










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488On-line version ISSN 2665-4385
Rev. Colomb. Entomol. vol.31 no.1 Bogotá Jan./June 2005
Observaciones acerca de la biología y los enemigos naturales de saccharicoccus sacchari y Pulvinaria pos elongata, dos homópteros asociados con la hormiga loca en caña de azúcar
Observations on the biology and natural enemies of Saccahricoccus sacchari and Puluinarza pos elongata two homopterans associatecl with the crazy ant in sugar cane
KATHERINEGiRON1 ,LUZ ADRIANA LASTRA2 , LUIS ANTONIO GOMEZ L3, NORA CRISTINA MESA C.4
1 Tesista Facultad de Ciencias Agropecuarias Universidad Nacional de Colombia sede Palmira E- mail: katherinegiron77@hotmail com
2 Investigadora de Cenicana Biologa Entomologa
3 Autor para correspondencia: Ingeniero Agrónomo. Ph. O. Entomólogo, Cenicaña. A. A. 9138 Cali. E-mail: lagomez@cenicaña.org
4 Profesora Asociada. Universidad Nacional de Colombia, sede Palmira. Facultad de Ciencias Agropecuarias. E-mail: ncmcobo@carpa.ciagri.usp.br
Resumen. La hormiga loca es un insecto que adquiere importancia económica agrícola cuando establece relaciones simbioticas con insectos chupadores fitofagos En cana de azucar se asocia con dos de ellos Saccliaricoccus sacchari y Pulvinaria pos elongata Cuando los niveles de infestacion son extre mos Pulinnaria en asociacion con la hormiga puede llegar a causar perdidas severas tanto en la concen tracion de azucar como en el tonelaje de la cana Con el objeto de conocer el ciclo de vida de ambas especies se establecieron ensayos bajo condiciones de laboratorio e invernadero y se observo la inciden cia de enemigos naturales en campo S .sacchari tuvo un ciclo de vida promedio de 54 8 dias pasando por dos instares y el estado adulto el numero de crias promedio por hembra es de 219 son gregarios y se ubican en los entrenudos del tallo Dos especies de la familia Encyrtidae aun sin identificar parasitan a S saccharz con un porcentaje de parasitismo del 47% Una especie de Dzadoplosis (Cecidomyiidae) se alimenta de los huevos de las hembras maduras, además un hongo identificado como Aspergillus pos pai asiticus afecto el 53 2% de los individuos en campo Puluinaria sp tuvo un ciclo de vida promedio de 77 días y pasa por dos ínstares y el adulto. EJ número de crías por hembra en promedio fue de 179, se ubican en el enves de las hojas y son poco moviles En invernadero se encontro a Diadoplosis coccidivora (Cecidomyiidae) alimentándose de la progenie de las hembras de ,Puluinaria sp. con porcentajes hasta del 85% En campo se encontro una avispa (Encyrtidae) emergiendo de individuos de segundo instar de esta ultima especie.
Palabras clave: Relaciones simbioticas. Ciclo de vida.
Summary. The crazy ant is an insect that becomes economically important to agriculture when it establishes symbiotic relationships with sap sucking insects. In sugar cane, it is associated with two of them: Sacctiaricoccus sacchan and Pulvinaria pos elongata When infestation levels are very high Pulvinaria in association with the ant can induce severe losses in both sugar concentration and tonnage of cane loorder to understand the life cycle of each species. experiments were set up under laboratory and green house conditions and the incidence of natural enemies was determined in the field S saccharz had a mean life cycle of 548 days, passing through two instars and the adult stage; mean progeny per female was 219 they are gregarious and concentrate on the stem internodes Two species of unidentified Encyrtidae parasitized S. sacchari, with a parasitism rate of 47%. A species of Diadoplosis (Cecidomyiidae) eats eggs of the mature females and a fungus identified as Aspergillus pos parasiticus affected 53 2% of individuals in the fleld, Pulvinaria sp. had a mean lite cycle of 77 days and passed through two instars and tlie adult. Mean progeny per female was 179; they are located on the underside of the leaf and are not very mobile ln the greenhouse Dzadoplosis coccidwora (Cecidomyiidae) fed on eggs of Pulvinaria sp females at rates up t 85%. ln the field, a wasp (Encyrtidae) was found emerging from second instar individuals of Putvinaria sp.
Key words: Symbiotic relationships. Life cycle.
Introduccion
Muchos insectos del orden llomoptera se simbioticamente con hormigas ocasionando, en la mayoria de los casos, impactos severos sobre el cultivo hospedero Este es el caso de la asociacion que se ha establecido en caña de azúcar entre saccharicoccus sacchari (Cockerell) (Pseudococcidae) y Pulvznaria pos elongata (Coccidae) con la hormiga loca Paratrechina fulva Mayr (llymenoptera Formi cidae), cuya presencia en el país ha causado daños tanto de tipo social, como ecologico y economico Un ejemplo de lo anterior son los registros existentes de su ataque directo al hombre, aves, mamíferos reptiles y otros insectos (Zenner 1985).
Segun llolldobler y Wilson (1990) la rela cion entre las hormigas y algunos homop teros se debe a que estos ultimos secretan liquidos azucarados importantes en la dieta alimenticia de las hormigas que, en retribución, los protegen de condiciones adrelacionan versas y ayudan a su diseminacion.
