










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488On-line version ISSN 2665-4385
Rev. Colomb. Entomol. vol.29 no.1 Bogotá Jan./June 2003
Efecto tóxico de proteínas Cryl de Bacillus thuringiensis sobre larvas de Teda solanivora (Lepidoptera Gelechiidae)
Bacillus thuringiensis Cryl proteins toxic effect against Teda solanivora (Lepidoptera: Gelechiidae)
WILSON MARTÍNEZ O.1, DANIEL UR1BE V.2, JAIRO CERÓN S.3
;1Autor para correspondencia: M.Sc. en Entomología. Laboratorio Entomología, Instituto de Biotecnología, Universidad Nacional de Colombia. A. A. 14490 Bogotá. E-mail: wilmarti@ibun.unal.edu.co
2M.Sc. Protección de cultivos. Qrupo de biopesticidas. Instituto de Biotecnología, Universidad Nacional de Colombia. E-mail: duribe@ibun.unal.edu.co
3Ph.D. Biotecnología. Qrupo de biopesticidas, Instituto de Biotecnología, Universidad Nacional de Colombia. E-mail: jaceron@ibun.unal.edu.co
Resumen. La polilla guatemalteca de la papa Teda solanivora Povolny se ha convertido desde mediados de la década pasada en una de las principales plagas que afectan los cultivos de papa en Colombia. Las estrategias de control de este insecto han involucrado, principalmente, el empleo de insecticidas químicos y en algunos casos métodos alternativos de control biológico y de prácticas culturales. El empleo de entomopatógenos como Bacillus thuringiensis y baculovirus presentan gran potencialidad para ser involucrados en programas de manejo integrado de T. solanivora. Algunos trabajos anteriores han evaluado el potencial de cepas nativas de B. thuringiensis para controlar esta plaga, sin embargo, aún no se han identificado cuál o cuáles proteínas Cry poseen actividad tóxica específica para la polilla guatemalteca de la papa. En el presente trabajo se analizó la actividad tóxica individual de las proteínas CrylAa, CrylAb, CrylAc, CrylBa, CrylCa, CrylDa y CryLEa de B. thuringiensis, en larvas de primer instar de T. solanivora empleando dieta natural como alimento. Se diseñó simultáneamente un método de bioensayo sencillo, que requiriera cantidades bajas de proteína y que garantizara su reproducibilidad. Las proteínas se evaluaron en concentración de 4 ug/cm2, que correspondió a la CL50 de la cepa HD1, incluida como control positivo en los bioensayos. Los resultados obtenidos indicaron una toxicidad mayor de la proteína CrylAc respecto de las demás proteínas evaluadas; se obtuvo un porcentaje de mortalidad promedio cercano al 20% con dicha proteína, mientras que los porcentajes de mortalidad con las demás proteínas no superaron el 7%. Con el fin de confirmar los resultados obtenidos, se realizaron bioensayos con la cepa HD73, la cual posee únicamente los genes CrylAc y Cry2A y la cepa recombinante de E, coli que posee el gen Cry2A. Como se esperaba, los resultados indicaron una actividad tóxica alta de la cepa HD73 y baja de la proteína Cry2A, obteniéndose mortalidades superiores al 70% y menores del 10%, respectivamente. Lo anterior evidenciaría que la proteína CrylAc presenta actividad tóxica específica contra T. solanivora; sin embargo, parece que la interacción con otras proteínas Cryl ocasiona una mayor toxicidad de dicha proteína, teniendo en cuenta los mayores porcentajes de mortalidad promedio respecto de la proteína individual, observados con las cepas HD1 e HD73, en las cuales la proteína CrylAc se encuentra en combinación con otras proteínas Cryl.
Palabras clave: Polilla guatemalteca. Bioensayos. Toxinas. Control biológico.
