y alta infestación (sin protección). Para esto se usaron los mismos genotipos utilizados en el ensayo anterior, dispuestos en un diseño de parcelas divididas con cuatro repeticiones, en el cual la protección química y la no protección constituyeron las parcelas mayores y los diferentes genotipos constituyeron las parcelas menores o subparcelas. Las distancias de siembra fueron las mismas del ensayo anterior. Los insecticidas usados en las parcelas protegidas para mantener los niveles de población tan bajos como fuera posible fueron spinosad (únicamente en la primera aplicación) e imidacloprid a dosis de 0.023 kg i.a./ha y 0.21 kg i.a./ha, respectivamente.
Se hicieron recuentos semanales de población. Los adultos de T. palmi se contaron en el foliolo central de una hoja de la parte superior de la planta y las larvas se contaron en el foliolo central de una hoja de la parte media de la planta. El tamaño de muestra fue de tres foliolos al azar por parcela para adultos y tres foliolos al azar por parcela para larvas en el primer experimento y de cuatro foliolos al azar en cada subparcela tanto para adultos como para larvas en los otros experimentos. Estos datos de población fueron usados para calcular áreas bajo la curva resultantes del acumulado de población a través del período vegetativo. En el primer experimento se graficaron sólo los cinco genotipos más contrastantes por su respuesta al ataque del insecto.
El daño causado por T. palmi a los genotipos en estudio se estimó mediante una escala visual de daño, de 1 a 9, donde 1 era sin daño visible y 9 era daño generalizado de la planta por defoliación y quemazón (CIAT 1999). En la madurez del cultivo se estimó la adaptación reproductiva (habilidad de la planta para formar vainas bajo la presión de insectos) mediante una escala visual, de 1 a 9, donde 1 era pésima, la planta ha perdido el 80% o más de su potencial de rendimiento y 9 era excelente, la planta muestra todo su potencial de rendimiento, altura de la planta, forma, número y tamaño de las vainas aparentemente normales. Por último se tomaron los rendimientos en glplanta en el primer experimento y en kgl ha para los otros experimentos.
En el tercer experimento se calculó el porcentaje de pérdida causado por T. palmi en cada genotipo:
% Pérdida =
ks/ha (protegido) - ks/ha (no protegido) /kg/ha (protegido) x 100
también se calculó el índice de susceptibilidad (pérdida en cada genotipo con respecto a la pérdida promedio calculada para el experimento) mediante la siguiente fórmula:
I.S.= 1 - (npxi/pxi) / 1 - (npx/px) . . donde:
npxi es el rendimiento promedio en kg/ha de un genotipo en parcelas no protegidas.
px,, es el rendimiento promedio en kg/ha de un genotipo en parcelas protegidas. npx, es el rendimiento promedio en kglha de todos los genotipos en parcelas no protegidas.
px, es el rendimiento promedio en kg/ha de todos los genotipos en parcelas protegidas. 'ibdos los datos de poblaciones de adultos y larvas fueron sometidos a análisis de varianza (ANDEVA) previa transformación a log (x) o log (x + 1) (primer experimento) y a Ecu (segundo y tercer experimentos). Cuando la prueba de F fue significativa, se procedió a hacer la separación de medias por diferencia mínima significativa (DMS) al 5% de confiabilidad (Statistix 1998). Los análisis de varianza de los experimentos en parcelas divididas y la separación de medias entre tratamientos y genotipos al 5% de confiabilidad se hicieron mediante el programa Mstat-C (1991). Los cálculos de áreas bajo la curva para adultos y larvas y los correspondientes análisis de varianza y separaciones de medias se hicieron mediante el programa SAS (1989) previa transformación de los datos de población a log (x+l).
