










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkRevista Colombiana de Entomología
versión impresa ISSN 0120-0488versión On-line ISSN 2665-4385
Rev. Colomb. Entomol. v.29 n.1 Bogotá ene./jun. 2003
Compared cytogenetic of Drosophila starmeri (Díptera: Drosophüidae) of two Colombian isolated arid ecosystems
CARLOS FERNANDO PRADA G.1, MA. MAGDALENA E. DE POLANCO2, MARINA ORDÓÑEZ V.3
1Biólogo. Universidad del Tolima. Ibagué, Tolima, Colombia. E-mail: biofer@lycos.com
2 Autor para correspondencia: Lie. Biología y Química. M. Se; Ph. D. en Genética. Laboratorio de Citogenética. Facultad de Ciencias, Universidad del Tolima. Altos de Santa Helena, Ibagué, Tolima, Colombia. E-mail: mmpol@eudoramail.com
3Lic. Química y Biología. M. Sc; Biología, Instituto de Genética. Universidad de los Andes. Bogotá, Colombia. E-mail: marordon@uniandes.edu.co
Resumen. Se establece la ruta filogenética del cromosoma 2 de Drosophila starmerí (Wasserman, Koepfer & Ward) de dos poblaciones de la costa norte colombiana (Camarones y Santa Marta) y de una tercera ubicada en el Desierto de la Tatacoa (Rosalía), separada de las dos primeras por más de 1.200 kilómetros. La filogenia se resume en el mapa elaborado, siguiendo el método de las inversiones traslapadas y las técnicas de microfotografía cromosómica y digitalización en escáner. De acuerdo con los resultados obtenidos, el arreglo estándar del cromosoma 2 de Drosophila starmeri se deriva del estándar de Drosophila martensis por cuatro inversiones traslapadas. El cromosoma presenta polimorfismo para tres arreglos diferentes. En este trabajo se propone una ruta filogenética nueva para el cromosoma 2. Los resultados podrían ser interpretados como un reflejo de la divergencia genética entre las poblaciones de la costa norte y de la Tatacoa, con una presunta interrupción del flujo genético, que empieza a vislumbrarse a través de los estudios morfológicos y de aislamiento reproductivo realizados, en las mismas poblaciones, por el grupo de investigación de citogenética, filogenia y evolución de poblaciones, de las Universidades del Tolima y Andes.
Palabras clave: Filogenias cromosómicas. Grupo repleta. Cromosomas politénicos. Inversiones traslapadas.
Summary. The phylogenetic route of Drosophila starmeri (Wasserman, Koepfer & Ward), chromosome 2, was established from two populations in the Colombian Morth Coast (Camarones and Santa Marta) and from a third one located in the Tatacoa desert (Rosalía). The latter was separated from the first two populations by more than 1.200 kilometers. The phylogenesis is summarized in the map, that was made using the method of the overlapped inversions and the techniques of chromosomal microphotography and scanner digitization. According to the results, the standard arrangement of the Drosophila starmeri chromosome 2 stems from the Drosophila martensis standard for four overlapped inversions. This chromosome shows polymorphism in three different arrangements. In this paper, we propose a new phylogenetic for chromosome 2. The results could be interpreted as a reflection of the genetic divergence between the populations of the florth Coast and the Tatacoa, with a presumed interruption of the genetic flow. This divergence begins to be observed through the morphological and reproductive isolation studies done on the same populations, by groups researching cytogenetic, phylogeny and populations evolution, from the Universities of Tolima and Andes.
Key Words: Chromosomal phylogenesis. Repleta group. Polytene chromosomes. Overlapped inversions.
Introducción
El grupo repleta es uno de los más grandes del género Drosophila. Cuenta actualmente con 89 especies descritas y posiblemente existan otras formas aún sin describir. En la evolución cromosómica del grupo, se registran 300 inversiones, que han logrado mantenerse (Wasserman 1954,1982; Polanco 1999).
Las especies del grupo repleta son de gran importancia en los estudios de evolución, ya que aportan evidencia científica tangible de especiación en diversas poblaciones naturales. Un ejemplo de especiación en el grupo repleta es señalado por Ruiz y Wasserman (1993), quienes afirman que los tres enjambres del complejo buzzatii son alopátricos y que el enjambre stalkeri podría ser tanto, el más primitivo, como el más avanzado evolutivamente. Este hecho se puede corroborar con estudios citogenéti-cos, de biología molecular, morfológicos, ecológicos, o de aislamiento reproductivo, los cuales son comúnmente utilizados para determinar las relaciones filogenéticas entre las especies del grupo repleta y entre otros miembros del género Drosophila (Wasserman 1982).