S. sacchari es un insecto cosmopolita. En Colombia ha sido registrado en la mayoria de zonas en donde se cultiva caña de azúcar encontrandose asociado con la hormi ga loca El dano en el cultivo ocurre cuando el insecto inyecta su estilete en el tallo de las plantas y extrae los azúcares, desdoblando la sacarosa del jugo. reduciendo el briz e incrementando los azúcares reduc[ores, por lo cual no se produce panela sino miel de mala calidad (Nieves 1999). Además, Autrey et al. (1995) y Avellaneda et al. (2001), registraron a este insecto como transmisor del Virus Baciliforme de la Caña (ScVB).
Existe información acerca del ciclo de vida de este insecto bajo condiciones de laboratorio en Australia, y se sabe que el modo principal de reproducción es partenogénico, aunque en algunos casos, debido a la presencia de machos, puede ser sexual. Su fecundidad es alta y una hembra adulta puede colocar más de 1.000 huevos (beardsley 1962, cit por lnkermari et al. 1986). En Colombia. hasta el momento, no se han realizado estudios de este tipo.
En el caso de Pulvinaría pos elongata, que de aquí en adelante se denominará Pulvinaria sp. considerando que falta una identificación más precisa. no existe mucha información acerca de su biología. Sólo Harnrnon y Williams (1984) suministran datos de su desarrollo; según ellos, las henibras son ovovivíparas y pueden reproducirse sexual o partenogénicamente. El ciclo de vida de esta especie tampoco se ha estudiado en Colombia hasta el momento. El daño ocurre cuando el insecto, al inyectar su estilete en los vasos vasculares del floema. extrae el líquido intracelular ocasionando reducciones y/o retrasos en el crecimiento de los tallos. clorosis y necrosamiento en los folíolos finalizando con la muerte prematura de la planta en pocas semanas (Etienne 1977).
Dentro de las colonias de los homópteros existen factores que pueden regular las poblaciones. Uno de ellos es la incidencia de enemigos naturales como insectos y hongos entomopatógenos. En el caso de 5. sacchari, es controlado por individuos de la familia Coccinellidae (Cryptolaetnus Pulvinaria), de la familia Anthocoridae (Opoblates woodwardi) (Inkerman et al. 1986), de la familia Encyrtidae (Anaggrus saccharicola) y de la familia Drosophilidae (Cacoxenus perspicax), al igual que por hongos entomopatógenos como Aspergiltus parasiticus, Metarhizium anisoplae, Penicilliurn sp. y Cordyceps sp. (De barro 1990). En el caso de Pulvinaría sp. no hay mucha información acerca de los enemigos naturales y en el ámbito de la familia Coccidae, Hammon y Williams (1984) reconocen que éstos pertenecen a los órdenes Hymenoptera, Lepidoptera, Diptera y Neuroptera.
Debido a la falta de conocimiento de ambas especies en Colombia, el presente trabajo pretendió conocer aspectos biológicos de ambos homópteros. reconocer la presencia de enemigos naturales y determinar el efecto de transporte y diseminación por parte de las hormigas sobre ellos, como preámbulo de posteriores estudios que involucren el impacto económico y la determinación de las posibles pérdidas que puedan ocasionar los insectos al cultivo de la caña de azúcar.
Materiales y Métodos
Localización
El trabajo sé llevó a cabo en el Centro de Investigación de la caña de azúcar de Colombia CENICAÑA, ubicado en el corregimiento de San Antonio de los Caballeros, Municipio de Florida (Valle) a una altitud de 1.020 msnm, con una temperatura promedio de 24 ± 5°C y con una humedad relativa del 65 ± 5%.
La fase de campo se desarrolló en el ingenio Providencia, hacienda Piedechinche, ubicada en el municipio de El Cerrito, departamento del Valle del Cauca, a una temperatura promedio de 22,7 ± 0,6°C y a una humedad relativa promedio de 84 ± 3% yen el ingenio Central Castilla, hacienda Vista Hermosa, ubicada en el municipio de Corinto, departamento del Cauca con una temperatura promedio de 22,7 ± 0.5°C y una humedad relativa de 82 ± 4%.
Determinación del ciclo de vida de los homópteros y diferenciación morfológica entre ínstares
saccharicoccus sacchari
Para conocer el ciclo de vida de esta especie hubo necesidad de emplear una técnica que permitiera hacer las observaciones de los individuos sin maltratarlos, lo cual resulta muy difícil si éstas se hacen en su microhábitat natural; es decir, en los nudos de los tallos recubiertos por las vainas de las hojas. Por lo tanto, se desarrolló una metodología en condiciones de laboratorio, basada en el uso de fragmentos de caña sobre agar, que consistió en mantener fragmentos de tallos de 4 cm de longitud de cañas de 3 meses de edad, en una mezcla de agar noble (0,8 g agar/l00 ml de agua) y metilparaben (0,0003 g disueltos en alcohol al 50%) para contrarrestar la contaminación de hongos y ácido láctico al 50% (0,1 ml/l00 ml de agua) para contrarrestar la infección de bacterias, servida en cajas plásticas pequeñas con tapas acondicionadas con filtros. Los fragmentos se desinfectaron con Vitavax 300® (carboxin, 0,8 g/l de agua). sumergiéndolos en agua destilada por 24 h antes de ser depositados sobre el agar y se infestaron con 2 ninfas recién emergidas, sobre 25 trozos de tallos para un total de 50 individuos. Las condiciones en que se desarrollaron los individuos fueron de laboratorio con una temperatura promedio de 25°C y una humedad relativa de 64%.