Summary. In the last decade Teda solanivora Povolny has become one of the most significant pest in Colombian potato growing áreas. In the past T. solanivora control strategies have been developed using chemical pesticides, although recently some alternative biológica! control and crop management practices have been used. The use of entomopathogenic microorganisms such as baculovirus and bacteria for T. solanivora control offers a new useful tool for developing an intégrate pest management program, Earlier works explored the potential use of Badllus thuringiensis strains in T, solanivora control. However, Bt Cry proteins specific to this insect have not been yet identified. In accordance with this, we analyzed the specificity of 7 Bt Cryl proteins (CrylAa, CrylAb, CrylAc, CrylBa, CrylCa, CrylDa and CrylEa) toward the first instar T. solanivora larvae. At the same time we designed an easy and reproducible bioassay method based in natural diet. Proteins were evaluated at 4 ug/cm2 dose, which was the LC50 of HD1 strain, used as a positive control in bioassays. Results showed a higher toxicity of CrylAc compared with the other Cryl proteins evaluated. CrylAc caused a 20% average mortality while no more than 7% were obtained with the rest of the proteins. In order to confirm these findings we evaluated HD73 strain which posses only CrylAc and Cry2A genes and recombinant E. coli strain with only Cry2A gene. As we expected, the bioassays indicated a high toxicity of HD73 and low toxicity of Cry2A protein, showing average moralities over 70% and below 10%, respectively. Results indicated that CrylAc is specific to T. solanivora, however, it seems that interaction with other proteins enhances CrylAc activity as observed in bioassays with HD1 and HD73 strains where Cryl proteins besides CrylAc are present.
Key words: Polilla guatemalteca. Bioassays. Toxins. Biological control.
Introducción
La polilla guatemalteca de la papa Tecla solanivora Fovolny, desde su introducción a Colombia en 1983, se ha constituido en uno de los principales factores de pérdidas en el rendimiento de dicho cultivo. Esta plaga se registró por primera vez en 1985 y desde esa época se han incrementado sus poblaciones hasta tal punto que a finales del siglo pasado fue una de las causas de las pérdidas cercanas al 100% del total de la producción de papa en el país (Ñústez et al. 1998). En Boyacá y Cundinamarca, que son las principales zonas productoras de papa de Colombia, se han estimado daños entre el 50 y 80% en los tubérculos tanto en campo como en almacenamiento (Hernández y Várela 1998). Actualmente esta plaga se encuentra distribuida en todas las zonas productoras de papa del país y su control se ha hecho empleando principlamente insecticidas químicos con eficiencias de control menores del 50% (Andrew y Acosta 1998).
Teniendo en cuenta las limitaciones de la estrategia química para el control de T. solanivora, se han venido proponiendo otras alternativas de control tendientes al desarrollo de un esquema de manejo integrado de la plaga, involucrando elementos de control cultural, etológlco y biológico (fiústez et al. 1998). Dentro de las estrategias de control biológico se ha explorado el uso de microorganismos entomopatógenos como virus (Arias et al. 1998; Trillos 1998), hongos (Sánchez et al. 1998), nemátodos (Alvarado et al. 1998; Parada^y Luque 2001) y bacterias, estas últimas evaluando cepas nativas y/o productos comerciales con la bacteria Bacillus thunnglensis, lográndose identificar cepas nativas y productos con alguna actividad tóxica contra T. solanivora (Aristizábal y Londoño 1998; Pérez et al. 1997; Tejada et al. 1998; Bosa et al. 1998, Castelblanco 2000).
B. thuringiensis es una bacteria entomopatógena con características propias que la hacen promisoria para ser introducida en programas MIP de la polilla guatemalteca de la papa; a pesar de esto, aún no se conoce cuál de las proteínas tóxicas que produce dicha bacteria es específica hacia este insecto. Se hace por lo tanto necesario realizar estudios al respecto, que permitan identificar cuál proteína Cry de B. thuringiensis presenta una actividad tóxica específica y así definir y optimizar su empleo como agente de control biológico de T. solanivora, ya sea empleándola como ingrediente activo de productos comerciales para aplicación directa o como fuente de genes para el desarrollo de plantas transgénicas resistentes a este insecto (Estrada 1997).
De acuerdo con lo anterior, el presente trabajo se orientó hacia la identificación de la o las proteínas Cry de B. thuringiensis con actividad tóxica específica hacia T. solanlvora, empleando para tal fin siete proteínas Cryl y dos cepas de referencia de esta bacteria.
