










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkRevista Colombiana de Entomología
versão impressa ISSN 0120-0488versão On-line ISSN 2665-4385
Rev. Colomb. Entomol. v.31 n.1 Bogotá jan./jun. 2005
Efecto del medio de cultivo en la virulencia de Nomuraea rileyi sobre Spodoptera frugiperda(Lepidoptera: Noctuidae)
Effect of the culture médium in the virulence of nomuraea rileyi on Spodoptera frugiperda
(Lepidoptera: Moctuidae)
LUISA FERNANDA CARO C.1, LAURA FERNANDA VILLAMIZAR R.2, CARLOS ESPINEL C.3,ALBA MARINA COTES4
1. Estudiante Microbiología industrial, Pontificia Universidad Javeriana, Bogotá. E-mail: luisa_donald@hotmail.com
2. Investigadora. Laboratorio de Control Biológico. Program. MIR C. j. Tibaitatá - Corpoica. Km 14 vía Mosquera. Teléfono 4227300 ext 1409. E-mail: laurafernandav@yahoo.es
3. Investigador. Laboratorio de Control Biológico. Programa MIR C. I. Tibaitatá - Corpoica. E-mail: cespinel@yahoo.com
4.Ph. D. Fitopatología. Investigadora. Laboratorio de Control Biológico. Programa MIP. C. I. Tibaitatá - Corpoica. E-mail cotesprado@hotmail.com
Resumen. En un estudio previo en laboratorio, se seleccionó el aislamiento Mm005 del hongo por ocasionar una mortalidad del 95% sobre el gusano cogollero del maíz Spodoptera frugiperda, la plaga más importante de este cultivo. Con miras a la producción de conidios de alta calidad y virulencia que permitan desarrollar un bioplaguicida eficiente, el objetivo del presente trabajo fue el de determinar el efecto del medio de cultivo y de algunos potenciales inductores de virulencia sobre las características microbiológicas de los conidios de fiomuraea rileyi y su actividad biocontroladora sobre S. frugiperda. Para tal fin, el hongo fue crecido en los sustratos millo, arroz, avena y soya. Se evaluó el rendimiento de conidios en los sustratos, la germinación y actividad biocontroladora de los mismos, seleccionándose el medio a base de soya, en el cual se obtuvieron los conidios con las mejores características. Adicionalmente, el hongo fue crecido en medio extracto de levadura-malta suplementado con potenciales inductores de virulencia que consistieron en dos inductores proteicos, un homogeneizado de larvas, un inductor no específico, un inductor a base de un polímero y un control sin inductor. Mediante un bioensayo se seleccionaron los inductores proteico 1 y polímero 1 por causar un aumento en la actividad biocontroladora. Posteriormente, se evaluó el efecto combinado del sustrato de producción masiva y los potenciales inductores de virulencia seleccionados sobre la germinación y actividad biocontroladora del hongo. Se encontró que el inductor polímero 1, combinado con el medio de producción, presentó un efecto positivo sobre las características evaluadas. Los resultados obtenidos sugirieren que el tipo y composición del medio de cultivo, determinan las características microbiológicas y la virulencia de la biomasa producida.
Palabras clave: Bioensayo. Inductor de virulencia. Patogenicidad. Producción masiva.
Summary. Isolate Nm005 of nomuraea rileyi was selected in a previous study by causíng 95% mortality in the fall armyworm Spodoptera frugiperda, the most important pest in maize crops. In order to produce hígh quality conidia for developing an efficient biopesticide, the objective of the present worK was to determine the effect of the culture médium and some potential virulence inductors on the microbiological characteristics and biocontrol activity of the ¡i. rileyi conidia. The fungus was grown on miliet, rice, oat and soybean substrates. After evaluating yield, germination and biocontrol activity of produced conidia on the substrates, the soybean médium was selected because the conidia presented the best characteristics. In addition, the fungus was grown on yeast-malt extract agar médium supplemented with several potential inductors of virulence that consisted of two proteic inductors, a homogenization of larvae, a non specific inductor, an inductor based on a polymer and a control with no inductor. By using a bioassay, the proteic inductor 1 and polymer inductor 1 were selected because they caused an increase in biocontrol activity. Afterwards, the combined effect of the selected substrate for mass production and the selected potential inductor of virulence on the germination and biocontrol activity of the fungus was evaluated. The polymer inductor 1, combined with the culture médium, liad a positive effect on the characteristics evaluated. The results obtained suggest that the type and composition of the culture médium determines the microbiological characteristics and virulence of the biomass produced.