El enjambre martensis está compuesto por cuatro especies: D. martensis (Wasserman 6e Wilson), D. uniseta (Wasserman, Koepfer Sí Ward), D. starmeri (Wasserman, Koepfer & Ward) y D. venezolana (Ruiz y Fontdevila).
Inicialmente se ubicó en el complejo mullen, pero ha sido trasladado por Ruiz y Wasserman (1993), al complejo buzzatii, que agrupa las especies de Suramérica, por no poseer la inversión 3a si no la 3r2.
La evolución citológica del enjambre martensis sigue siendo en esencia la misma descrita por Wasserman y Koepfer (1979), la única diferencia radica en que en la evolución de la especie D. martensis hacia D. buzzatii (Fatterson Sí Wheeler), debe ser reversada. El ancestro del enjambre martensis debió haber presentado un alto polimorfismo que ha evolucionado en el presente a D. starmeri, el cual muestra una marcada diferenciación geográfica entre sus poblaciones. Las otras especies del enjambre emergieron de este ancestro fijando sus propias inversiones (Polanco 1998).
Se han realizado diversas investigaciones encaminadas hacia el esclarecimiento de los procesos evolutivos en el enjambre martensls, aprovechando las poblaciones naturales aisladas (costa norte colombiana y Desierto de la Tatacoa). Una de ellas es la de Castañeda (2000), quien realizó un análisis morfométrico para 8 caracteres. En este trabajo se afirma que entre las poblaciones de D. starmerí de Rosalía y las de la costa norte, existe una distancia genética, atribuida al obligado aislamiento geográfico y genético que se da entre ellas, lo que seguramente ha permitido que a lo largo del tiempo se vaya acumulando una gran cantidad de diferencias genéticas.
En el trabajo de Rojas (2001) se analiza el aislamiento reproductivo en especies del enjambre martensis (D. martensis, D, starmerí y D. uniseta) y se concluye de forma general que entre las poblaciones de la misma especie se alcanza un grado de divergencia genética representado en una disminución de los híbridos. Esta disminución es típica de poblaciones alopátricas, y aisladas como en el caso de la costa norte o la del Desierto de la Tatacoa. En contraste con lo anterior, cuando las poblaciones son relativamente aledañas como en el caso de Camarones y Santa Marta, no se observa una disminución en la productividad de los híbridos. Castañeda (2000) propone que aunque las poblaciones de la costa han presentado divergencia para algunos caracteres morfológicos, ésta parece no haber afectado la productividad de los híbridos provenientes de cruces interpoblacionales.
Estudios realizados a nivel citológico en D. starmerí, en Suramérica, han arrojado diferentes datos, como los presentados por Wasserman y Koepfer (1979), que plantean un cromosoma 2 hipotético 2abd2s6Px<5y los arreglos 2abd2s6Px6z6y6a7c7y 2abd2s6Px6z6a7 como posibles polimorfismos. Ruiz y Fontdevila (1981) registran la presencia de dos razas de D. starmerí, a las que se les denominó Occidental y Oriental. Una de las razas posee las inversiones 2z6, 2V6, 2a7, 2t6, 2q9, Xs y Xy y está restringida a la región Occidental que incluiría las localidades de Sta. Rita y Riohacha, en Colombia, y Cata y Lagunillas en Venezuela. La inversión 2W6 se encuentra solamente en la raza Oriental, en la que se hallan las localidades de Zuata y Guaca en Venezuela.
Ruiz y Wasserman (1993) también señalan el arreglo 2abmnz7e2 como el estándar y los arreglos 2e2, 2Py 2z6 como posibles polimorfismos para D. starmerí.
Por otro lado, Polanco (1998) plantea una posible ruta filogenética para el cromosoma 2 estándar de D. martensis (el arreglo 2ABCDEFOabcd), de la población de Riohacha, que difiere de los arreglos propuestos por los autores Wasserman y Koepfer (1979), Ruiz y Fontdevila (1981) y Ruiz y Wasserman (1993).
Materiales y Métodos
Los sitios de muestreo fueron: Camarones en cercanías a la ciudad de Riohacha (La Guajira), Santa Marta (Magdalena), y Rosalía en el Desierto de la Tatacoa (Huila) (Fig. 1). Dichos sitios se seleccionaron con base en trabajos anteriores, en los que se ha detectado la presencia del enjambre martensis.