En este ensayo se registró el momento de la muda, los cambios morfológicos que pueden presentarse y el número de días que tardan en llegar al estado adulto. Se evaluó la duración del adulto y la producción de huevos por hembra tanto de campo como de invernadero.
La diferenciación morfológica entre instares se constató mediante observaciones al microscopio y al estereoscopio de individuos de diferentes ínstares. Para facilitar la diferenciación de ciertas estructuras, algunos de los individuos coleccionados se aclararon con kOH al 10% por 20 min a una temperatura de 70°C, posteriormente se les extrajo el fluido interno y se tiñeron con fucsina ácida, luego se pasaron por alcohol al 95% para fijar la tinciór y finalmente se montaron sobre portaobjetos en bálsamo de Canadá. Los indiiduos que no se sometieron a este proceso se observaron directamente en el estereoscopio. Las características medidas fueror longitud del cuerpo, número de segmentos antenales y aparición de nuevas estructuras a medida que el insecto pasaba de un ínstar a otro.
Para determinar la duración del adulto y el número de huevos por hembra, se emplearon dos metodologías. La primera consistió en colocar individualmente 20 insectos de campo en el tercer ínstar sobre fragmentos de hojas de caña mantenidos en cajas de Petri plásticas, con algodones odontológicos humedecidos, a fin de conservar la humedad de tejido. La segunda se realizó con base en la metodología del agar, tomando 15 hembras en el tercer ínstar criadas en invernadero y colocando una hembra por tallo de caña. Para ambos casos se contabilizó el número de ninfas por hembra y con la segunda metodología además. se determinó la duración del adulto.
Puluinaria sp.
El ciclo de vida de esta especie se determinó empleando una metodología que consistió en infestar artificialmente 9 plantas de caña de 2,5 meses de edad sembradas en materas con 5 ninfas/planta recién emergidas del insecto, para un total de 45 individuos. Las plantas de aislaron en jaulas de tul ubicadas en invernadero. Se contabilizó el tiempo en que los individuos duran en cada instar el tiempo en que el insecto tarda en llegar al estado adulto. Las condiciones ambientales en que se desarrolló este ensayo fueron de invernadero, a una temperatura de 26°C y una humedad relatia de 80%.
Una segunda metodología para determinar la duración del adulto y el número de huevos por hembra se hizo a partir de 30 ninfas recién emergidas en el laboratorio y ubicadas individualmente sobre 30 plantas de caña sembradas en vasos de icopor. Después de que el insecto llegó al momento de la madurez. se iniciaron los registros sobre la longevidad y el número de huevos que coloco cada hembra.
La diferenciación morfológica entre los ínstares se realizó colocando a los individuos en glicerina, 9 de cada ínstar, para observarlos al microscopio y detectar los cambios morfológicos de sus estructuras en los diferentes ínstares.
Reconocimiento y evaluación de enemigos naturales sobre Saccltaricoccus saccharl y Pulvinaria sp.
Para identificar la presencia de enemigos naturales, se realizaron muestreos en la zona de Corinto (Cauca), Hacienda Vista Hermosa, y en la zona de El Cerrito (Valle), Haciendas Piedechinche y Providencia.
En el caso de S. sacchari, se recolectaron 45 hembras en total para la zona de Corinto (15 por lote) y 15 para la zona de El Cerrito (Hacienda Piedechinche). Cada una se introdujo en una caja plástica con tapa utilizada para análisis coprológico (3 cm de diámetro por 2 cm de profundidad) y se colocaron sobre una bandeja con espuma humedecida, mantenida en el laboratorio. Mientras duró el ensayo, de cada caja se contabilizaron los individuos que emergían de las hembras.
Para establecer la importancia de los entomopatógenos sobre S. sacctiari, se tomaron 30 tallos al azar de un lote experimental establecido en el ingenio Providencia, con una población de 65 individuos de diferentes ínstares, y se contabilizaron los cuerpos momificados y/o que manifestaran un desarrollo micelial verde y denso. Los individuos afectados se separaron y posteriormente se tomaron muestras de micelio para obtener un cultivo puro del hongo en medio PDA. mediante aislamientos sucesivos.
Para Putvinaria sp., se cortaron hojas de cañas de 9 meses de. edad que estuvieran infestadas con el insecto. Las hojas se llevaron al laboratorio y se introdujeron en dos unidades cilíndricas de PVC y acetato, a fin de registrar los parásitos después de su emergencia. De igual manera, en invernadero se realizó una evaluación en plantas de caña de 6-7 meses de edad, sembradas en materos plásticos y de los cuales se tomaron al azar cinco hojas de caña infestadas, en dos fechas distintas. De cada una de las hojas se recortaron 10 cm al azar para contabilizar el número total de hembras y el número de hembras parasitadas. Las hojas provenían de un ensayo de invernadero en donde se habían establecido plantas con y sin hormiga loca por lo que fue posible evaluar la incidencia de los parásitos en las condiciones mencionadas. Los datos se analizaron mediante el programa SAS, utilizando el diseño experimental completamente al azar con 2 tratamientos, 2 repeticiones. Los resultados se evaluaron mediante la prueba de comparación de Tukey (0,05%).