Materiales y Métodos
Material entomológico
El material entomológico se obtuvo a partir de una cría de T. solanivora mantenida bajo condiciones de laboratorio (60% HRy 15°C, en promedio), en el laboratorio de entomología del Instituto de Biotecnología de la Universidad Nacional de Colombia, en dieta natural consistente en tubérculos de papa pastusa. Los adultos se mantuvieron en frascos de vidrio y se alimentaron con una solución de agua-miel impregnada en un algodón. Las oviposiciones se recolectaron semanalmente y se colocaron en recipientes plásticos para esperar la emergencia de las larvas. Las larvas neonatas se emplearon en los bioensayos y para el mantenimiento de la cría.
Cepas bacterianas
Los bioensayos se realizaron empleando tres tipos de cepas bacterianas: una cepa de Bt de referencia, una cepa nativa y siete cepas recombinantes de Escherlchia co/í que poseen genes Cryl individuales. Las cepas de referencia empleadas, HDl y HD73 poseen los genes crylAa, crylAb y crylAc la primera y crylAc y cry2A la segunda. Las cepas de E. coli presentan los genes crylAa, crylAb, crylAc, cryTBa, crylCa, crylDa, crylEa y Cry2A. Cada gen codifica para su correspondiente proteína Cry. Se seleccionaron estas últimas cepas por poseer de forma individual tanto los genes cry, que presenta en combinación la cepa de referencia HDl y que han sido identificadas como activos hacia larvas de primer instar de T. solanivora (Pérez et al. 1997; Castelblanco 2000), como otros que potencialmente pueden tener actividad tóxica contra este insecto.
Todas las cepas se crecieron en medio líquido Luria Bertani. Las cepas de referencia se hicieron crecer hasta alcanzar una esporulación del 95%, recuperándose la biomasa por centrifugación y obteniéndose la mezcla espora-cristal después de sucesivos lavados con agua destilada-estéril. Las proteínas producidas por las cepas recombinantes se purificaron siguiendo el método de Regev et al. (1996). La proteína obtenida en ambos casos se cuantificó con el método de Lowry (Perbal 1988).
Método de bioensayo
El método del bioensayo se estandarizó mediante la realización de ensayos preliminares con la cepa de referencia HDl, en los cuales se evaluó la mortalidad de las larvas, la reproducibilidad del ensayo, la facilidad de realización y la cantidad de proteína requerida para realizar el bioensayo.
Se evaluaron tres tipos de métodos: El primero consistió en la inmersión de prismas de papa de 0,25 cm3 en las muestras a evaluar por 5 min en agitación constante. Luego los prismas se dejaron secar al ambiente y se colocaron en placas para cultivo celular de 24 pozos. En cada pozo se puso un prisma de papa y se infestó con una larva de T. solanivora. El segundo método consistió en un cilindro de papa de 1 cm de diámetro por 1,5 cm de alto, dispuesto en un vaso des-echable y rodeado con parafina o agar para disminuir la superficie de alimentación del insecto. Dicha superficie se contaminó con las muestras a evaluar. Una vez seca la muestra se infestó la papa con 1 larva de T. solanivora. En el último método se empleó una dieta artificial preparada con puré de papa casero o puré comercial, adicionándole sustancias preservantes y agar para darle consistencia a la dieta (Pérez et al. 1997). La dieta se dispensó en placas de 24 pozos similares a las del primer método y se contaminó superficialmente con la muestra. Una vez seca la muestra, se infestó cada pozo con una larva de T. solanivora.
Cada ensayo se realizó por duplicado empleando 30 larvas de primer instar de T. solanivora por cada réplica. La lectura de los bioensayos se realizó a los 10 días, contabilizando el número de larvas muertas en cada método evaluado.
Determinación de las concentraciones letales de la cepa HDl
Empleando el método de bioensayo seleccionado se realizaron bioensayos para determinar las concentraciones letales 10, 50 y 90 de la cepa de referencia HDl. Para tal fin, se realizó un bioensayo empleando 7 dosis de la cepa y un testigo relativo con agua destilada. Se emplearon 48 larvas por tratamiento con dos repeticiones cada uno y el ensayo se hizo por duplicado. Se realizó la lectura del bioensayo a los 7 días, determinando el número de larvas muertas por dosis. Los datos obtenidos se procesaron mediante el paquete estadístico PROBIT POLO-PC.