Las colectas se realizaron entre las 5:30-7:30 y 17:30-18:30 h. Una hora antes se colocaron las trampas para cebar las moscas, las cuales estaban compuestas de diversas frutas fermentadas. Posteriormente, se capturaron las moscas con una jama de nylon, se clasificaron y cultivaron en laboratorio.
Se realizaron cruces intraespecíficos, en los cuales se evidenciaron heterocariotipos, que constituyen un material ideal para verificar las filogenias ya que las zonas que comprenden los lazos (loops) permiten, en la mayoría de los casos, detectar los puntos críticos que revelan las roturas que dieron origen a los arreglos que los conforman (Polanco 1982). Estos cruces intraespecíficos se muestran con mayor detalle en la tabla 1.

Se disecaron las glándulas salivares de las larvas en tercer estadio, provenientes de los cultivos de D. starmeri y de ios híbridos intraespecíficos y se les adicionó una gota de carnoy (o solución fijadora) por 1 minuto. Para la población de D. starmeri de la Tatacoa, el carnoy, se preparó en una proporción de 3:1 de etanol, ácido acético. Para la población de D. starmeri, Santa Marta y Camarones y para todos los híbridos, se utilizó carnoy "modificado" en una proporción de 4:3:1 de agua, etanol y ácido acético, respectivamente. Para la tinción se utilizó acetorceína láctica por tres horas, posteriormente se cubrió el preparado y se realizó el aplastamiento con un golpe de martillo de reflejos.
Una vez obtenidas las placas, se tomaron diversas fotografías del cromosoma 2 de cada una de las poblaciones muestreadas (Polanco 1998), a partir de las mismas se hizo el bandeo y se obtuvieron los mapas, que sirvieron como patrones para establecer las secuencias de inversión de cada población. El patrón básico de bandeo utilizado fue el mapa fotográfico de D. repleta (Wollaston) de Polanco (1998), por ser esta la única especie monomórfica del grupo repleta.
Posteriormente, se realizaron comparaciones con las diferentes propuestas presentadas por otros autores, para verificar la concordancia.
Para la elaboración de los mapas se utilizaron un escáner ScanJet 6200C y el programa Adobe FotoDeluxe 2.0, que permitió intensificar los contrastes, ampliar varias veces determinadas zonas para facilitar el bandeo, modificar la posición de las imágenes, rotándolas en la dirección que se desea, adicionar la nomenclatura de bandeo y, finalmente, imprimirlas con un máximo de fidelidad, con la ayuda de una impresora láser.
Por último, se planteó la posible ruta filogenética del cromosoma 2 deD. starmeri por el método de las inversiones traslapadas (también llamadas complejas), el cual presenta una gran ventaja, pues se puede seguir el rastro que dejan las diferentes inversiones en un cromosoma determinado a lo largo de la evolución del mismo, y en consecuencia se puede postular una ruta filogenética en la que se evidencian las sucesivas reordenaciones, mas no la dirección de la evolución (Polanco 1998; Sturtevan y Dobzansky 1986; Ruiz et al. 1992).
Resultados y Discusión
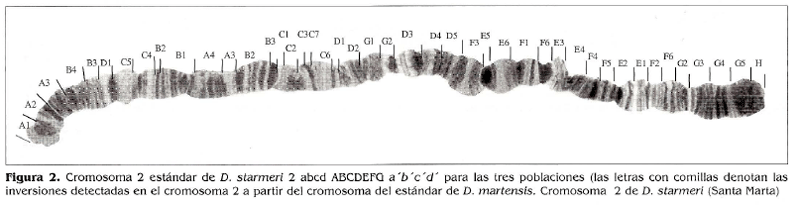
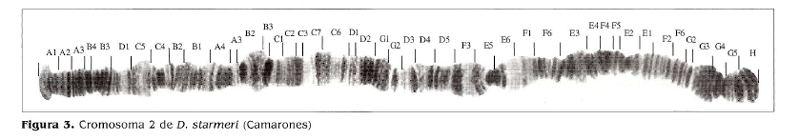
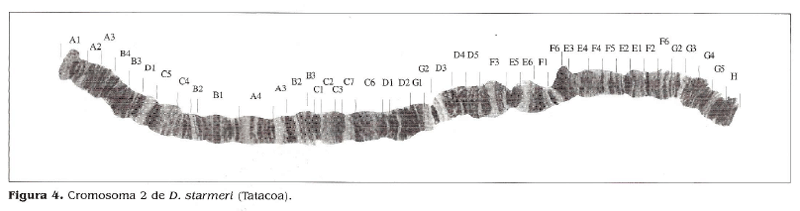
El resultado del mapeo de D. starmeri muestra un homocariotipo igual para las tres poblaciones, observándose un heterocariotipo para la población de Santa Marta, la cual es polimórfica para las inversiones/' y e' (Figs. 2,3,4).