Resultados y Discusión
Determinación del ciclo de vida de los homópteros y diferenciación morfológica entre ínstares
Sacckaricoccus sacchari
La metodología de los fragmentos de talios de caña sobre agar mostró ser la adecuada por Cuanto no permitió que se secaran los trozos de caña mientras duró el desarrollo de los individuos además, de facilitar el control de microorganismos contaminantes.
El ciclo de vida completo de las hembras duró 54,8 días en promedio (‘tabla 1) y pasó posiblemente por dos instares ninfales y un estado adulto. Es de resaltar el hecho de que la literatura registra que el ciclo de vida de esta especie, bajo condiciones de laboratorio en el estado de California (USA). dura aproximadamente 30 días y que la hembra pasa por 3 ínstares antes de mudar al estado adulto (Mctçenzie 1967). IJichanco y Villanueva (1932) citados por Mckenzie (1967) registraron hasta 7 ínstares en ambos sexos. Posiblemente, las diferencias se atribuyan a las condiciones en las cuales se criaron los insectos, que no fueron muy semejantes a las que se dan en el ambiente natural, rodeadas de las yaguas de las hojas, además, de las condiciones alimenticias restringidas suministradas al insecto durante todo su ciclo de desarrollo. Los machos observados en los experimentos establecidos fueron muy escasos en las cotonias por lo que se asume que predoniinó el modo principal de reproducción partenogénico.
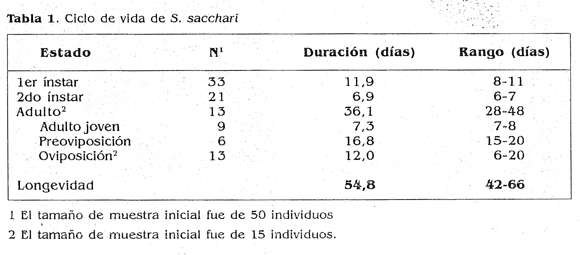
En cuanto a la duración del adulto y el número de huevos por hembra, se observó que, después de la segunda muda, el ciclo de los individuos se estancaba por cuanto no iniciaban el período de oviposidón. Parte de las hembras se aclararon y se vio que estaban llenas de huevos, lo que sugiere que las condiciones de cría a las que se sometieron los individuos no estimularon este proceso por estar en un espacio tan reducido y con una deficiencia nutricional evidente para las crías. Debido a esto, se trajeron colonias de campo y de invernadero y se estableció, basados en las características morfológicas junto con el aclarado de algunos individuos, que después de la segunda muda el insecto permanece en un estado en el cual no se notó ningún cambio de tipo reproductivo (Adulto joven). Sin una muda aparente, los adultos pasaron a un estado en el cual comenzaron a formar huevos (preoviposidón) y por último a la etapa de oviposidón. Las hembras no se alimentaron durante el período de gravidez, tal como lo menciona Beardsley (1960).
Las hembras traídas de campo colocaron en promedio 218,8 (± 36,8) huevos, con un rango de 181 a 273 huevos en un tiempo de 14 días, 20 (± 3,68) crías/día, mientras que las hembras provenientes del invernadero colocaron en promedio 82,76 (± 57,4) huevos con un rango de 6 a 166 en un periodo de 13 días, ambos casos sin la presencia de la hormiga loca. La longevidad del adulto en promedio fue de 12 (± 4,4) días en condiciones de laboratorio. Estas diferencias sedebieron posiblemente al lugar de procedencia del insecto, puesto que las hembras de campo se albergaban y alimentaban naturalmente en plantas de caña con mayor desarrollo de entrenudos, con mayor disposición de nutrientes y de espacio para la formación de raíces, lo cual las hace más vigorosas con respecto a las plantas desarrolladas en invernadero en materas individuales. Otros de los factores que pudieron afectar la producción de huevos entre ambas poblaciones, son las condiciones ambientales contrastantes (condiciones de campo y condiciones de invernadero) en las cuales el insecto se desarrolló.
Al comparar estos resultados con lo que registra la literatura se encuentran diferencias entre el número de huevos colocados por tina hembra de campo superando, en algunos casos, los 1.000 huevos, según Williams (1985). Nuevamente cabe la posibilidad de que algunos factores externos como temperatura, humedad, edad y variedad de la planta o aun la atención por parte de las hormigas (Beardsley 1960), influyeran en el número de huevos por hembra. Por lo tanto, el número reducido de descendencia, particularmente en el caso de las hembras recolectadas en plantas de invernadero, es un indicador de que la metodología no reflejó el potencial reproductivo de la especie.
