










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkRevista Colombiana de Entomología
versión impresa ISSN 0120-0488versión On-line ISSN 2665-4385
Rev. Colomb. Entomol. v.30 n.2 Bogotá jul./dic. 2004
Observacion de la patogenicidad de Metarhizium anisopliae en soldados Nasutitermes sp. (Isoptera: Termitidae)
Observation of the pathogenicity of Metarhizium anisopliae in Nasutitermes sp. soldiers (Isoptera: Termitidae)
ANA ISABEL GUTIERREZ G.1, YAMILLE SALDARRIAGA O.2
1Biologa. Investigador asociado al Grupo de Mitologia Instituto de Biologia, Facultad de Ciencias Exactas y Naturales. Corporacion de Patologias Tropicales. Universidad de Antioquia. E-mail: anaisaguti@hotmail.com
2Autor para correspondencia: Licenciada en Biologia y Quimica. M. Sc. Profesor de Micologia. Instituto de Biologia, Facultad de Ciencias Exactas y Naturales. Corporacion de Patologias Tropicales. Universidad de Antioquia. A. A. 1226. Medellin, Colombia. Fax: 2330120. E-mail: ysaldar@matematicas.udea.edu.co
Resumen. Soldados de la termita Nasutitermes sp., plaga del arbol del nogal cafetero Cordia alliodora (Ruiz & Pav.) Oken de la familia Boraginaceae, de gran importancia economica y medica, se inocularon in vitro con el hongo entomopatogeno Metarhizium anisopliae (Metsch.) Sor. a una concentracidn de 1x 107 conidios/ml del hongo impregnadas en lonas y papel de filtro. Observaciones del desarrollo de la infeccion del hongo fueron realizadas con el microscopio de luz. El desarrollo del hongo se observo en placas tenidas con azul de lactofenol, utilizando la tecnica de aclaracion en soluciones de KOH al 5, 10, 15 y 20%. Las termitas infectadas murieron de 1 a 3 dias post-inoculacion, el hongo penetrando la cuticula a las 24 h, colonizando el tejido graso a las 72 h e invadiendo los organos internos (intestino) despues de 72 h de la infeccion. A las 120 h masas compactas de conidioforos empezaron a emerger de la cuticula presentando coloracion blanca, ubicandose en las regiones intersegmentales de la superficie ventral (patas, torax) y partes bucales de los insectos. El micelio se torno verde a las 144 h de inoculacion, y a las 192 h se hizo mas oscuro y polvoso, invadiendo totalmente el cuerpo de la termita.
Palabras clave: Patogeno. Hyphomycete. Isoptera. Micosis. Inoculacion. Invasion.
Summary. Soldiers of Nasutitermes sp., pests of Cordia alliodora (Ruiz & Pav.) Oken, family Boraginaceae, a tree of great economic and medicinal importance were inoculated in vitro with the fungus Metarhizium anisopliae (Metsch.) Sor. impregnated on canvases or filter paper at a concentration of 1x107 conidias/ml. Observations on the development of the fungal infection were made with a light microscope. The development of the fungus was observed on mounted slides with lactofenol blue, using a clarifying technique with KOH solutions at 5, 10, 15 and 20%. Infected termites died I to 3 days post-inoculation, the fungus penetrating the cuticle at 24 hours, colonizing the fat tissue at 72 hours and invading the internal organs (gut) after 72 hours of infection. At 120 hours a white compact mass of conidiophores began to emerge from the cuticle at the intersegmental regions of the ventral surface (legs, thorax) and mouthparts of the insects. After 144 hours of inoculation the mycelia changed to green, and at 192 hours the mycelia turned dark and powdery when the termite body was totally invaded.
Key words: Pathogen. Hyphomycete. Isoptera. Mycosis. Inoculation. Invasion.
Introduccion
Las termitas se consideran como los insectos mas nocivos y destructores en las zonas templadas y tropicales. Atacan la madera, construcciones, plasticos, textiles, cauchos, cultivos agncolas y forestales (Garces 1997; Harris 1971).
En el bosque tropical, la mayoria de las termitas que se han encontrado, pertenecen a las familias KaloTermitidae, RhinoTermitidae o Termitidae, de las cuales la subfamilia Nasutitermitinae representa mas del 50% de las especies, siendo el genero Nasutitermes el mas abundante (Martius 1994). En Colombia se han registrado las tres familias mencionadas.
Nasutitermes spp., exopterigotas Neoptera son insectos sociales, representados en numerosos obreros y soldados no alados (Watson y Gay 1991; Borror et al. 1989). Atacan los arboles del nogal cafetero Cordia alliodora (Ruiz & Pav.) Oken de la familia Boraginaceae, que son de gran importancia economica y medica (Varon y Morales 1997).
Muchos autores han utilizado diferentes organismos en el control biologico de las termitas, como el virus de la polyhedrosis nuclear de Spodoptera littoralis (Hassan 1990), bacterias como Bacillus thuringiensis (Gavino 1984; Logan et al. 1990), nematodos de la familia Steinernematidae y del genero Steinernema carpocapsae (Coppel y Liang 1987; Mix y Beal 1985). Asi mismo, han sido probados con notable exito los hongos Beauveria bassiana y Metarhizium anisopliae (Yendol y Paschke 1965; Zoberi y Grace 1990; Sajap y Kaur 1990; Zoberi 1995; Alves et al. 1995; Milner et al. 1998; Delate et al. 1995).