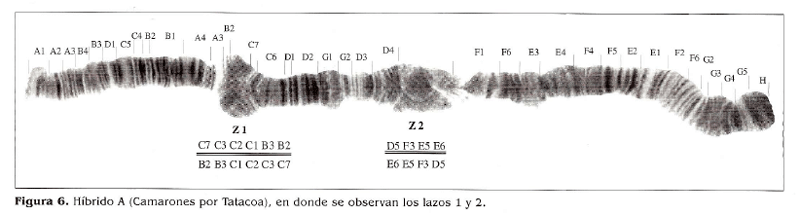
En los cromosomas 2 de los cruces híbridos A, a, B y b, se observó un heterocariotipo con 2 lazos. El mapeo de estos lazos da una secuencia que se repite en todos los cromosomas 2 híbridos para las dos zonas; determinándose, de forma exacta, los puntos de rotura de las inversiones. De acuerdo con esto, las secciones implicadas en estas dos inversiones son:
Las fórmulas de cada uno de los arreglos observados en los heterocariotipos híbridos A y B, son:
Tátacoa/Santa Marta-Camarones: B^CjC, C A /C^C^B, , D5F3E5E6 / E6E5F3D5.
En la comparación de los mapas de las tres poblaciones, con las secuencias estándar señaladas por Wasserman y Koepfer (1979), Ruiz y Fontdevila (1981) y Ruiz y Wasserman (1993), se evidencia una amplia diferencia, tanto con el arreglo estándar como en los polimorfismos observados en este estudio. El análisis realizado a la ruta filogenética del cromosoma 2 estándar de D. martensís registrada por Folanco (1998), muestra que la misma se ajusta bien a la filogenia del cromosoma 2 de D. starmeri y, teniendo en cuanta la cercanía evolutiva entre las dos especies, se ha tomado como base para proponer la evolución del cromosoma 2 en D. starmeri.
El cromosoma 2 estándar de D. starmeri propuesto en este trabajo, se originó por cuatro inversiones (a',b',c',yd')a partir del cromosoma 2 estándar de D. martensis propuesto por Polanco (1998) (Fig. 8).
La filogenia muestra que el arreglo estándar del cromosoma 2 de D. starmeri es 2ABCDEFGabcda 'b'c'd' y existen además, otros tres arreglos en el mismo cromosoma:
2ABCDEFGabcda'b'c'd'e', 2ABC DEFGabcda'b'c'd'e'f', 2ABCDEF G a b c d a'b'c'd'f.
Los cruces A y B (Tabla 1) muestran una posible divergencia a nivel cromosómico entre las poblaciones de la costa norte colombiana y la del Desierto de la Tatacoa, ya que en la descendencia híbrida se observaron lazos que evidencian el polimorfismo presente en el cromosoma 2.
Estos resultados concuerdan con los análisis morfológicos realizados por Castañeda (2000) y de aislamiento reproductivo realizado por Rojas (2001) en los cuales se manifiesta un grado de divergencia genética entre las poblaciones de la costa norte y de la Tatacoa. En las poblaciones de Santa Marta y Camarones no se observan diferencias a nivel cromosómico, lo cual es consistente con lo observado por Rojas (2001), aunque existe un grado de polimorfismo, señalado en diferentes investigaciones como las de Castañeda (2000), Sánchez (1987) y Hoenigsberg eí al. (1987).
Conclusiones
- Existe un grado de divergencia en las ordenaciones del cromosoma 2, en las poblaciones de la costa norte y el Desierto de la Tatacoa, representado en el arreglo estándar y tres inversiones diferentes.
- La ruta filogenética nueva planteada para el cromosoma 2 de D. starmeri, de las poblaciones muestreadas, se deriva de la de Polanco (1998) por cuatro inversiones traslapadas.
- Los resultados de los análisis cromosómicos son consistentes con los de Castañeda (2000) y Rojas (2001) y confir-
man la divergencia entre las poblaciones de los dos ecosistemas.