Las hembras al ovipositar producen el ovisaco, una secreción filamentosa de aspecto algodonoso que envuelve los huevos. Minutos después de colocados los huevos, emergen las ninfas, y permanecen un tiempo cerca de la madre para luego desplazarse a buscar ubicación y alimento. En contraste, Williams (1985) observó diferencias en el período de incubación y eclosión que según él puede durar de 10 a 14 h. Este aspecto puede variar de acuer do con los factores externos que circun den al insecto.
Si bien todo pareciera indicar que los individuos presentes en el Valle del Cauca pertenecen a la especie saechari del género saccharicoccus, en realidad no se hizo una identificación formal del insecto, ni se envió a un especialista y queda la duda de que todas las diferencias que se hallaron para este piojo harinoso, en relación con lo registrado en la literatura, podrían expor el hecho de ser una especie diferente, pero faltaría por confirmar esta aseveración.
Los resultados obtenidos en las observa al microscopio y al estereoscopio, arrojaron las siguientes características que permitieron diferenciar los instares por los cuales pasan las hembras.
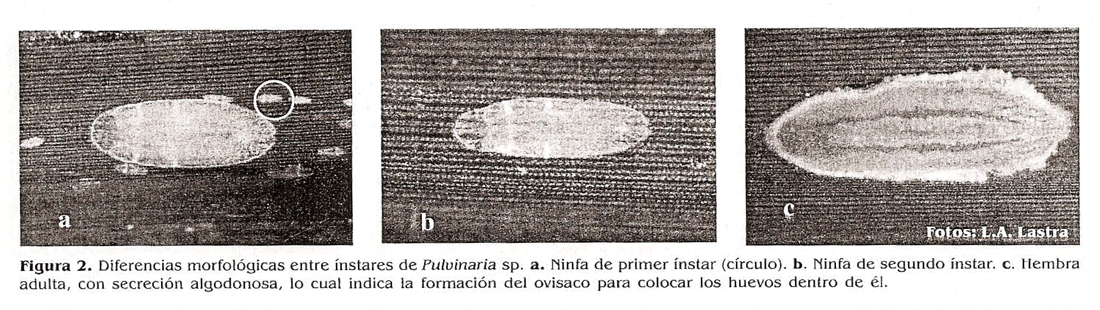
Huevo. es de color habano claro y a medi que se acerca el momento de eclosionar se torna rosado, mide aproximadamente 0,4 mm de longitud y es ovalado (fig.1a). 1er instar. En este estado el insecto rio se recubre de cera, es de color rosa fuerte, tiene una longitud promedio de 0,6 mm y 0,15 mm de ancho, seis segmentos antenales y setas anales (fig. 1b).

2do instar. En este instar el insecto se recubre de una cera o polvo harinoso de color blanco, conserva las setas anales y tiene una longitud promedio de 0,9 mm y 0.3 mm de ancho; en este momento, las ninfas presentan siete segmentos antena les y es el cambio más notorio en relación con el primer ínstar (Fig. 1c).
Adulto. Al igual que el segundo ínstar, el estado adulto aun se recubre de cera, tiene una longitud promedio de 3,22 mm y 1,7 mm de ancho, tiene siete segmentos antenales Se observo que la presencia de setas anales en los insectos sirve como medio para evitar que las gotas de las secreciones azucaradas caigan o se pierdan y por el contrario las mantienen cerca al insecto, facilitando a las hormigas su toma. Esta observación también la hizo Kunkel (1973) citado por llólIdobler y Wilson (1990) (Fig. 1c).
En el caso de los machos, el primer instar es muy similar al de las hembras y es a partir del segundo instar que se observan diferencias entre ellos. Entre las más notorias están el ancho del cuerpo, siendo el macho más delgado que la hembra, y la coloiscion, que es más oscura en el macho (fig.1d). Las diferencias morfológicas y el momento en el que muda este último fueron difíciles de determinar debido a que la ninfa, a partir del segundo instar, se envuelve en filamentos algodonosos formando una especie de capullo que dificulta la observación de las exuvias (fig.1e). El estado adulto es muy diferente a la hembra por ser un especímen alado de coloración rosa oscuro(fig.1f), con dos setas visibles en el ano; tiene una longitud promedio de 1.05 mm (u icluyeudo las setas, 4 mm, y posee 9 segmentos anlenales. Infortunadamente la población de machos no excedió los doce individuos durante el período total de observación, tazón por la cual se dificuil ó determinar aracterísticas tales como longevidad, el número (le instares y caracterización de los mismos, adcnias de que son muy frágiles y que el i iesgo de mortalidad fue mayor por manipulación o por contaminación del medio en el cual se mantuvo el tejido vegetal.
Pulvinaria sp.
En el caso de Puk’uiaria sp. no se observaron machos deniro de las poblaciones, por lo cual la determinación del ciclo de vida sólo se efectuó para las hembras. El ciclo total de la especie dura en promedio 76,7 días pasando por dos instares o estados ninfales y el estado adulto. El primei instar es móvil y prefiere el envés cte las hojas más jóvenes, mientras que el segundo y el estado adulto, una vez se fijan en el envés, no se vuelven a mover de ese lugar. Estos datos concuerdan con lo expresado por Hammon y Williams ti 984), que registi aioii dos o tres estados ninfales para la tamilia Coccidae, segun la especie considerada. Una vez el insecto alcanza el estado inmadnro perinailece un Periodo sin cambios aparentes adulto joven), se da inicio a la formación de huevos (adulto en preoviposición) y finalmente ocurre la oviposición (Tabla 2). No son gregarios y se distribuyc’n por toda la planta causan do un amarillamiento y necrosamiento en las hojas, que se inicia en los bordes. flajo condiciones de alta infestación apaiece lumagina. Al no haber encontrado machos, se asume que la forma de reproduccióu es partenogénica bajo condiciones del valle geográfico del río Cauca.