Los hongos son uno de los agentes mas promisorios para el uso como insecticidas biologicos (Milner y Staples 1996). Entre los hongos probados como agentes de control bio!ògico en termitas estan Aspergillus flavus Entomophthora, Aspergillus sp. y Penicillium sp. (Espero 1984; L[j]utikova 1990). Ademas se utilizaron para controlar poblaciones de Stromatium fulvum (Villers) (Coleoptera: Cerambycidae) en plantaciones de Casuarina en Egipto (Hassan 1990). Igualmente M. anisopliae, Aspergillus fumigatus , Paecilomyces fumosoroseus y P. cicadae presentaron alta infectividad en termitas Coptotermes formosanus (Shiraki) bajo condiciones de laboratorio al aplicar suspensiones acuosas de 6,0 x 107 conidios/ml a una termita cebo (Chai 1995). Conidiobolus coronatus produjo una mortalidad del 100% sobre esta misma especie de termita despues de dos dias de aplicacion (Wells et al. 1995). Por otro lado, se comprobo que el ascomycete Cordycepioideus es patogeno para las termitas Macrotermes subhyalinus (Rambur) que construyen monticulos en Kenya (Ochiel et al. 1997).
Estudios en Australia demostraron que ciertos aislamientos de M. anisopliae pueden ser aplicados para matar colonias de termitas en termiteros, arboles y monticulos. El metodo consiste en capturar termitas, infestarlas con el patogeno y reintroducirlas al termitero para iniciar el contagio entre ellas (Milner y Staples 1996; Beron 1983). Ensayos in vitro para probar la patogenicidad de este hongo en termitas Microcerotermes en diferentes concentraciones demostraron que produce una mortalidad del 51% a una concentracion de 1 x 107 conidios/ml y del 80 -100% por encima de dicha concentracion (Zuluaga 2002).
M.anisopliae (Metsch.) Sor. es un hongo anamorfico (Kirk et al. 2001) aislado del escarabajo Anisoplia austriaca por Metchnikoff en 1879 y produce la muscardina verde en los insectos, por lo cual dicho hongo ha sido utilizado como agente microbiano contra insectos plaga (Tanada y Kaya 1993; Zimmermann, 1993; Steinhaus 1949). Es un hongo cosmopolita, saprofito, parasito de insectos, con un amplio rango de hospederos, que puede identificarse facilmente por sus conidioforos ramificados y por sus conidios cilindricos verdes, los cuales forman una capa densa y compacta. En el inicio del crecimiento las colonias presentan una coloracion blanca y las colonias maduras se tornan amarillentas, verde olivaceas, verde oscuras y crema rosaceas o vinaceas (Humber 1997; Tanada y Kaya 1993; Barnett 1967).
M.anisopliae posee dos variedades que se diferencian principalmente por el tamaño de sus conidios: la variedad mas comun anisopliae de conidios pequeños (3,5-9,0 um) y la variedad major de conidios grandes (9,0-18 um) (Kirk et al. 2001; Domsch et al. 1980). Este hongo crece facilmente en medios sintèticos solidos y liquidos, su temperatura optima de crecimiento oscila entre 25-30°C y sus conidios pueden sobrevivir liofilizados hasta por 16 años (Tanada y Kaya 1993; Samson 1981; Steinhaus 1949). Produce metabolitos secundarios como destruxinas A, B, C, D, E y una desmetildestruxina B, los cuales son toxicos y actuan como micotoxinas. Su toxicidad se da por la ingestion y a traves del integumento y su efecto varia segun la especie de insecto atacado, pudiendo causar paralisis tetanica y flacida in vivo, despolarizando membranas musculares al activar los canales de calcio; afecta varias organelas celulares (mitocondrias, reticulos endoplasmaticos y membranas nucleares), paraliza celulas y causa disfuncion del intestino medio, los tubulos de malpigio y los hemocitos. Tambien ejercen una accion inmunodepresora en el insecto e inhiben la encapsulacion multicelular del hongo (Aldana 1998; Tanada y Kaya 1993; Burges y Hussey 1971). Entre los mecanismos de invasion de los hongos entomopatogenos estan el de ingestion y el de contacto a traves de la cuticula, siendo el mas frecuente el de contacto. En este, el hongo penetra la cuticula del insecto y llega al hemocele, donde libera una serie de metabolitos toxicos que inhiben el sistema inmune del insecto, el cual finalmente muere debido a los daños mecanicos y fisiologicos sufridos (Boucias y Pendland 1988, 1991; Tanada y Kaya 1993; Aldana 1998). Por ingestion, el hongo ingresa por las partes bucales y el intestino, siendo estos canales alimentarios inhospitalarios a la penetracion del hongo debido a los efectos de anaerobiosis, enzimas digestivas, pH adverse velocidad del alimento y e! papel protector de la membrana peritr6fica (Dillon y Charnley 1986).