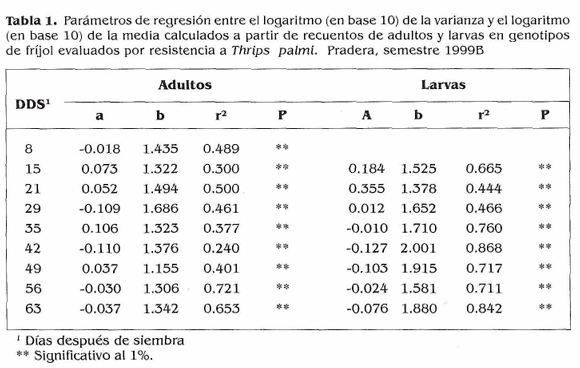
Para estudiar la agregación o distribución del insecto en el campo se usó el método de la ley de poder de Taylor basada en la relación entre el logaritmo de la varianza y el logaritmo de la media, bien sea de adultos o de larvas encontradas (Taylor 1961; Taylor et al. 1978; Taylor y Woiwod 1980 citados por Lewis 1997). La ecuación de regresión es log (SZ) = log (a) + (b) log (m), donde a y b son el intercepto y la pendiente, respectivamente. La pendiente (b) es una medida de agregación, y el intercepto log (a) es un factor de escala relacionado con el tamaño de la muestra. Una pendiente de 1.0 indica dispersión al azar, una pendiente < 1.0 sugiere dispersión uniforme, y una pendiente > 1.0 indica agregación (Taiylor 1961; Harcourt 1965; Southwood 1978 citados por Pearsall y Myers 2000).
Todos los datos de calificaciones de daño, adaptación reproductiva y rendimiento fueron sometidos a análisis de varianza y posterior separación de medias por DMS al 5% utilizando los programas Statistix (1998) y Mstat-C (1991).
Resultados y Discusión
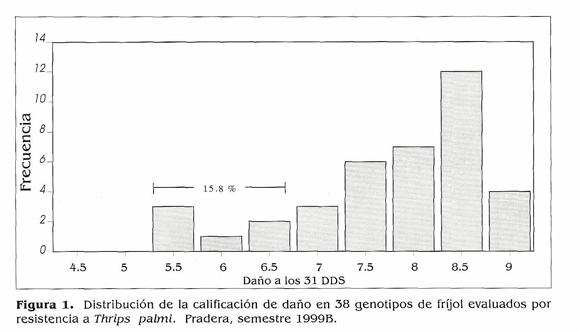
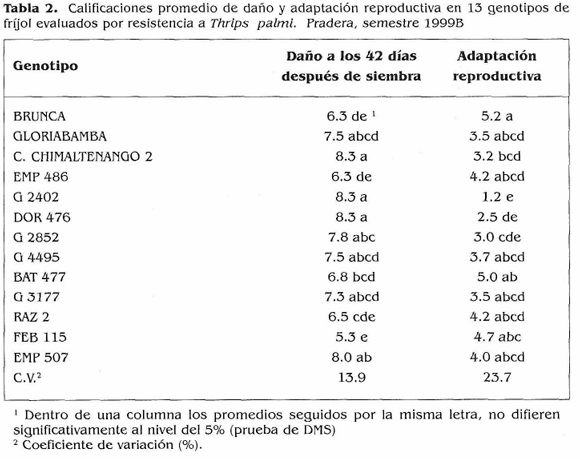
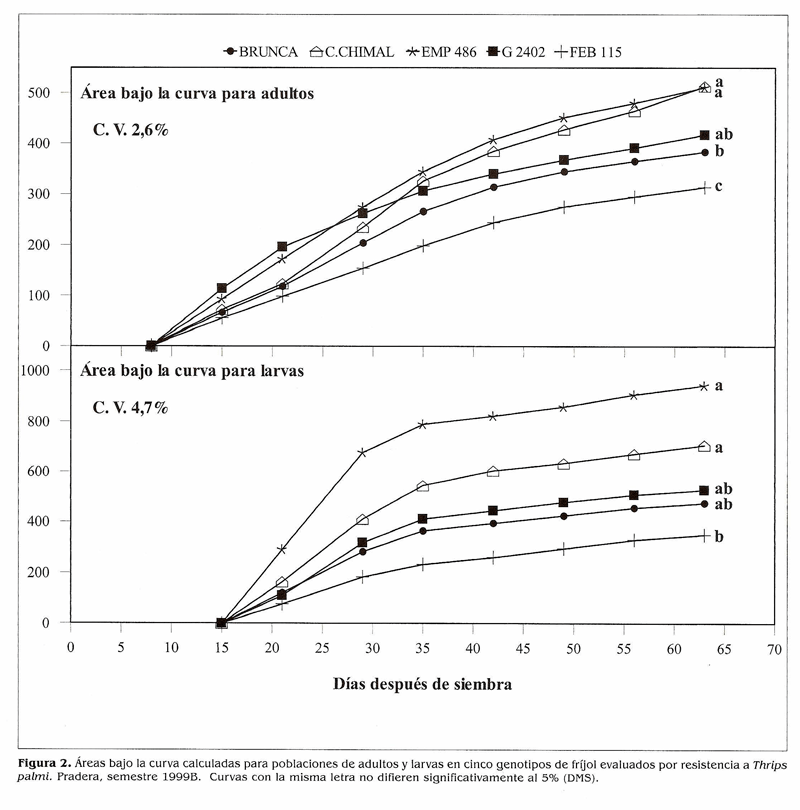
Tamizado de genotipos de frijol por su resistencia a T. palmi. Al inicio del experimento T. palmi ocurrió en poblaciones muy altas que causaron daños tempranos y fuertes en la mayoría (84,2%) de los genotipos evaluados a los 31 días después de siembra (DDS) (Fig. 1). Por estas razones se continuó el ensayo con 13 genotipos. El estudio de la distribución del insecto mostró que a través del periodo vegetativo del cultivo todos los valores de pendiente (b) fueron significativamente mayores de 1.0 (P < 0.001) indicando un patrón de distribución agregado para adultos y larvas de T. palmi en cultivos de fríjol sembrados en el campo (Tabla 1). El área bajo la curva de la población en cinco genotipos contrastantes (Fig. 2) mostró a los testigos susceptibles Compuesto Chimaltenango 2 y G 2402 con altos e intermedios acumulados de población, por lo que sus calificaciones finales de daño fueron las más altas (Tabla 2). EMP 486 presentó el mayor acumulado de población, mientras que Brunca y FEB 115 tuvieron los menores; estos tres materiales fueron menos afectados por el insecto, Podría ser que mecanismos de antibiosis, antixenosis actúen como defensas de la planta, pero datos obtenidos en el campo generalmente no permiten identificar estos mecanismos con absoluta precisión, por lo que habría necesidad de adelantar estudios en invernadero para dilucidar si estos mecanismos están presentes en estos genotipos. Brunca, EMP 486, BAT 477, G 3177, RAZ 2 y FEB 115 mostraron cierta resistencia a T. palmi la cual se reflejó en calificaciones de daño menores y en adaptaciones reproductivas regulares o intermedias (Tabla 2).
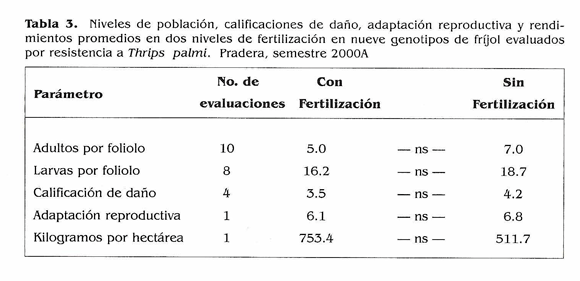
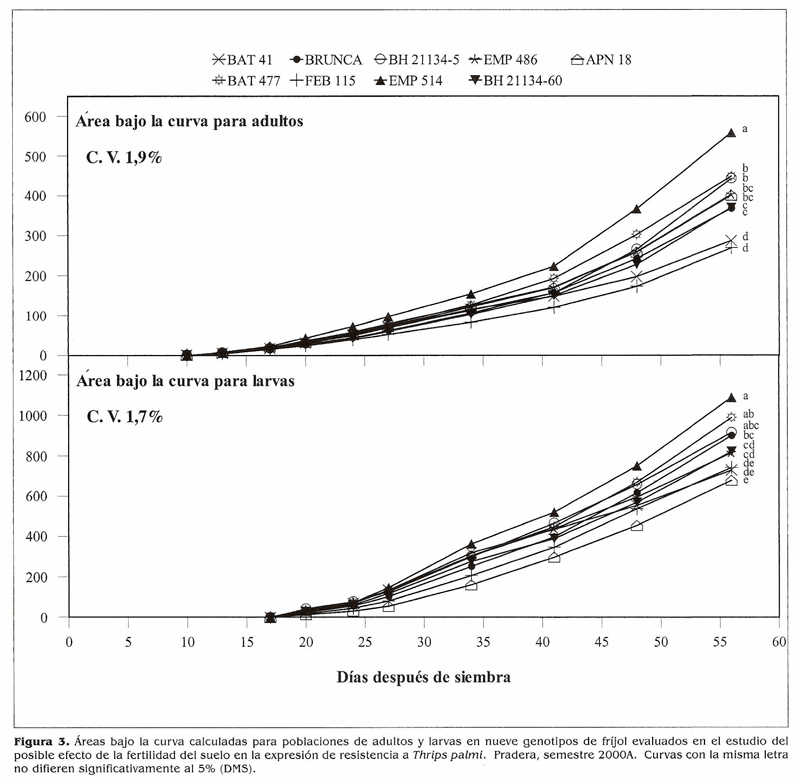
Efecto de la fertilidad del suelo en la expresión de resistencia a T. palmi. En este experimento no se encontraron diferencias significativas entre los niveles de fertilidad para ninguno de los recuentos y/ o evaluaciones (Tabla 3). El cálculo del área bajo la curva de la población de T. palmi (Fig. 3) mostró al genotipo susceptible EMP 514 con los acumulados de población más altos y con uno de los daños mayores (Tabla 4). APN 18, también escogido por ser susceptible mostró el daño mayor, lo