Evaluación de las proteínas Cryl
Las proteínas Cryl y la cepa HD73 se evaluaron con el método seleccionado, empleando la dosis obtenida como CL50 de la cepa de referencia HDl. En cada ensayo se emplearon 48 larvas por tratamiento y tres repeticiones de cada uno. Así mismo, se utilizó un tratamiento testigo consistente en la solución buffer empleada en la purificación de las proteínas evaluadas. La lectura del bioensayo se realizó a los 7 días contabilizando el número de larvas muertas y los respectivos porcentajes de mortalidad con cada proteína. Se empleó un diseño de bloques al azar. Los datos obtenidos como número de individuos muertos por tratamiento se transformaron como -vX+1. Los datos transformados se sometieron a análisis de varianza y prueba de comparación múltiple de Duncan con a de 0.05.
Resultados y Discusión
Método de bioensayo
El método de inmersión de los prismas en la suspensión de la muestra se evaluó empleando una suspensión de liofiiizado de B. thuringiensis, cepa HDl, a las concentraciones de 25 y 100 ug/ml. Los valores promedios de mortalidad obtenidos correspondieron a 68,3 ug/ml y 91,4 ug/ml, respectivamente. El análisis de varianza indicó que no hubo diferencias significativas entre las réplicas pero sí entre las dosis (P<0.01, n=30). El coeficiente de variación para el bioensayo fue de 8,2%, siendo aceptable para los bioensayos con B. thuringiensis (Dulmage 1981; Lecuona 1996). De acuerdo con los resultados obtenidos, este método de bioensayo presentó buenas perspectivas para ser empleado en las evaluaciones posteriores.
En los ensayos con parafina se emplearon 60 unidades por tratamiento y el mismo número de larvas de primer instar de T. solanivora. En el ensayo con paraflna la preservación de la dieta fue buena en todos los tratamientos y hubo un coeficiente de variación aceptable del 9,0%. Sin embargo, no se observaron diferencias entre las dosis evaluadas (2,5 y 25 ug de proteína/cm2) y el testigo (P<0,01, n=60). Esto se debió a que en algunos casos se presentó separación de la parafina de la papa, dejando espacios por donde las larvas pudieron consumir dieta sin toxina. Así mismo, este método resultó bastante dispendioso lo cual causa errores de manipulación y por consiguiente un incremento en el error experimental.
En el ensayo con agar la dieta también se preservó muy bien y el coeficiente de variación fue del 10,9%. Igual que como sucedió con el método anterior, no se observaron diferencias significativas entre las dos dosis evaluadas y el testigo (P<0,01). Además, este método resultó dispendioso, lo cual disminuye la reproducibilidad y sensibilidad del mismo.
Los resultados obtenidos en los bioensayos que emplearon puré de papa casero o comercial, dieron resultados muy variables no habiendo reproducibilidad entre las repeticiones. Por otro lado, estos ensayos presentaron los porcentajes más altos de mortalidad en el testigo, superando el 30% y no permitieron un desarrollo normal de las larvas, como lo debería hacer una dieta artificial adecuada. Lo anterior hizo que los resultados obtenidos no fueran confiables por lo que no fueron tenidos en cuenta para los posteriores análisis. Los resultados obtenidos con este tipo de bioensayo no con-cuerdan con los obtenidos por Pérez et al. (1997), quienes utilizando una dieta similar, realizaron bioensayos con cepas nativas de B. thuringiensis y larvas de primer instar de T. solanivora. Aunque ellos señalaron que el 90% de las larvas sobrevivieron los 7 días de la evaluación, no se especifica si la dieta realmente soportó el desarrollo de T. solanivora durante todo su ciclo de vida. Este debería ser el criterio esencial para el empleo de una dieta en evaluaciones de entomopatógenos, ya que se garantiza que en realidad no haya ningún efecto colateral sobre el desarrollo nutricional y fisiológico de un insecto que pueda influir en los resultados obtenidos.
La variabilidad de los resultados obtenidos y específicamente la mortalidad alta en el testigo se atribuyen a características de las dietas a base de puré, que difieren mucho de la dieta natural, en características como textura, olor, nutrientes y presencia de sustancias preservantes en el caso del puré comercial. De acuerdo con los resultados, los métodos de bioensayo que emplean puré como sustrato alimenticio se descartaron. Por lo tanto, en el presente trabajo se decidió realizar los bioensayos empleando únicamente dieta natural.