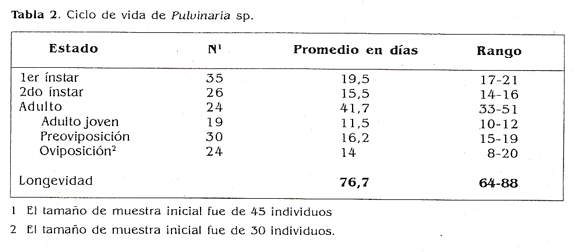
En cuanto a la duración del adulto y al promedio de huevos por hembra, los individuos provenientes de campo, dispuestos en hojas de caña en condiciones de laboratorio, colocaron un promedio de 218,2 (± 120,8) huevos por hembra, con un rango que varió entre 54 y 450 en el momento del conteo; es decir, que se despegaron las hembras de las hojas para contar del número de crías en ese instante. Con la segunda metodología, que consistió en hacer un seguimiento del desarrollo y de su progenie a las hembras criadas en laboratorio sobre plantas de caña, se constató que pusieron un promedio de 179,3 (t 179,4) huevOs por hembra con un rango entre 15 y 815, en un período de 20 días. La duración del adulto fue en promedio de 14 (± 4,1) días. El número de crías que registra la literatura en el caso de Puluinaria iceryl es de más de 1.200, en hembras desarrolladas sobre caña de azúcar. Por tanto, se cree que varios factores pudieron afectar la descendencia baja con la segunda metodología: el cambio de las condiciones ambientales (T°y H. R). la edad de la planta en que se criaron las hembras, el espacio en donde se desarrollaron y la manipulación que sufrieron como resultado de la cría en laboratorio, en comparación con las hembras de campo desarrolladas en plantas más vigorosas, y bajo condiciones diferentes.
La diferenciación morfológica de los ínstares fue más evidente en el número de setas marginales alrededor del cuerpo. específicamente entre los ojos, en el tamaño del cuerpo y en la presencia de setas anales, La descripción de las características anteriores se hace a continuación para cada ínstar.
H uevo. Mide aproximadamente 0,5 mm de longitud, es de color crema y se va oscureciendo a medida que se acerca el momento de la eclosión. 1er ínstar. Es de color habano, tiene una longitud promedio de 0,5 mm y posee dos setas anales largas; el número de setas marginales entre los ojos varía entre 9 y 11, y no hay presencia de bandas dorsales (Fig. 2a)
2do instar. Longitud promedio de 1,3 mm. pierde las setas anales, el número de setas marginales entre los ojos varía entre 20 y 25 y las bandas dorsales comienzan a hacerse visibles (fig. 2b).Adulto. Longitud promedio de 3,5 mm (hembras de laboratorio), aunque en algunos casos pueden llegar a medir hasta 1 cm; setas anales ausentes, número de setas marginales entre 30 y 40, bandas dorsales de color marrón que se oscurecen a medida que el insecto se acerca al momento reproductivo, además de la aparición de secreciones algodonosas alrededor del abdomen de la hembra, lo cual también es un indicativo de madurez y oviposición. Esas secreciones algodonosas son el ovisaco que la hembra forma para mantener y proteger su cría hasta que eclosionan las ninfas (Fig. 2c).
Reconocimiento y evaluación de enemigos naturales sobre saccharicoccus sacchari y Pulvinaria sp.
En visitas realizadas al campo, se encontraron escamas secas y con perforaciones en el cuerpo, al igual que algunos piojos harinosos con coloraciones sintomáticas de parasitismo. cuerpos momificados con perforaciones y en algunos casos desarrollos miceliales de color verde oliva en hembras adultas. Por otro lado, en invernadero también se cletectó la presencia de enemigos naturales atacando a Pulvinaria sp. Ir tal razón, se realizó un reconocimiento de los enemigos naturales de estos chupadores en las zonas de Corinto y El Cerrito Complementariamente, se determinó la incidencia de ataque de estos organismos sobre la escama Puivinaria sp. y el piojo harinoso.
El material de S. sacchari, recolectado en Corinto, se obtuvo de suertes con niveles de infestación bajo. Los resultados obtenidos (Tabla 3) mostraron que de 45 individuos recolectados en tres suertes, 21 mostraron síntomas de parasitismo para un porcentaje cercano al 50%. De éstos, emergieron 161 individuos pertenecientes a dos especies (sp. 1 y sp. 2) de la familia Encyrtidae. De la especie 1 se obtuvieron 108 individuos y de la sp. 2 53, resultando una relación de 2:1 entre las especies.