Agradecimientos
Los autores agradecen la colaboración y paciente revisión de la profesora Adriana P. Rojas. M. Se. de la Facultad de Ciencias de la Universidad del Tolima y la financiación de la investigación por parte del Comité Central de Investigaciones de la Universidad del Tolima y el Instituto de Genética de la Universidad de los Andes.
Literatura citada
CASTAÑEDA, H. C. 2000. Análisis morfológico comparativo en Drosophila starmeri de las poblaciones de Camarones (La Guajira), Santa Marta (Magdalena) y Tatacoa (Desierto de la Tatacoa, Huila). Bogotá, (Tesis M. Se. Universidad de los Andes, Instituto de Genética). 192 p. [ Links ]
HOENIGSBERG, H. F.; MONTAÑO, A.; MORENO-ROSSI, A.; SANZ DE LA ROSA, M.; ORDÓÑEZ, M,; DE BUENDÍA, G. 1987. Genética de poblaciones en el Trópico Americano XXXIV. Las enzimas diagnósticas del enjambre matrensis del Grupo repleta de Drosophila de La Guajira Colombiana. Evolución Biológica 1: 297-334. [ Links ]
POLANCO, M. M. E. DE. 1982. Factores genético-evolutivos determinantes de la micro-evolución de los arreglos de las inversiones paracéntricas del III cromosoma de Drosophila pseudoobscura del altiplano de Bogotá, D. E. Colombia. Bogotá. (Tesis M. Se. Universidad de los Andes, Instituto de Genética). 485 p. [ Links ]
POLANCO, M. M. E. DE. 1998. Estudios cromosómicos comparados de Drosophila repleta (cepa de siboney Cuba) Vs Drosophila repleta (Warthon 1942) y Drosophila martensis (Valledupar, Barrancas y Riohacha). Bogotá. (Tesis Doctorado Genética de Poblaciones. Tomo I. Universidad de los Andes, Instituto de Genética). 272 p. [ Links ]
POLANCO, M. M. E. DE. 1999. Papel de las inversiones cromosómicas en la evolución de las especies del género Drosophila. Memorias II Congreso nacional de estudiantes de Bacteriología (ACEBEC). p: 27-32. Medellín. Colombia. [ Links ]
ROJAS, M. A. P. 2001. Análisis del aislamiento reproductivo en las especies cactofílicas del enjambre martensis, en dos regiones colombianas (Costa riorte y Desierto de la Tatacoa). Bogotá. (Tesis M. Se. Universidad de los Andes, Instituto de Genética). 198 p. [ Links ]
RUIZ,A.;FONTDEVILA,A. 1981. Ecología y evolución del subgrupo mulleri de Drosophila en Venezuela y Colombia. Acta Científica Venezolana VI. 32: 338-345. [ Links ]
RUIZ, A.; WASSERMAN, M. 1993. Evolutionary cytogenetics of the Drosophila buzzatii species complex, Heredity 70 : 582-596. [ Links ]
RUIZ, A.; FOIYTDEVILA, A.; WASSERMAM, M. 1992. The evolutionary history of Drosophila buzzatii. III. Citogenetic relation-ships between two sibling species of the buzaatii Cluster. Genetics 101: 503-518 [ Links ]
SÁNCHEZ, A. B. 1987. Relaciones filogenéticas en los Clusters buzzatii (grupo repleta) y martensis de Drosophila. Barcelona. (Tesis Doctoral. Publicacions de la Universitat Autónoma de Barcelona. Facultat de Ciénces). 458 p. [ Links ]
STURTEVAM, A.; DOBZHANSKY, TH. 1986. Inversions in the third chromosome of wild races of Drosophila pseudoobscura, and their use in the study of the species. Proc. Nati. Acad. Sci. 22: 448-450 [ Links ]
WASSERMAM, M. 1954. Cytological studies of the repleta group. The University of Texas Publ. 5422: 130-152 [ Links ]
WASSERMAN, M. 1982. Evolution and specia-tion in selected groups: the repleta species group. The genetics and biology of Drosophila. Ashburner (ed.) Vol. 3b: 61 - 139. [ Links ]
WASSERMAN, M.; KOEPFER. 1979. Cytogenetics of the South American Drosophila mu-llerl complex: The martensis cluster, more interespecific shaving of inversions. Genetics 93: 935-946. [ Links ]
Recibido: May. 15/ 2002 Aceptado: Ago. 11 / 2002


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}