Key words: Bioassay. Inductors of virulence. Pathogenicity. Mass production.
Introducción
El daño ocasionado por el gusano cogollero del maíz, SpofJopíera frugiperda (J. E. Smith, 1797), es quizá uno de los más importantes a nivel económico, debido a que es una especie polífaga que se encuentra difundida en todos los países americanos, provocando grandes pérdidas en los rendimientos de muchos cultivos (García 2000).
Durante mucl10s años,'para reducir los efectos nocivos de S. frugiperda. se ha dependido del uso de insecticidas químicos. utilizando el doble o el triple de las dosís indicadas (pérez 2003). En muchas ocasiones la efectividad ha sido baja, debido a las aplicacíones realizadas después del momento crítico de la plaga y de la etapa fenológica más apropiada del cultivo o cuando los daños son irreversibles; incluso se ha pretendido aminorarla cuando prácticamente el cultivo alcanza un tamaño que imposibilita la entrada de las máquinas al campo; además, éste ha traído como consecuencia el desequilibrio biológico desde el establecimiento del cultivo (García y Pulido 1984).
La creciente necesidad de reducir el uso de agroquímicos para el control fitosanitario de los cultivos, hace necesario desarrollar tecnologías que permitan de forma práctica, económica y efectiva obtener productos a partir de microorganismos, insectos o nernátodos con calidad y en cantidades suficientes para su aplicación masiva en las zonas cultivadas (Ferrer 2001).
S. frugiperda es atacado por una gran diversidad de enemigos naturales como parasitoides, depredadores y patógenos, entre los cuales se destaca el hongo Nomuraea rileyi (Farlow) Samson (Hiphomycetes). Este hongo entomopatógeno se encuentra distribuido en amplios agroeco-sistemasy zonas geográficas, donde juega un papel importante en la regulación de las poblaciones, no sólo del cogollero sino de otras especies de importancia económica como: Atiticarsia gernmatalis (Hubner) (Lepidoptera: Noctuidae), Pseudoplusia includens (Walker) (Lepidoptera: Noctuidae), TYichoplusia ni (Hubner) (Lepidoptera: noctuidae) y Helicoverpa uirescens (F.) (Lepidoptera: Noctuidae), lo cual hace que este hongo se pueda aprovechar como agente biocontrolador potencializando su virulencia (León y Pulido 1991).
En el laboratorio de Control Biológico del Programa de Manejo Integrado de Plagas de Corpoica, Centro de Investigaciones Tibaitatá, se estandarizaron diferentes sustratos de producción masiva para la cepa de rí. rileyi Mm012, la cual fue seleccionada por su alta eficiencia para el control de 5. frugiperúa (Arriero 2001). Sin embargo, este aislamiento presentó disminución en la actividad biocontroladora, al ser cultivado en medios de producción masiva y al ser sometido a procesos de formulación, además de ser un aislamiento muy variable en su crecimiento. Por tales razones, fue necesario iniciar los estudios de desarrollo con otro aislamiento de /Y. rileyi codificado como Nm005, el cual causó porcentajes de mortalidad altos del insecto bajo condiciones de laboratorio (95% mortalidad acumulada) y es una cepa de menor variabilidad (Villamizar et al. 2002).
Para continuar con las investigaciones tendientes al desarrollo de un bioplaguicida con este aislamiento, el objetivo del presente trabajo fue el de seleccionar un medio eficiente para la producción masiva de conidios y determinar su efecto sobre la actividad biocontroladora de este aislamiento, así como buscar potenciales inductores de virulencia, con miras a la producción de conidios de alta calidad que permitan desarrollar un bioplaguicida eficiente.