La ontogenia del hongo M.anisopliae se observo mediante estudios histologicos en obreros de Coptotermes curvignatthus Holmgr. en Malasia (Sajap y Kaur 1990). Igualmente se realizaron estudios de infeccion bucal y a traves del tracto digestivo por el hongo M. anisopliae de la langosta Schistocerca gregaria Forsk (Dillon y Charnley 1986).
El objetivo de este estudio fue evaluar el mecanismo de accion de M. anisopliae sobre los soldados y obreros de Nasutitermes sp. mediante la tecnica de aclaramiento con hidroxido de potasio.
Materiales y Metodos
Area de estudio
Se realizaron cuatro colecciones de termitas en la quebrada la Volcana (Loma de los Balsos) sector del Poblado al sureste de Medellin, Antioquia (Colombia) caracterizada por presentar una zona de vida clasificada como bosque humedo premontano (bh-P) con temperatura promedio de 20°C, precipitacion media anual de 1.000-2.000 mm y una faja altitudinal de 1.450 msnm (Espinal 1990).
Coleccion y mantenimiento de las termitas
Las termitas Nasutitermessp. se colectaron en un tronco de un arbol que pertenece al genero Cordia (Familia: Boraginaceae). Se capturaron termitas de diferentes castas (soldados y obreros) realizando incisiones en el termitero y/o colectando trozos de madera del arbol. Las temitas se introdujeron, utilizando pincel, en recipientes plasticos o de vidrio a los cuales previamente se les adicionaron los sustratos provenientes del termitero y se cubrieron con vinilpel durante el transporte hacia el laboratorio.
Para la reproduccion en el laboratorio se utilizaron recipientes de fibra de vidrio moldeado de 20 x 25 cm de diametro; a los recipientes se les agrego una capa de grava gruesa 2 cm de espesor, seguida de una capa de grava fina del mismo espesor, finalmente una capa de 10 cm de tierra del mismo termitero con arena fina (25% en volumen) (Norme Francaise 1990). Estos recipientes se cubrieron con muselina; los bordes del recipiente se impregnaron con vaselina para evitar el escape de las termitas.
Las colonias se mantuvieron en oscuridad, con circulacion de aire y a una temperatura de 26°C ± 2°C y 75% de humedad relativa. La humedad de la tierra, la temperatura y la humedad relativa dentro del termitero se controlaron diariamente (norme Francaise 1990).
Cultivo del hongo
La cepa del hongo M. anisopliae (Ma 45), aislada de un coleoptero (de una subfamilia de la familia Melolonthidae) fue donada por CORPOICA (Corporacion Colombiana de investigacion Agropecuaria). El hongo se cultivo en tubos y cajas de Petri con medio Agar Sabouraud Dextrosa (SDA) (Oxoid LTD., Basingstoke, Hampshire, England) a temperatura ambiente (21-26°C), durante 10-12 dias y se aimaceno a 4°C. La cepa se replico cada quince dias en SDA y se observaron los conidios y el micelio al microscopio (Aldana 1998; Zoberi y Grace 1990).
Pruebas de patogenicidad
Para las pruebas de patogenicidad se utilizaron mas termitas soldados que obreros por ser la casta mas numerosa dentro de la colonia estudiada. A partir de una solucion madre de conidios obtenida de los cultivos del hongo se impregnaron durante 5 min, 10 lonas y 10 papeles de filtro circulares de 100 x 15 mm con una concentracion de 1 x 107 conidios/ml. Las lonas y los papeles de filtro impregnados se introdujeron en frascos medianos que contenian sustrato proveniente del termitero y se adicionaron 5 termitas soldados (de 4 mm) y un obrero (de 3 mm de longitud) y se cubrieron con muselina. La supervivencia de las termitas se verifico diariamente durante ocho dias. Como testigos se utilizaron lona y papel de filtro impregnados con agua destilada esteril. Estos ensayos se realizaron por triplicado.
Durante los ensayos se controlaron diariamente las condiciones de oscuridad, temperatura y humedad relativa y se verifico la actividad de las termitas.
En total, en los ensayos, se utilizaron 150 termitas soldados con el inoculo impregnado en el papel de filtro, 150 con el inoculo impregnado en lonas y 100 en cada testigo. Se trataron 30 obreros en cada inoculo y 10 en cada testigo.
Para observar la penetracion del hongo en el insecto, se realizo la tecnica de aclaramiento de la termita descrita por Stehr (1987), utilizando soluciones de KOH al 5, 10, 15 y 20% a diferentes tiempos (24, 48 y 72 h) de exposicion. Luego, los insectos se sumergieron en una solucion agua: alcohol (l:3v/v) y despues en fenol puro. Posteriormente, se realizaron preparaciones del insecto en portaobjetos que se tineron con azul de lactofenol para observar hifas y esporas en las partes internas del cuerpo de la termita.
Micosis
Para evaluar el desarrollo del hongo y la presencia de daños en las termitas se colocaron los soldados muertos y obreros, en cajas de Petri esteriles, con papel de filtro Whatman N° 1 humedo, esteril y se observo la aparicion de micelio durante ocho dias.
Analisis estadistico
La mortalidad para los cuatro tratamientos (hongo en papel de filtro y en lona y agua destilada en papel de filtro y en lona) se comparo en cada dia utilizando la prueba de homogeneidad de proporciones, la cual se realizo en el paquete estadistico Statxact 4.0.1 (Cytel Software Corporation, Cambridge, Ma).