cual sugiere que este genotipo es altamente susceptible a T. palmi. El caso del otro tes- I tigo susceptible, BAT 41, fue diferente. Como es tan precoz, es posible que se haya f presentado evasión o escape al ataque del l insecto por lo que sufrió el menor daño. PEB 115, nuevamente mostró niveles significativamente inferiores de población y podría poseer antibiosis o antiienosis como mecanismos que le ayudan a mantener menores poblaciones de T. palmi que los demás. Sin embargo, como se mencionó anteriormente, se deben adelantar estudios en invernadero para determinar los mecanismos que confieren resistencia a estos genotipos. Debido a la baja población al inicio del ensayo los daños finales no fueron tan altos y la adaptación reproductiva fue regular para los genotipos; a excepción de APN 18 y EMP 514 que presentaron los daños mayores y al lado de BAT 41 tuvieron adaptaciones reproductivas malas (Tabla 4).
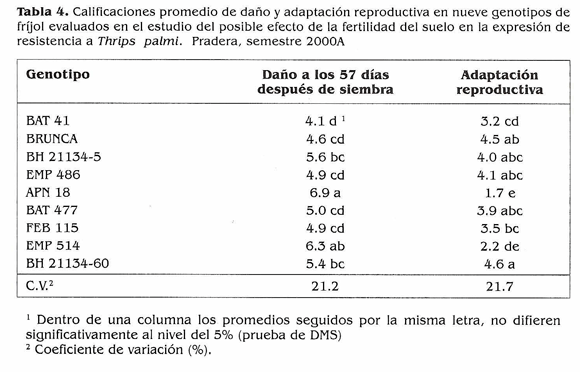
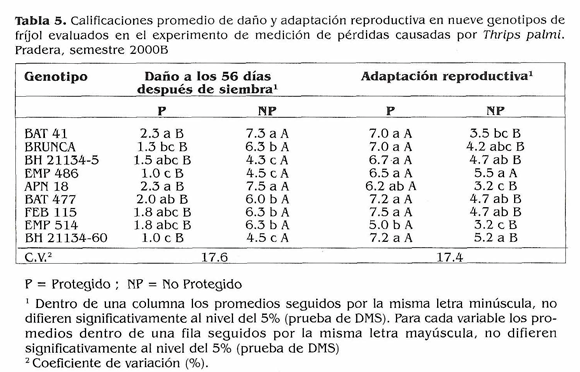
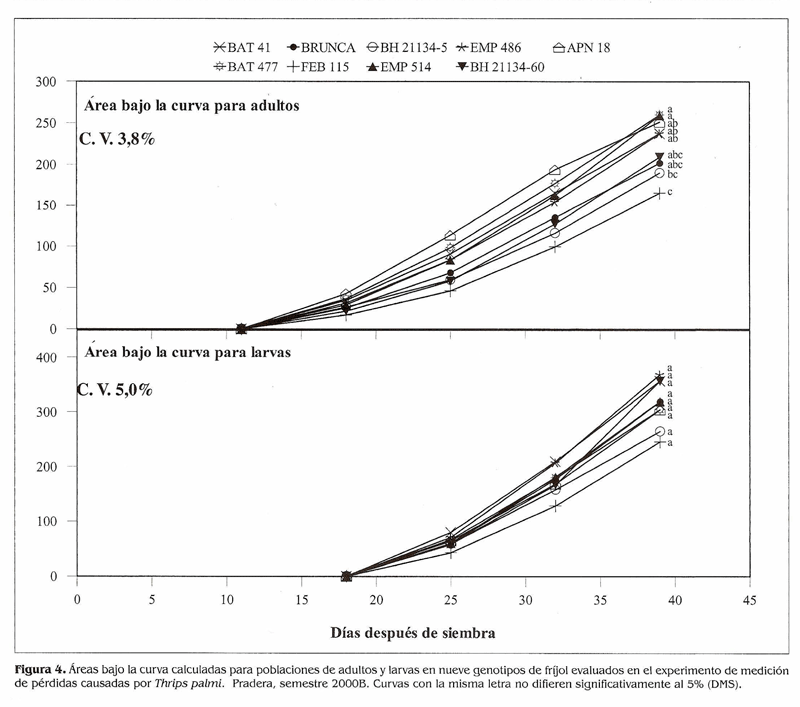
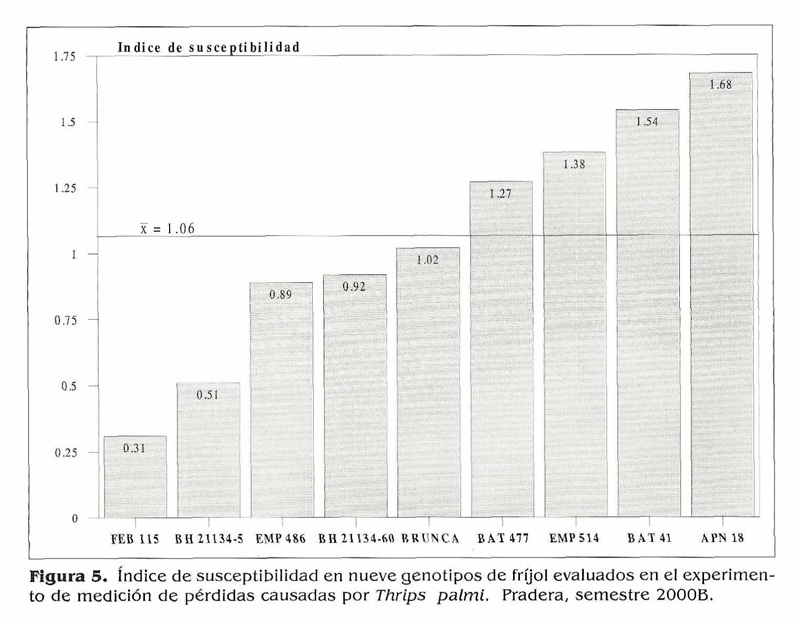
Medición de pérdidas causadas por T. palmi en genotipos de fríjol. Uno de los principales objetivos de este trabajo fue estudiar el comportamiento de las poblaciones de T. palrni en condiciones de infestación natural, razón por la cual se utilizaron los promedios de las parcelas no protegidas en las variables relacionadas a la población. Se presentaron incrementos de población desde los primeros días, por lo que hubo daños tempranos en los genotipos susceptibles y sólo se hicieron conteos del insecto hasta los 39 DDS. El área bajo la curva para las poblaciones (Fig. 4) mostró pocas diferencias en adultos, y para larvas no