Los resultados obtenidos con la evaluación de los diferentes métodos de bioensayo indicaron que el mejor método para la evaluación de las cepas y proteínas de B. thuringiensis es el que emplea prismas de papa, porque garantiza la reproducibilidad de los resultados y es lo suficientemente sensible como para detectar diferencias entre los tratamientos.
Una vez seleccionado el método general de bioensayo se realizó un ajuste respecto a la forma de contaminar los prismas de papa con las muestras a evaluar. Debido a que en el método inicial se embebían todos los trozos simultáneamente, se necesitaba gran cantidad de muestra, lo cual en el caso específico de las proteínas recombinantes resultaba difícil de obtener. Por lo tanto, se optimizó este aspecto realizando la contaminación directa de los prismas de papa con micropipeta y un volumen de muestra tal, que permitiera impregnar homogéneamente el prisma de papa.
Concentraciones letales de la cepa de referencia
En la determinación de las concentraciones letales de la cepa de referencia HDl se empleó un rango de dosis entre 0,5 y 10 |ig de proteína por cm2 de dieta. Los resultados promedio obtenidos en los bioensayos realizados se observan en la figura 1.
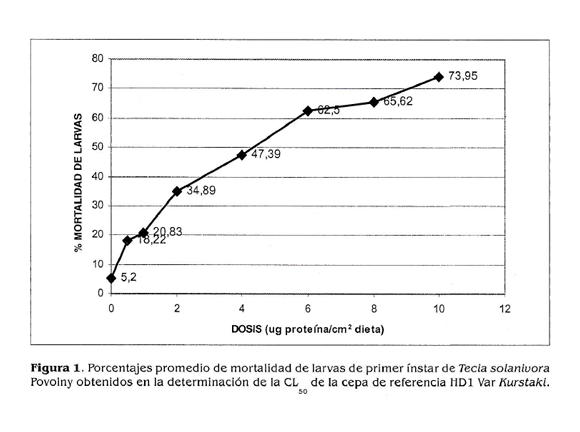
Los datos obtenidos en los bioensayos se procesaron mediante el paquete estadístico PROBIT POLO PC, encontrándose como CL50 de la cepa de referencia HDl un valor de 4,37 Ug de proteína/ cm2 de dieta (Tabla 1).
Si se tienen en cuenta los datos de CL50 para la cepa HDl registrados por Pérez eí ai. (1997) y se convierten a las mismas unidades que se emplearon en este trabajo (jig/ cm2), se encuentra que el valor obtenido es mayor al señalado por dichos autores, quienes obtuvieron una CL50 de 2,84 ug/cm2. Sin embargo, ya que dicha diferencia no es muy amplia, puede ser atribuible a los diferentes métodos de bioensayo empleados en ambos trabajos, ya que factores como el tipo de alimento utilizado y diferentes métodos de suministro de las proteínas en la dieta pueden originar respuestas diferentes en el insecto empleado.
Evaluación de las proteínas Cryl
El análisis de varianza de los datos obtenidos en los bioensayos con las proteínas Cryl evaluadas, indicó diferencias altamente significativas entre los tratamientos (P<0,01). Los valores promedio de mortalidad obtenidos oscilaron entre 19,26% para la proteína CrylAc, que resultó la proteína más tóxica y 1,56% para la proteína CrylD, que fue la menos tóxica. Las proteínas CrylAa, CrylAb, CrylB y CrylC presentaron porcentajes de mortalidad promedio de 6,24, 6,76, 4,16 y 5,72, respectivamente. El análisis de Duncan indicó diferencias entre los promedios de individuos muertos obtenidos con la proteína CrylAc y las demás proteínas evaluadas (cc=0.05). Las mortalidades promedio de las proteínas CrylAa, CrylAb, CrylB, CrylC y Cry2A fueron similares entre sí y las proteínas CrylE y CrylD fueron diferentes a las anteriores, pero similares entre sí (a = 0.05) (Fig. 2). Estas dos últimas proteínas fueron las menos tóxicas para las larvas de T. solanivora, no mostrando diferencias estadísticas (a=0.05) con el testigo empleado en los bioensayos, el cual tuvo una mortalidad promedio de 3,12%.