Resultados y Discusion
El porcentaje de mortalidad de los soldados fue diferente tanto entre los tratamientos como entre los dias. En efecto, la mortalidad en el primer dia fue mayor cuando se uso agua destilada en papel de filtro (46%). En el segundo dia la mortalidad mayor (82%) se presento empleando el papel de filtro impregnado con el Meterhizium anisopliae mientras que la lona con el hongo resulto eficaz solo al tercer dia (Tabla 1). En la Tabla 2 se presentan los porcentajes de mortalidad para los obreros. no se encontraron diferencias entre tratamientos en ninguno de los tres dias. Se observo tambien que el porcentaje mayor de mortalidad ocurrio en el segundo dia para todos los tratamientos. La comparacion entre tratamientos se realizo para cada dia, tanto para los soldados como para los obreros, segun la metodologia de Snedecor y Cochran 1978.
Las termitas obreros y soldados presentaron mortalidad al segundo dia debido a la accion del hongo M. anisopliae, que se observo mediante la tecnica de aclaracion en KOH (10 y 15%) dentro el cuerpo de estos, resultados que concuerdan con los estudios realizados por Sajap y kajur (1990); Milner et al. (1998) y Delate et al. (1995). Igualmente, los testigos presentaron mortalidad debido a factores como: la fragilidad natural (no permite ser sometidos a tecnicas de asepsia en el laboratorio antes de los bioensayos) y a contaminantes naturales como Aspergillus flavus y Serratia marcescens (entomopatogenos) que habitan en su mismo nicho ecologico (Steinhaus 1949; Tanada y Kaya 1993; Martius 1998). Asi mismo, los testigos se sometieron a la tecnica de aclaracion en KOH (10 y 15%) y no mostraron invasion en sus tejidos por el hongo M. anisopliae.
En cuanto a la forma de invasion del hongo M. anisopliaeen las termitas, a las 24 h post-inoculo los conidios se observaron sobre la cuticula, superpuestos y estrechamente unidos a las setas y poros de las setas (Fig. 1), en las patas, en el femur y tibia; tibia y tarsomeros, unas tarsales y tergitos abdominales. Luego, estos formaron tubos germinales continuando su crecimiento. Esto concuerda con los estudios realizados por Boucias y Pendland (1988) con M.anisopliae y B. bassiana donde se demostro que la topografia y propiedades quimicas de la epicuticula facilitan la adhesion conidial y la orientacion del tubo germinal, encontrando que los conidios de estos hongos tienen preferencia por toda la region del cuerpo del insecto y la superficie de la cuticula que contiene espinas cuticulares cortas.
Ademas, la forma de invasion, iniciando en la cuticula, concuerda con los resultados descritos por Sajap y Kaur (1990) y Guarles y Bucks (1995) quienes realizaron ensayos en termitas Coptotermes curvignathus donde los conidios de M. anisopliae germinaron sobre la cuticula y el tubo germinal penetro dentro de la cuticula a las 24 h post-inoculacion y luego en el exoesqueleto.
La interaccion conidio-cuticula (union de los carbohidratos del hongo M. anisopliae y las proteinas de la cuticula del insecto) no ha sido totalmente esclarecida (Latge et al. 1988).
Los tubos germinales formados por micopatogenos en artropodos producen celulas apresoriales que secretan sustancias mucilaginosas y exoenzimas que ayudan a la penetracion del hongo a traves de la cuticula del insecto (Zacharuk (1970b); Schabel (1978); Travland (1979); Fargues (1984); Charnley (1984); Fersson et al. 1984; St. Leger et al. 1986; Murrin y Nolan (1987); St. Legier et al. (1989a y b); sin embargo, dicha estructura no se observo en este estudio.
McCauley et al. (1968) y Zacharuck (1970a) observaron que los conidios de M.anisopliae se pueden unir a toda la superficie cuticular, remover facilmente y permanecer unidos a la epicuticula asociada con pliegues protegidos. Solo algunos micopatogenos terrestres reconocen especificamente la cuticula hospedante. Ademas, se encontro que los conidios de un linaje de M.anisopliae aislados del Scarabaeido (Scarab) Cetonia aurata se unen facilmente a la cuticula de C. aurata, pero no a las larvas del Scarab Oryctes rhinoceros (Fargues 1984). En este estudio se observo claramente que los conidios de M.anisopliae se unen a la cuticula de Nasutitermes sp. (Fig. 1).