hubo diferencias estadísticas: los acumulados mayores los tuvieron los testigos susceptibles BAT 4ly APN 18 que presentaron las calificaciones finales de daño más altas (Tabla 5): los acumulados de población menores lo presentaron BH 21 134-5 y FEB 115; Brunca y BH 21154-60 presentaron acumulados intermedios. Esto sugiere que posiblemente estén actuando antixenosis o antibiosis como posibles mecanismos de resistencia, pero la caracterización de estos mecanismos de esto requeriría trabajos de invernadero en condiciones controladas. Los testigos susceptibles BAT 41, APN 18 y EMP 514 tuvieron malas adaptaciones reproductivas en las parcelas no protegidas; los demás genotipos finalizaron con calificaciones de daño menores de 6.5 y tuvieron adaptaciones reproductivas regulares (Tabla 5). Los testigos susceptibles BAT 41, APN 18 y EMP 514 presentaron l a s mayores pérdidas en rendimiento con porcentajes superiores al 50% (Tabla 6); Brunca, EMP 486, BAT 477 y BH 21134-60 tuvieron pérdidas entre 35 y 49%, a pesar de haber mostrado resistencia a T. palmi. Los porcentajes de pérdidas ocasionados por el ataque del insecto a los genotipos mencionados son bastante altos, lo que demuestra que si no se hace ningún control en contra de esta plaga cuando se presenta en poblaciones altas, ésta puede ocasionar reducciones considerables en los rendimientos. BH 21134-5 y FEB 115 tuvieronlas pérdidas menores y no mostraron diferencias significativas en sus rendimientos en los niveles de infestación (protección y no protección), lo cual sugiere que podrían llegar a desarrollarse sin ningún control o con controles mínimos bajo infestaciones fuertes de T. palmi. En estudios posteriores se tratará de dilucidar