Los datos anteriores indican que la proteína CrylAc presentó la actividad tóxica más alta contra las larvas de T. solanivora. Pérez et al. (1997) propusieron que las proteínas CrylAa, CrylAb, CrylAc de la cepa HDl y las proteínas CrylAb, CrylB y CrylD de una cepa nativa denominada Bt-40 que evaluaron eran las responsables de la actividad de dichas cepas; sin embargo, por tratarse de cepas con genes múltiples no pudieron determinar exactamente la proteína responsable de la toxicidad. Los datos obtenidos en el presente trabajo permiten proponer que la proteína CrylAc ejerce un papel fundamental en la mayor actividad tóxica de la cepa HDl (CL50 de 56,9 |ig/ml), respecto de la cepa nativa Bt-40 (CL50 de 95,52 ug/ml).
Por otro lado, la proteína CrylAb puede ser la responsable de la actividad tóxica de algunas cepas nativas, ya que esta fue la segunda proteína más activa de los bioensayos realizados causando mortalidades hasta del 10%. Esto estaría de acuerdo con lo propuesto por Castelblanco (2000), quien evaluó varias cepas nativas de B. thuringiensis que presentaban genes Cryl y concluyó que la actividad tóxica de algunas de las cepas se debía a la presencia del gen CrylAb. Igualmente, se puede relacionar la toxicidad observada con la proteína CrylAb con los valores altos de mortalidad (>90%) de larvas de P operculella, obtenidos por Cañedo y Lagnoui (2001), cuando evaluaron plantas transgénicas de papa parda pastusa a las cuales se les introdujo el gen crylAb de B. thuringiensis. Lo anterior, debido a las similitudes de P operculella con T. solanivora, por las cuales dichos autores proponen la evaluación de la resistencia de estas plantas transgénicas hacia T. solanivora.
La corroboración de la actividad tóxica específica de la proteína CrylAc se obtuvo con los ensayos realizados con la cepa HD73 (con genes crylAc y cry2A, únicamente) y la cepa E. coli recombinante con el gen cry2A (Fig. 3). En dichos bioensayos, empleando la CL50 de la cepa HDl, la cepa HD73 presentó una mortalidad promedio de 77,6%, atribuible principalmente a la presencia de la proteína CrylAc, según los resultados preliminares. Se descarta un efecto tóxico causado por la proteína Cry2Aya que en las evaluaciones preliminares con esta proteína, en forma individual, se obtuvo un valor de mortalidad promedio de tan sólo el 5%.
Es importante tener en cuenta que en algunos casos las proteínas Cry presentes en una cepa determinada pueden estar expresadas de forma diferencial constituyendo cada una un porcentaje diferente del total de proteína presente en el cristal proteico. De esta forma, y debido a que en la mayoría de cepas nativas de B. thuringiensis existe más de una proteína Cry presente, es de esperarse que la actividad tóxica varíe, aunque las cepas contengan el mismo tipo y número de genes cry. Por lo tanto, esto explicaría la variabilidad de los resultados obtenidos en los trabajos realizados hasta el momento con cepas nativas de B. thuringiensis (Pérez et al. 1997; Castel-blanco 2000), que aunque poseen los mismos genes identificados por técnicas moleculares, su toxicidad hacia larvas de T. solanivora no es la misma.
Por otro lado, la presencia o ausencia de una protoxina particular CrylA puede afectar de forma marcada la solubilidad de todo el cristal proteico, debido posiblemente a las uniones disulfuro presentes entre estas protoxinas relacionadas, afectando así la disponibilidad de un protoxina dentro del intestino larval y por consiguiente la efectividad tóxica del cristal completo (Aronson 1995). Lo anterior también podría explicar la variación en la toxicidad de cepas nativas que poseen el gen Cry lAc en diferentes combinaciones con otros genes tipo Cryl.