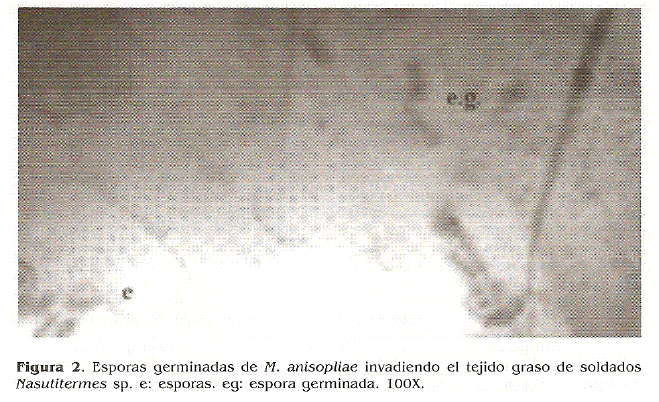
Una vez que los conidios llegan al interior de la cavidad del cuerpo, el hongo crece y mata el insecto. Aunque se ha observado que a las 48 h post-inoculacion los cuerpos hifales son abundantes en el hemocele de las termitas, en las placas aclaradas con KOH y tenidas con azul de lactofenol, no se encontraron hifas en el hemocele debido a que la tecnica de aclaramiento con KOH no permitio observar la invasion del hongo en el hemocele y a las 72 h de la infeccion se detectaron hifas invadiendo principalmente el tejido graso (Fig. 2) e intestino (Fig. 3), continuando hacia los tejidos musculares, nervioso y aparato digestivo mostrando una invasion fungica en todos los organos de la termita Nasutitermes sp. Observaciones que concuerdan con los resultados obtenidos por Sajap y Kaur (1990) y Tanada y Kaya (1993) en las termitas C. curvignathus, donde el hongo M.anisopliae invadio extensivamente el tejido graso, los tejidos musculares e intestino en el periodo comprendido entre 60-72 h despues de la infeccion.
Aunque las termitas Nasutitermes sp. murieron entre 1-3 dias despues de la inoculacion del hongo entomopatogeno, no se observo el micelio emergiendo de la cuticula del insecto hasta 96 h post-inoculacion (Fig. 4a), lo que concuerda con los resultados obtenidos por Sajap y Kaur (1990) en C. curvignathus en los que la emergencia del micelio se presento a las 100 h post infeccion.
En este estudio se logro la infeccion de soldados termitas adultos que tienen cuerpos esclerotizados (Fig. 4b), los cuales ofrecen cierta resistencia hacia la accion fungica ya que la cuticula melanizada es relativamente resistente al ataque enzimatico por hongos (Charnley y St. Leger 1991). Los hongos usualmente causan mortalidades por uno de los siguientes aspectos: deficiencia nutricional, invasion y destruccion de tejidos (que puede ser antes o despues de la muerte del insecto) y liberacion de toxinas (Tanada y Kaya 1993).
La micosis se evaluo durante ocho dias despues de la inoculacion del hongo M.anisopliae. El micelio en el cuerpo de la termita tomo coloraciones desde blanca hasta verde oliva claro, verde oliva oscuro con tintes grisaceos hasta verde oscuro, con un aspecto algodonoso al inicio del crecimiento hasta polvoso en la fase final. A las 96 h se observo micelio en forma de filamentos hialinos sobre la parte ventral del torax y sobre las patas. A las 120 h las termitas presentaron un micelio blanco en forma de hilos transparentes ocupando todo el cuerpo de la termita. El hongo se encontro invadiendo la superficie ventral de la cabeza (gulamenton, prementon, ligula, palpos labiales; maxilas: galea, lacinia, palpos maxilares y mandibulas). El micelio se observo de una coloracion blanca con un aspecto algodonoso en la cabeza (gena y postgena), en forma de un escudo en la cervix, en la parte posterior del protorax, dorso-pleura, prosternum, mesosternum y metasternum, y en las articulaciones de las patas anteriores, medias y posteriores (articulaciones entre femur y tibia, tibia y tarsomeros y unas tarsales). El hongo no se encontro invadiendo la termita dorsalmente; solamente se observo micelio blanco en uno de los tergitos abdominales posteriores en la membrana intersegmental.
A las 144 h de la inoculacion, el micelio blanco se torno de color verde ocupando las mismas partes que se mencionaron anteriormente y el nasuto empezo a cubrirse de micelio en su parte apical. A las 168 h, la superficie dorsal del cuerpo de la termita se cubrio totalmente de un micelio hialino y en los tergitos abdominales posteriores se encontro micelio blanco. El micelio verde empezo a tomar una coloracion verde oliva que prolifero en partes bucales, superficies dorso y ventro-laterales del torax y en las articulaciones de las patas. A las 192 h el micelio verde se tomo mas oscuro pero de consistencia polvosa e inicio la invasion de la cabeza (piezas bucales, genas, postgenas), prosternum, mesosternum, metasternum y patas. El hongo lleno los tergitos toracicos y comenzo a cubrir los tergitos abdominales posteriores. Finalmente, la termita quedo invadida totalmente por el hongo.
Conclusiones
- El mecanismo de invasion del hongo M. anisopliae en la termita Nasutitermes sp. es via integumentaria, iniciandose con la adherencia de la espora a la cuticula, la penetracion al intestino y al tejido graso conllevando a la muerte del insecto y su posterior esporulacion.
Agradecimientos
Los autores expresan los mas sinceros agradecimientos al profesor Jaime Calle por la revision y correccion del manuscrito, a los profesores Constantino Mantilla, Sandra Uribe S., Fabio Pineda G., Jose Rincon, Luis Carlos Mejia, Abel Diaz C. y a los companeros del laboratorio de Micologia y Microbiologia del Instituto de Biologia y del Posgrado de Entomologia de la Universidad Nacional Sede Medellin que contribuyeron a la ejecucion de este trabajo. Tambien al Comite para el Desarrollo de la Investigacion (CODI), al Instituto de Biologia Universidad de Antioquia y a la Universidad Nacional de Colombia Sede Medellin por la financiacion y apoyo a este trabajo.