Materiales y Métodos
Se utilizó el aislamiento de nomuraea rileyi codificado como Mm005, presente en el Banco de Germoplasma de Microorganismos del Laboratorio de Control Biológico de Corpoica, procedente de un cultivo de maíz en la finca Panorama, Puerto Qaitán (Meta) aislado a partir de larvas de Spodoptera frugiperda infectadas naturalmente en campo y conservado en tubos con agar extracto de levadura - extracto de Malta (YM) inclinado; se realizaron subcultivos del hongo y se conservaron en el mismo medio.
Producción masiva de Nomuraea rileyi.
Se evaluaron los sustratos millo, arroz, soya molida y avena para la producción masiva de /Y. rileyi. Estos se prepararon y suplementaron siguiendo metodologías previamente estandarizadas en el Laboratorio de Control Biológico de Corpoica para la inducción de esporuiacion de aislamientos de ri. rileyi (Arriero 2001).
En bandejas de lámina galvanizada se pesaron 150 g de matriz sólida (millo, arroz) y se humedecieron con 150 mi de hidrolizado de proteína al 8%. Para el medio de soya se pesaron 200 g de soya molida y se humedecieron con 200 mi de hidrolizado de proteína al 8%. Para el medio con avena, se utilizó 67% de avena en hojuelas y 33% de fuente de proteína y se adicionaron 350 mi de agua, esta mezcla fue licuada hasta su total homogeneización y se adicionaron 250 g de la mezcla en cada bandeja. Éstas se esterilizaron durante 20 minutos a 119 °C y 15 libras de presión y cada bandeja se inoculó con 20 mi de suspensión de conidios, la cual se preparó en una solución de TWeen 80 al 0,1% y se ajustó a una concentración de lxlO6 conidios.mi-1, a partir de un cultivo en caja en medio YM incubado durante 15 días. Las bandejas se incubaron a 25°C y 55% de humedad relativa, con luz constante directa durante 10 días. Terminado el tiempo de incubación se evaluaron parámetros microbiológicos como germinación, rendimiento y actividad biocontroladora.
Germinación. Se tomaron 0,1 g de conidios puros y se llevaron a un volumen de 10 mi con Tween 80 al 0,1%. A partir de esta suspensión se realizaron dos diluciones seriadas y se sembraron en cajas de Petri con medio YM. Transcurrido el tiempo de incubación (24 h), se agregó una gota de azul de lactofenol con el propósito de teñir los conidios del hongo. En el microscopio con un aumento de 40X, se realizó la lectura del número de conidios totales (germinados y no germinados) en 10 campos ópticos seleccionados aleatoriamente, expresando los resultados como porcentaje de conidios germinados.
Rendimiento de conidios en los medios de producción masiva. Según la metodología utilizada por Arriero (2001) para evaluar el rendimiento de conidios en las bandejas, se tomaron tres muestras de 1 cm2 de medio de producción masiva esporulado de manera diagonal para cada unidad experimental. Los tres fragmentos por bandeja se homogeneizaron en 300 mi de Tween 80 al 0,1% con una batidora Hamilton Beach Scovill por 10 min a velocidad máxima. Posteriormente, la cantidad de conidios se evaluó mediante recuento en cámara de rieubauer y se expresó el resultado como conidios.mi"1. Cada valoración se realizó por triplicado, utilizando tres unidades experimentales (tres bandejas) por tratamiento.
Evaluación del efecto de los diferentes sustratos de producción masiva en la actividad biocontroladora de los conidios de Nomuraea rileyi NmOO5.