Los resultados aquí presentados representan un aporte para el desarrollo de nuevas estrategias biológicas para el control de la polilla guatemalteca de la papa. Surge así la recomendación que aquellas cepas nativas que contengan en su genoma el gen crylAc y expresen dicha proteína, son susceptibles de ser evaluadas como potenciales ingredientes activos de bioplaguicidas para el control de T. solanivora en papa en almacenamiento; así mismo, dicho gen debe ser tenido en cuenta para ser empleado en aquellos trabajos que busquen el desarrollo de plantas de papa resistentes a esta plaga. Adicionalmente, se puede continuar la exploración de otros genes potencialmente útiles para el control de esta plaga, como el caso del gen cry5 el cual causó mortalidades hasta del 100% en larvas de Phothorimaea opercullella Zeller que atacaron tubérculos de papa transformada genéticamente con dicho gen (Mohammed et al. 2000).
Conclusiones
- Se determinó que la proteína CrylAc de B. thuringiensis presenta la mayor actividad tóxica específica hacia larvas de primer instar de T. solanivora.
- Se evidenció que la actividad tóxica hacia T. solanivora de la proteína CrylAc, en algunos casos, puede ser incrementada por la presencia de otras proteínas Cryl.
- Se desarrolló un método de bioensayo basado en dieta natural reproducible, sensible y fácil para realizar bioensayos de evaluación de la toxicidad de proteínas y/o cepas de B. thuringiensis hacia T. solanivora.
Agradecimientos
Los autores agradecen al estudiante de Ingeniería Agronómica Alejandro Aranguren por su apoyo en la realización de los bioensayos y mantenimiento de la cría del insecto. Igualmente, al Instituto de Biotecnología de la Universidad Nacional de Colombia y a Colciencias por la financiación de este trabajo.
Literatura citada
ALVARADO, B.; ENRÍQUEZ, S.; BECERRA, O. 1998. Patogenisidad de Steínernema feltiae Cepa Villapinzón, sobre Tecia solanivora atacando tubérculos de Solanum tuberosum variedad sabanera. Memorias. Seminario. Perspectivas sobre nemátodos fitopatógenos y entomopató-genos en Colombia, p. 40. Bogotá [ Links ].
ANDREW, R.; ACOSTA, A. 1998. Mesa de trabajo sobre control químico de Tecia solanivora. Memorias. Taller de planeación estratégica para el manejo de Tecia solanivora en Colombia, p. 23. Bogotá [ Links ].
ARIAS, J.; TRILLOS, O; OSORIO, G. 1998. Adaptación de tecnología para la formulación y producción del insecticida biológico Baculovirus pthorimae para el control de la polilla guatemalteca de la papa Tecia solanivora Povolny. Memorias. Taller de planeación estratégica para el manejo de Tecia solanivora en Colombia, p. 3. Bogotá [ Links ].
ARISTIZÁBAL, C; LONDOÑO, M. 1998. Pruebas de patogenicidad de aislamientos nativos de Bacillus thuringiensis sobre la polilla de la papa Tecia solanivora Povolny. Memorias. Taller de planeación estratégica para el manejo de Tecia solanivora en Colombia. p. 31. Bogotá [ Links ].
ARONSON, A. 1995. The protoxin composition of Bacillus thuringiensis insecticidal inclusions affects solubility and toxicity. Applied and Environmental Microbiolgy 61 (11): 4057-4060. [ Links ]
BOSA, F.; PRÍAS, J.; COTES, A. 1998. Evaluación de la actividad insecticida de cepas nativas de Bacillus thuringiensis para el control de larvas de Tecia solanivora Povolny (Lepidoptera: Gelechiidae). Memorias. Taller de planeación estratégica para el manejo de Tecia solanivora en Colombia. p. 50. Bogotá [ Links ].
CAÑEDO, V.; LAGNOUI, A. 2001. Evaluación del efecto de las plantas transgénicas en el desarrollo de las polillas de la papa Pthorimaea operculella (Zeller), Symme-trischema tangolias (Oyen) y Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). Resúmenes XXVIII Congreso Sociedad Colombiana de Entomología, p. 15. Pereira. [ Links ]
CASTELBLANCO, A. 2000. Diseño de una metodología para evaluar la actividad biopla-guicida de cepas nativas de Bacillus thuringiensis contra larvas de primer instar de Tecia solanivora Povolny en laboratorio. Trabajo de Grado. Facultad de Agronomía. Universidad Nacional de Colombia. Bogotá. 63 p. [ Links ]
DULMAGE, H. 1981. Insecticidal activity of ¡so-lates of Bacillus thuringiensis and their potential for pest control, p. 193-222. En: H.D. Burges (Ed.). Microbial control of pest and plant diseases 1970-1980. Academic Press. London. England. [ Links ]
ESTRADA, N. 1997. Resistencia genética como una alternativa de manejo de la polilla de la papa. Resúmenes. Seminario experiencias y avances en el manejo de la polilla guatemalteca de la papa. p. 3. Bogotá [ Links ].