Literatura citada
ALDANA, R. 1998. Evaluacion de polimorfismo genetico y de la produccion de toxinas en cuatro aislados del hongo entomopatogeno Metarhizium anisopliae (Metsch.) Sor. Posgrado Biologia. Facultad de Ciencias Exactas y Naturales. Departamento de Biologia. Universidad de Antioquia. 44 p. [ Links ]
ALVES, S. B.; ALMEIDA, J. E. M.; MOINO, A.; STIMATIC, J. L.; PEREIRA, R. M. 1995. Uso de Metarhiziuma nisopliae e Beauveria bassiana no controle de Cornitermes cumulans em pastagens . Ecossistema Faculadade de Agronomia "Manoel Carlos Goncalves". Brasil. 20: 50-57. [ Links ]
BARNETT, H. L. 1967. Ilustrated genera of imperfect fungi. Burguess Publishing Company, Minneapolis (U.S.A.). 225 p. [ Links ]
BERON W. C. 1983. Daños y control de termitas. Seminario como trabajo de investigacion. Facultad de Ciencias. Universidad nacional de Colombia. Medellin. 68 p. [ Links ]
BORROR, D. J.; TRIPLEHORN, C. A.; JOHNSON, N. F. 1989. An introduction to the study of insects, p. 234-241. Saunders College Publishing. 6 ed. Fort Worth, United States of America. 875 p. BOUCIAS, D. G.; PENDLAND, J. C. 1988. Non specific factors involved in the attachment of entomopathogenic Deuteromycetes to host insect cuticle. Applied and Environmental Microbiology 54 (7): 1795-1805. [ Links ] [ Links ]
BOUCIAS, D. G.; PENDLAND, J. C. 1991. Attachment of mycopathogens to cuticle: The initial event of mycoses in arthropods hosts, p 101-127. En: G. T. Cole and Harvey C. Hoch (eds.). The fungal spore and disease initiation in plants and animals. Plenum Press. New York - London. 555 p. [ Links ]
BURGES, rt. D.; HUSSEY, N. W. 1971. Microbial control of insects and mites. Academic Press. Great Britain, p. 398-401. 861 p. [ Links ]
CHAI, Y. G. 1995. Preliminary studies on the pathogenicity of some enthomogenous fungi to Coptotermes formosanus. Chinese Journal of Biological Control 11 (2): 68-69. [ Links ]
CHARNLEY A. K. 1984. Physiological aspects of pathogenesis in insects by fungi: A speculative review. En: Anderson J. M., Rayner A. D. M., Walton D. (eds.). Invertebrate: Microbial Interations, British Mycological Symposium. University Press, Cambridge. 6: 229-271. [ Links ]
CHARNLEY A. K.; ST. LEGER R. J. 1991. The role of cuticle-degrading enzymes in fungal pathogenesis in insects, p. 267-283. En: G. T. Cole and riarvey C. floch (eds.). The fungal spore and disease initiation in plants and animals. Plenum Press. New York-London. [ Links ]
COPPEL, H. C; LIANG, M. C. 1987. Rhabditoid nematodes associated with subterranean termites. Forestry Research Notes. USA. N° 274, 5 p. [ Links ]
DELATE, K. M.; GRACE, J. K.; TOME C. H. M. 1995. Potential use of pathogenic fungi in baits to control the Formosan subterranean termite (Isoptera: RhinoTermitidae). Journal of Applied Entomology 119 (6): 429-433. [ Links ]
DILLON R. J.; CHARNLEY, A. K. 1986. Invasion of the pathogenic fungus Metarhiziumanisopliae through the guts of germfree desert locust, Schistocerca gregaria. Mycopathologia 96: 59-66. [ Links ]
DILLON, R. J.; CHARNLEY, A. K. 1991. The fate of fungal spores in the insects gut. p. 129-156. En: G. T. Cole and Harvey C. Hoch (eds.). The fungal spore and disease initiation in plants and animals. Plenum Press. New York- London. 555 p. [ Links ]
DOMSCH, K. H.; GAM, W. AND. ANDERSON, T-H. 1980. Compendium of soil fungi. Academic Press. Vol 1. 859 p. [ Links ]
ESPERO, L. V. 1984. Survey of entomogenous fungi associated with termites (Philippines). Scientific Journal 6(1): 23-24. [ Links ]
ESPINAL, L. S. 1990. Zonas de Vida de Colombia. Universidad Nacional de Colombia Seccional Medellin. Facultad de Ciencias. Departamento de Ciencias p. 61-62. 121 p. [ Links ]
FARGUES, J. 1984. Adhesion of the fungal spore to the insect cuticle in relation to pathogenicity. p. 90-110. En: Infection Processes of fungi (D. W. Roberts and J. R. Aist, (ed.). Rockefeller Foundation Conference Report. [ Links ]
GAO, D. R.; ZHU, B. Z.; GAN, B. R.; HE., S. S.; YUAN, S. X. 1985. A new toxic bait for the control of forestinfecting termites. Journal of the Nanjing Institute of Forestry. China 5: 128-131. [ Links ]