si los mecanismos de antibiosis y antixenosis están presentes en estos genotipos. El índice de susceptibilidad promedio fue 1,06 el cual se obtuvo al promediar el índice de susceptibilidad de los nueve genotipos. Además de mostrar resistencia a T. palrni, Brunca, BH 21134-5, EMP 486, FEB 115 y BH 21134-60 presentaron menor reducción en sus rendimientos y estuvieron por debajo del índice de susceptibilidad general (Fig. 5), es decir, que podrían llegar a desarrollar todo su potencial de rendimiento con controles mínimos (pocas aplicaciones de insecticidas) en contra de T. palmi sin ocasionar pérdidas significativas. Mientras que BAT 477 y los tres testigos susceptibles requieren un mayor control (más aplicaciones calendario) de la plaga para no causar pérdidas tan altas como las que mostraron en este experimento. Las fuentes de resistencia aquí identificadas tienen la ventaja de pertenecer al acervo genético primario en frijol, por lo cual podrían ser utilizadas directamente en un programa de mejoramiento genético por resistencia a la plaga.

Conclusiones
- Se determinó que la distribución de T. palmi en cultivos de fríjol en el campo es agregada, lo que sumado a su agresividad y dificultades de control hacen más difícil el manejo de la plaga.
- A través de los experimentos, los genotipos Brunca, BH 21134-5, EMP 486, BAT 477, FEB 115 y BH 21134-60 mostraron niveles de resistencia a T. palmi que se reflejaron en bajas calificaciones de daño, adaptaciones reproductivas regulares o intermedias y en mejores rendimientos.
- En las condiciones experimentales del presente trabajo no hubo ningún efecto de las condiciones nutricionales de la planta en la expresión de resistencia a T. palmi.