HERNANDEZ, L.; VÁRELA, A. 1998. Dinámica poblacional y comportamiento de la polilla guatemalteca Tecia solanivora Povolny en el municipio de Ventaquemada, Boyacá. Memorias. Taller de planeación estratégica para el manejo de Tecia solanivora en Colombia, p. 34. Bogotá [ Links ].
LECUONA, R. 1996. Microorganismos patógenos empleados en el control microbiano de insectos plaga. Universidad de Buenos Aires. Buenos Aires. 338 p. [ Links ]
MOHAMMED, A.; DOUCHES, D.; PETT, W.; GRAFIUS, E.; COOMBS, J.; LI, W.; MADKOUR, M. 2000. Evaluation of potato tuber moth (Lepidoptera : Gelechiidae) resistance in tubers of Bt-cry5 transgenic potato Iines. Journal of Economic Entomology 93 (2): 472-476. [ Links ]
ÑÚSTEZ, C; ESTRADA, N.; BUITRAGO, G.; CARO, M.; PORRAS, P; NARANJO, A. 1998. Introducción. Memorias. Taller de planeación estratégica para el manejo de Tecia solanivora en Colombia, p. 2. Bogotá [ Links ].
PARADA, J.; LUQUE, E. 2001. Acción de Steínernema feltiae Filijev (Rhabditida: Steinerne-matidae) sobre Tecia solanivora Povolny (Lepidoptera: Gelechüdae), bajo condiciones de campo. Resúmenes XXVIII Congreso Sociedad Colombiana de Entomología. p. 31. Pereira. [ Links ]
PERBAL, B. 1988. A practical guide to molecular cloning. Second edition. John Wiley and Sons Ed. New York. USA. 547 p. [ Links ]
PÉREZ, O.; RODRÍGUEZ, A.; COTES, A. 1997. Método rápido y confiable para evaluar la actividad insecticida de cepas nativas de Bacillus thuringiensis contra Tecia solanivora Povolny (Lepidoptera: Gelechüdae). Revista Colombiana de Entomología 23 (3-4): 113-118. [ Links ]
REGEV, A.; KELLER, M.; STRIZHOV, N.; ZILBERS-TEIN, A. 1996. Synergistic activity of Bacillus thuringiensis delta-endotoxins and a bacterial endochitinase against Spodoptera litoralis larvae. Applied and Environmental Microbiology 62 (10): 3581-3586. [ Links ]
SÁNCHEZ, C; CÁRDENAS, A.; SOTELO, F.; VELANDIA, J. 1998. Identificación y evaluación de la patogenicidad de dos hongos aislados de larvas de polilla guatemalteca Tecia solanivora Povolny. Memorias. Taller de planeación estratégica para el manejo de Tecia solanivora en Colombia, p. 38. Bogotá [ Links ].
TEJADA, C; SORIANO, J.; VÁRELA, A. 1998. Evaluación de insecticidas en el manejo de Tecia solanivora Povolny en almacenamiento y en cultivos de papa Solanum tuberosum en los municipios de Motavita, TLinja y Subachoque. Memorias. Taller de planeación estratégica para el manejo de Tecia solanivora en Colombia. p. 52. Bogotá [ Links ].
TRILLOS, O. 1998. Evaluación de la factibilidad de producción semicomercial de Baculovirus pthorimae para el control de la polilla de la papa Tecia solanivora. Memorias. Taller de planeación estratégica para el manejo de Tecia solanivora en Colombia. p. 59. Bogotá [ Links ].
Recibido: Abr. 01 / 2002 Aceptado: Ago. 07 / 2002


{kind=link}
{kind=link}
{kind=link}