GARCES, J. E. 1997. Generalidades sobre las principals plagas insectiles en plantaciones forestales de Colombia (Guia de campo). Trabajo Final de Especializacion. Universidad Nacional de Colombia. Facultad de Ciencias y Ciencias Agropecuarias. Posgrado en Entomologia. Medellin, p. 293- 311. [ Links ]
GAVINO, A. A. 1984. Efficacy of Bacillus thuringiensis Berliner (Backie, biochem product) as biological insecticide against subterranean termites Philippines. Scientific Journal 6 (1): 25. [ Links ]
HARRIS, W. V. 1971. Termites. Their recognition and control. Longman. Second ed. London,Great Britain. 186 p. [ Links ]
HASSAN, F. A. 1990. Important insects pests of Casuarina in Egypt, p. 102-109. En: Advances in casuarina research and utilization. Proceedings of the Second International Casuarina Workshop. Cairo, Egipto. [ Links ]
HUMBER, R. A. 1997. Fungi Identification, p. 153-185. En: L. Lacey (ed.). Manual of techniques in insect pathology. USDA, ARS, Academic Press (Great Britain). [ Links ]
KIRK, R M.; CANNON, P F.; DAVID, J. C AND J. A. STALPERS. 2001. Ainsworth & Bisby 's Dictionary of the fungi. Edited Cabi Publishing. 624 p. [ Links ]
LATGE, J. R; MONSIGNY, M.; PROVOST, M. C. 1988. Visualization of exocellular lectins in the entomopathogenic fungus Conidio-bolus obscurus. Journal of Histochemistry and Cytochemistry 36: 1419-1424. [ Links ]
L[J]UTIKOVA, L. I. 1990. Entomogenous fungi as control agents for termite Anacanthotermes ahngerianus Jacobs (lsoptera, HodoTermitidae). p. 634-635. En: G.K. Veeresh; B. Mallick; C.A. Viraktamath (eds.). Social insects and the environment. [ Links ]
LOGAN, J. W. M.; COWIE, R. H.; WOOD, T. G. 1990. Termite (lsoptera) control in agriculture and forestry by nonchemical methods: a review. Bulletin of Entomological Research 80 (3): 309-330. [ Links ]
McCAULEY, v. J. E.; Zacharuk, R.Y; TINLINE, R.D. 1968. Histopathoiogy of green muscardine in larvae of four species of Elateridae (Coleoptera). Journal of Invertebrate Pathology 12: 444-459. [ Links ]
MARTIUS, C. 1994. Diversity and ecology of termites in Amazonian forest. Pedobiologia 3 (5): 407-428. [ Links ]
MARTIUS, C. 1998. Perspectivas do controle bioiogico de cupins (Insecta: lsoptera). Revista Brasilera de Entomologia 41: (2-4): 179-194. [ Links ]
MILNER, R. J.; STAPLES, J. A. 1996. Biological control of termites: results and experiences within a CSIRO project in Australia. Biocontrol Science and Technology 6 (1): 3-9. [ Links ]
MILNER, R. J.; STAPLES, J. A.; LUTTON, G. G. 1998. The selection of an isolate of the Hyphomycete fungus, Metarhizium anisopliae, for control of termites in Australia. Biological control: theory and applications in pest management 11 (3): 240-247. [ Links ]
MIX, J.; BEAL, R. H. 1985. Can nematodes halt subterranean termites?. Pest Control 53 (2): 22-23. [ Links ]
MURRIN, F.; NOLAN, R. A. 1987. Ultrastructure of the infection of spruce budworm larvae by the fungus Entomophaga aulicae. Canadian Journal of Botany 65: 1694-1706. [ Links ]
MYLES, T. G.; STRACK, B. H.; FORSCHLER, B. 1998. Distribution and abundance of Antennopsis gallica (Hyphomycete: Gleohaustoriales), an ectoparasitic fungus, on the eastern subterranean termite in Canada. Journal of Invertebrate Pathology. 72 (2): 132-137. [ Links ]
NORME FRANCAISE N. F. 1990. Determination de I' efficacite preventive contre Reticulitermes santonensis de Feytaud (Methode de laboratoire). European Standard Comite Europeen de Normalisation. 15 p. [ Links ]
OCHIEL, G. S.; EVANS, H. C; EILENBERG, J. 1997. Cordycepioideus, a pathogen of termites in Kenya. Mycologist 11 (1): 7-9. [ Links ]
PEARCE, M. J. 1999. Termites and Pest management. Cab International. [ Links ]
Printed and bound in the UK at the University Press, Cambridge p. 127-137. [ Links ]
PERSSON, M.; HALL, L.; SODERHALL, K. 1984. Comparison of peptidase activity in some fungi pathogenic to arthropods. Journal of Invertebrate Pathology 44: 342-348. [ Links ]
GUARLES, W; BUCKS, C. 1995. Non toxic termite treatments (include related articles). Organic Gardening. Dec: 44-50. [ Links ]