- En general las poblaciones de T. palmi tuvieron una alta incidencia en el daAo ocasionado a las plantas y éste a su vez incidió negativamente en la adaptación reproductiva, y en los rendimientos de los genotipos.
- En este trabajo se evaluaron 41 genotipos y 33 (80,5%) de ellos presentaron alta suceptibilidad a T. palmi. - Las pérdidas estimadas en los rendimientos para los genotipos que mostraron resistencia al insecto variaron entre 11,9 y 49,4% y para los testigos susceptibles entre53.3 y 65.1%. FEB 115, BH 21134-5, EMP 486, BH 2 1134-60 y Brunca estuvieron por debajo del índice de susceptibilidad promedio del experimento, es decir que sus pérdidas fueron menores que la pérdida promedio del ensayo.
- Hay variabilidad genética para la respuesta al ataque de T. palmi. El presente trabajo permitió identificar algunos genotipos con resistencia a la plaga, pero no permite establecer con exactitud los mecanismos de resistencia que estén actuando en contra del insecto.
- Las fuentes de resistencia aquí identificadas pueden ser usadas en un programa de mejoramiento genético por resistencia a T. palmi.
Literatura citada
CERMELI, M.: MONTAGNE, A. 1990. Thrips palmi Karny (Thysanoptera: Thripidae) nueva plaga para Venezuela. Boletín de Entomología Venezolana 5 (29): 192. [ Links ]
CERMELI , M .: MONTAGNE , A . 1993 . Situación actual de Thrips palmi Karny (Thysanoptera: Thripidae) en Venezuela. Manejo Integrado de Plagas. Centro Agronómico Tropical de Investigación y Enseñanza - CATIE . Turrialba Bol. No . 29 : 22-23 [ Links ]
CIAT (CENTRO INTERNACIONAL DE AGRICULTURA TROPICAL). 1999. Annual Report. Project IP-l. Bean Improvement for Sustainable Productivity, Input Use Efficiency, and Poverty Alleviation. 139 p. [ Links ]
DURÁN, I .: MESA, N.: ESTRADA, . Revista Colombiana de Entomología 25 (3-4109-120 [ Links ]
FAO. 1991. First record of Thripspalmi in continental United States. Plant Prot. Bull. 39 (4): 188. [ Links ]
JIMÉNEZ, L. 1998. Recomendaciones al grupo de trabajo que trabaja en la problemática de Thrips palmi Karny en el Oriente Antioqueño - Colombia. Informe sobre visita de trabajo. Facultad de Agronomía, Universidad de La Habana, La Habana. 35 p. [ Links ]
KAWAI, A. 1990a. Control of Thripspalmi Karny in Japan. Japan Agricultural Research Quarterly 24 (1): 43-48. [ Links ]
KAWAI, A. 1990b. Life cycle and population dynamics of Thrips palmi Karny. Japan Agricultural Research Quarterly 23 (4): 282-288. [ Links ]
LEWIS, T. 1997. Thrips as Crop Pests. CAB International. Wallingford, UK. 740 p. [ Links ]
MESA, M.; ZULUAGA, J. 1997. Estudios básicos sobre una nueva amenaza para la horti-cultura Colombiana. Thrips palmi (Thysanoptera: Thripidae). Universidad Nacional de Colombia. palmira. 3 p. (manuscrito no publicado). [ Links ]
MSTAT-C. 1991. Microcomputer Statistical Program User's guide. Michigan State University. [ Links ]
PEARSALL, I.: MYERS, J. 2000. Evaluation of sampling methodology for determining the phenology, relative density, and dispersion of western flower Thrips (Thysanoptera: Thripidae) in nectarine orchards. J. Econ. Entomol. 93 (2): 494-502. [ Links ]
SAS Institute. 1989. SASISTAT User's guide version 6. 41h Ed. Vol 2. SAS Institute, Cary, M.C. 846 p. [ Links ]
STATISTIX. 1998. Analytical Software. Statistix for Windows. User's manual. 333 p. [ Links ]
Recibido: May. 15 / 2002 Aceptado: Ago. 08 1 2002


{kind=link}
{kind=link}
{kind=link}
{kind=link}