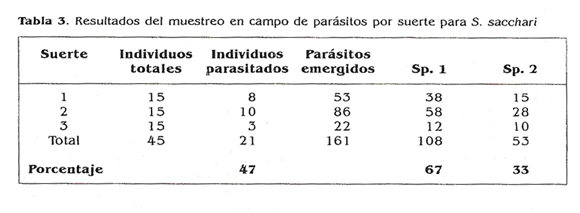
Por otro lado, dentro en una colonia conformada por 13 hembras maduras junto con su cría, se registró la presencia de una mosca Cecidomyiidae cercana al género Diadoplosis y de la cual se registraron 27 adultos; las larvas de esta mosca depredan los huevos de S. ,sacchari y se pueden hallar entre una y tres larvas por ovisaco. Este mismo individuo se encontró en la zona cJe Piedechinche, donde de 47 hembras adultas de la escama se recuperaron 25 moscas. En la muestra coleccionada en la Hacienda Piedechinche, de 15 hembras en aislamien.. Lo, se encontró que dos resultaron parasitadas: de una de ellas, emergieron cuatro adultos de la especie 1 y de la otra emergieron tres individuos de la especie 1 y cinco de la especie 2 , ya registradas en Corinto. Al emerger dos especies de un sólo individuo se plantea la pregunta: ¿se trata de una relación de hiperparasitismo? o ¿un caso de parasitismo múltiple?. Aún cuando existen especies hiperparásitas dentro de esta familia (borror et al. 1992; Ooulet y Huber 1993), hubo evidencias de que se trataba más bien de parasitismo múltiple teniendo en cuenta que la emergencia de ambos parásitos fue casi simultánea.
La evaluación de parasitismo sobre Pulvinana sp. en campo resultó ser un poco más difícil considerando que sólo se consiguió un campo en Corinto con una infestación escasa, del cual se recolectó una muestra de 45 individuos, que se mantuvieron en las unidades de aislamiento. De las dos unidades emergieron 24 parásitos, 22 emergieron de hembras de 2 ínstar (92%) y 2 de hembras adultas (8%), manifestándose una marcada preferencia del parásito por hembras de segundo ínstar. Al contabilizar el número de perforaciones de emergencia de las escamas ocasionadas por el parásito dio como resultado 1,33 (± 0,48) perforaciones por hembra en promedio. Los individuos fueron identificados por la Dra. María del Pilar Hernández de CJAT como correspondientes al orden llymenoptera, familia Encyrtidae.
Bajo condiciones de invernadero se encontró en forma abundante la presencia de Diadoplosis coccidivora Pelt (Diptera: Cecidomyiidae), especie que ya había sido registrada e identificada por el CABI. La hembra de la mosca pone sus huevos sobre o en las proximidades de la escama, luego cuando eclosionan las larvas se ubican bajo las hembras e inician su actividad depredadora de huevos. Considerando que esta evaluación se realizó en un experimento en donde se disponía de plantas con Pulviriaria sp. con asistencia de la hormiga loca y de plantas con Pulvinaria sp. sin su asistencia, se determinó el parasitismo bajo ambas condiciones. En presencia de la hormiga loca, éste fue del 53,2%, en tanto que cuando la escama no es asistida por la hormiga éste ascendió a 85,1%. El análisis de datos indicó que los promedios fueron diferentes con una significancia del 6%, que si bien no es alta permite suponer que efectivamente las hormigas ejercen un grado de protección a las escamas de las moscas depredadoras, confirmando lo registrado en la literatura acerca de la protección que le pueden dar a las escamas de sus enemigos naturales (fig. 4 a).

De igual manera, pero de forma más esporádica, se encontró a Anicefus annula tus (Hym: Encyrtidae) otro parásito detectado con anterioridad e identificado igualmente por el CABI (fig. 4b y 4c).
Reconocimiento de hongos entomopatógenos sobre S. sacchari
La detección de piojos rosados enfermos no siempre se llevó a cabo. Existen condiciones posiblemente asociadas con altas precipitaciones y alta humedad relativa que determinan el ataque del hongo al piojo harinoso y cuando éstas se dan, la incidencia puede ser alta, y por lo tanto, si la incidencia es nula se traduce en la ausencia de dichas condiciones. El hongo hallado afectó, bajo las condiciones en que se hizo el muestreo, 53,2 % de los individuos (8 = 65), atacando el 94% de hembras adultas (8 = 16) y el 83% de las ninfas de segundo ínstar (11 = 23), mientras que ninguno de los individuos de primer ínstar fue afectado (8 = 26). Los individuos atacados se ubicaron principalmente en el tercio inferior que es donde se localizan la mayoría de los piojos en el tallo. La sintomatología manifestada fue un desarrollo micelial verde, acompañado de la momificación del piojo afectado. El patógeno fue identificado como Aspergillus sp. (fig. 5). En un principio fue considerado como un patógeno secundario, pero después se aceptó, con base en la literatura, que se trataba de Aspergillus pos parasiticus. El gran problema de esta especie, junto con otras pertenecientes al mismo género. es la producción de aflatoxinas, las cuales son catalogadas como carcinogénicas 5, hecho que limitaría su producción masiva, en el caso de considerarlo como un agente promisorio de control del piojo harinoso.
Conclusiones
-Las condiciones de cría tanto de saccharicoccus saccízari como de Pulvinaria sp. influyen en el desarrollo del ciclo de vida de ambas especies y en su fecundidad.