SAJAR A. S.; KAUR, K. 1990. Histopathoiogy of Metarhizium anisopliae, an entomopathogenic fungus infection in the termite, Coptotermes curvignathus. Pertanika. Malaysia 13 (3): 331-334. [ Links ]
SAMSON, R. A. 1981. Identification: Entomopathogenic Deuteromycetes. p. 93-105. En: H. D. Burges (ed.), Microbial control of pests and plant diseases. 1970-1980. Academic Press Inc. Ltd. London (Great Britain). [ Links ]
SCHABEL, H. G. 1978. Perccutaneous infection of Hylobius paales by Metarhizium anisopliae. Journal of Invertebrate Pathology 31: 180-187. [ Links ]
SNEDECOR, G. W.; COCHRAN, W. G. 1978. Metodos estadisticos. Cecsa. Mexico, p. 297-302. [ Links ]
St. LEGER, R. J.; BUTT. T. M.; GOETTEL, M.S.; STAPLE, R. C; ROBERTS, D. W. 1986. Cuticle-degrading enzymes of entomopathogenic fungi: Mechanisms of interaction between pathogen enzymes and insect cuticle. Journal of Invertebrate Pathology 47: 295-302. [ Links ]
St. LEGER, R. J.; BUTT. T- M.; GOETTEL, M.S.; STAPLE, R. C; ROBERTS, D, W. 1989a. Production in vitro of appressoria by the entomopathogenic fungus Metarhizium anisopliae. Experimental Mycology 13: 274- 288. [ Links ]
St. LEGER, R. J.; BUTT. T. M.; GOETTEL, M.S.; STAPLE, R. C; ROBERTS, D. W. 1989b. Synthesis of proteins including a cuticle degrading protease during differentiation of the entomopathogenic fungi Metarhizium anisopliae. Experimental Mycology 13: 253-262. [ Links ]
STEHR, F. W. 1987. Techniques for collecting, rearing, preserving and studying immature insects, p. 7-18. En: Frederick W. Stehr (ed.). Immature Insects. Kendall/ Hunt Publishing Company. Dubuque Iowa, U.S.A. [ Links ]
STEINHAUS, E. A. 1949. Fungous Infections (Mycoses). Principles of Insect Pathology. McGraw-Hill Book Company, Inc. United States of America. 757 p. [ Links ]
TANADA, Y.; KAYA, H. K. 1993. Insect pathology. Academic Press, Inc. Harcourt Brace Javanovich. Printed Unites States of America, p. 359-361. [ Links ]
TRAVLAND, L. B. 1979. Initiation of infection of mosquito larvae (Culisetainornata) by' Coelomyces psorophorae. Journal of Invertebrate Pathology 33: 95-105. [ Links ]
VARON, R T.; MORALES, S. L. 1997. Arboles de Aburra. Area Metropolitana del Valle de Aburra. Editorial Colina. Medellin. p 137. [ Links ]
WATSON, J. A. L.; GAY, F. J. 1991. lsoptera (Termites), p. 330-347. En: The Insects of Australia, (ed.) Csiro. Vol II. Cornell University Press. Ithaca, New York. [ Links ]
WELLS, J. D.; FUXA, J. R.; HENDERSON, G. 1995. Virulence of four fungal pathogens to Coptotermes formosanus (lsoptera: RhinoTermitidae). Journal of Entomological Science 30 (2): 208-215. [ Links ]
YENDOL, W. G.; PASCHKE, J. D. 1965. Pathology of an Entomophthora infection in the eastern subterranean termite Reticuli-termes flauipes (Kollar). Journal of Invertebrate Pathology 7: 414-422. [ Links ]
ZACHARUK, R. Y. 1970a. Fine structure of the fungus Metarhizium anisopliae infecting three species of larval Elateridae (Coleoptera). I. Dormant and germinating conidia. Journal of Invertebrate Pathology 15:63-80. [ Links ]
ZACHARUK, R. Y. 1970b. Fine structure of the fungus Metarhizium anisopliae infecting three species of larval Elateridae (Coleoptera). II. Conidial germ tubes and appressoria. Journal of Invertebrate Pathology 15: 81-911. [ Links ]
ZOBERI, M. H. 1995. Metarhizium anisopliae, a fungal pathogen of Reticulitermes flauipes (lsoptera: RhinoTermitidae). Mycologia 87 (3): 354-359. [ Links ]
ZOBERI, M. H.; GRACE, J. K. 1990. Isolation of the pathogen Beauveria bassiana from Reticulitermes flauipes (lsoptera: RhinoTermitidae). Sociobiology 16 (3): 289-296. [ Links ]
ZIMMERMANN, G. 1993. The entomopathogenic Fungus Metarhizium anisopliae and its potential as a biocontrol agent. Pesticide Science Great Britain. 37: 375-379. [ Links ]
ZULUAGA, M. P. 2002. Susceptibilidad de Microcerotermes sp. (lsoptera: Termitidae) a Metarhizium anisopliae (Metsch) Sor. en plantaciones de Eucalyptus tereticornis. Tesis presentada para obtener el titulo de Biologa. Universidad de Antioquia. Facultad de Ciencias Exactas y Naturales. Instituto de Biologia. 56 p. [ Links ]
Recibido: Nov. 13/2002 Aceptado: Nov. 29/2003


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}