- A pesar de que los cambios morfológicos en ambas especies no son muy notorios se pudieron determinar algunos de ellos que sirven de referencia al momento de conocer el instar en que el insecto se encuentra en un momento dado. En el caso de S. .sacchari la característica morfológica principal es el tamaño del insecto y el número de segmentos antenales para diferenciar el primer ínstar del segundo. Para el caso de Puluinaria sp., una de las características morfológicas importantes es el número de setas existentes en la periferia de la cabeza entre los ojos, el cual varía entre instares.
- Tanto en condiciones de campo como de invernadero se encontraron diferentes parásitos dentro de las poblaciones de ambos homópteros; sin embargo, su actividad controladora se afectó, cuando los insectos se encuentran en compañía de la hormiga loca, lo cual verifica la protección otorgada por ella, mencionada en la literatura.
- A pesar de que Aspergillus pos parasiticus afecta individuos deS. .sacchari en campo, la producción masiva y posible liberación del hongo se ve limitada por producir sustancias carcinogénicas, además de ser condicionado por aspectos ambientales específicos para desarrollar- se, situación inmanejable en campo.
Agradecimientos
Los autores agradecen a la Doctora María del Pilar Hernández (dAT), al doctor Ramiro Besosa, a la doctora Yolanda Gutiérrez (INCAUCA), al Doctor Carlos Enrique Gómez (CORPOICA) por su colaboración; al personal del laboratorio de entomología (Cenicaña) por su apoyo y disposición y a COLCIENCIAS y CENICAÑA por la financiación del proyecto.
Literatura citada
AUTREY, L. J.; BOOLELL, S.; JONES, E; LOCIÇHART, B. E.; NADIE A. 1995. Distribution of sugarcane Bacilliform Virus in various geographical regions. ISSCT. Proceedings XXI Congress 2. p. 5-14. March 1992. Bangkok-Thailand. Ksekart Univ. Press. [ Links ]
AVELLANEDA, M. C.; DIAZ-GRANADOS, C.; AN GEL, J. C.; VICTORIA, J. I. 2001. transmisión del virus baciliforme de la caña de azúcar (Sugarcane Bacilliform badnavirus ScBV) empleando saccharicoccus sacchari como vector. Cenicaña XXII Congreso de la Asociación Colombiana de Fitopatología y Ciencias afines. Medellín. 48 p. [ Links ]
BEARDSLEY, J. 1960. Observations On sugar cane mealy bugs in Hawaii. Journal Series of the Experimental Station, Hawaii an Sugar Planters. [ Links ]
BORROR, O.; TRIFLEI-IORN, C. H.: JHONSON, N. 1992. As introduction to the study of insects. Sexta edición. Edit. Harcourt Brace College Publisher. Ohio. U.S.A. 873 p. [ Links ]
De BARRO, E J. 1990. Natural enemies and other species associated with saccharicoccus sacchari (Cockerell) (Hemiptera: Fseudoccocidae) in the Bundaberg area, southeast Queensland. J. Aust. Ent. Soc. 29: 87-88. [ Links ]
ETIENNE, J. 1977. Pulvinaria iceryi a potential pest of sugarcane. XVI Congress 1. 9-20Sept. 1977. Sao Paulo-Brasil. International Society of Sugarcane Technologist. 719- 728. [ Links ]
GOULET, H. HUFER, J. 1993. Nymenoptera of the world: An identification guide to families. Centre for land and biological resources research . Otawa, Ontario 668 p. [ Links ]
HAMMON, A.; WILLIAMS, M. L. 1984. The soft scale insects of Florida (Homoptera: Coccidae: Coccoidea). 11. Florida Department of Agriculture & Consumer Services. [ Links ]
HOLLDOBLER, B.; WILSON, E. 0. 1990. The Ants. The Belknap press of Harvard University Press Carnbridge, Massachusetts. [ Links ]
INkERNAN, E A.; ASHBOL, N. J.; CARVER, M.: WILLIAMS, D. J. 1986. Observations on the pink Sugarcane mealy bug, saccharicoccus sacchari (Cockerell) in Australia (Nomoptera: Pseudococcidae). Indonesia. XIX Congress of ISSCT (1): 612-618. [ Links ]
MckENZIE, H. L. 1967. Mealybugs of California, with taxonomy, Biology and Control of North American Species (Homoptera: Coccoidea: Pseudoccocidae). University of Californa Press. Berkeley, Los Angeles. [ Links ]
NIEVES, J. J . 1999.Manejo, Prevención y Control de la Hormiga Loca Paratrechina fulva (Mayr), en caña para panela. ICA. Bogotá. D. C. p. 4-21. [ Links ]
WILLIAMS, O. J. 1985. Australian mealybugs. British Museum (Natural History) Australia, 431 p. [ Links ]
ZENNER DE POLANIA. I. 1985. Hábitos alimenticios y relaciones simbióticas de la ‘Hormiga Loca” Nyglanderia fuina con otros artrópodos. Revista Colombiana de Entomología 11(1): 3-10. [ Links ]
Recibido: Abr. 22/2003 Aceptado: Abr. 03/2004


{kind=link}
{kind=link}