Se realizaron bioensayos en condiciones de laboratorio a 25°C y 70% de humedad relativa. Se utilizaron los tratamientos: conidios producidos en millo, arroz, soya y avena, un testigo absoluto y un testigo tratado, consistente en Tween 80 al 0,1%. Para tal fin, se preparó una suspensión madre tomando las esporas de cada sustrato con un pincel estéril; a partir de esta suspensión se realizaron diluciones ajustando la concentración a 1 x 107 conidios.mi1, la cual, según estudios previos, correspondió a la CL90 para el aislamiento rimO 12 (Bosaeía/. 2002). Se aplicó un volumen de 1,2 mi de cada tratamiento sobre el haz y el envés de hojas de higuerilla (Ricinus communis) previamente desinfectadas con una solución de hipoclorito de sodio al 0,5%. En cubetas plásticas de 16 onzas se colocó una servilleta húmeda, una hoja de higuerilla asperjada con el tratamiento y una larva de 5. frugiperda en tercer instar, proveniente de una cría del Instituto de Biotecnología de la Universidad nacional, cada recipiente constituyó una unidad experimental. El diseño experimental fue completamente al azar con treinta repeticiones por tratamiento. Se llevó a cabo el registro de mortalidad a partir de las lecturas realizadas los días cuarto, sexto y décimo hasta obtener el 100% de mortalidad en alguno de los tratamientos. Con el fin de evidenciar la presencia del hongo, las larvas muertas se colocaron en cámaras húmedas.
Los resultados se corrigieron calculando el porcentaje de eficacia mediante la fórmula de Schneider - Orelli (Ciba - Qeigv 1973)
Eficacia (%) =
Donde:
b= % de individuos muertos en el tratamiento
k= % de individuos muertos en el testigo
Efecto de los potenciales inductores de virulencia en la velocidad de crecimiento de N. rileyi Nm005. Se prepararon medios suplementados con los potenciales inductores de virulencia, utilizando tres cajas de Petri por tratamiento donde cada una constituyó una unidad experimental. Con un sacabocados de 4 mm de diámetro se tomaron fragmentos de un medio de cultivo con agar YM crecido por el hongo durante 15 días y se colocó un fragmento esporulado en el centro de caja de Petri con los medios suplementados. las cajas se incubaron por 9 días a 25°C. La lectura se realizó en los días tres, seis y nueve de incubación midiendo el diámetro de la colonia formada. El resultado se informó en milímetros.
Evaluación del efecto de potenciales inductores de virulencia sobre la actividad biocontroladora de los conidios de N. rileyi Nm005. ri. rileyi fue cultivado en cajas de Petri con medio semisintético YM, el cual se suplemento con los potenciales inductores de virulencia a una concentración de 0,5% P/R Los Inductores se adicionaron en forma de polvo con un tamaño de partícula inferior a 150 (i y estos consistieron en inductor proteico 1, inductor proteico 2, un homogeneizado de larvas de S. frugiperda, un inductor no específico, inductor polímero 1. También se realizó un cultivo en medio YM sin inductor como tratamiento control. Cada caja se inoculó con 0,1 mi de una suspensión de conidios ajustada a una concentración de 1 x ]O6 conidios.mi"', provenientes de un cultivo en caja en medio YM de 15 días de edad. Se incubaron durante 15 días a 25°C y con luz constante directa. Con los conidios provenientes de cada tratamiento se prepararon suspensiones y se realizaron bioensayos siguiendo la metodología descrita anteriormente, utilizando una concentración subletal del bongo equivalente a la concentración letal 50 encontrada en un estudio previo (105 conidios.mi1), para poder evidenciar un posible efecto inductor de virulencia (Bosa et al. 2004).
Efecto combinado del sustrato de producción masiva y el potencial inductor de virulencia sobre las características de /Y. rileyi (Nm005). Se evaluó el efecto combinado del sustrato de producción masiva escogido por su mayor rendimiento, suplementado con el inductor de virulencia seleccionado sobre la actividad biocontroladora mediante un bioensayo realizado con la metodología descrita previamente y utilizando la concentración subletal 103 conidios.mi'. El medio de producción masiva sin inductor constituyó el tratamiento control. También se determinó el efecto combinado del sustrato con el inductor sobre la germinación y el rendimiento de conidios empleando las técnicas descritas anteriormente.
Diseño experimental. Para todas las pruebas se utilizó un diseño completamente al azar con tres repeticiones por tratamiento. Los resultados se sometieron a diferentes pruebas estadísticas, así: a) un análisis de varianza y una prueba de Kruskal-Wallis para la germinación y el rendimiento cuando se seleccionó el sustrato de producción masiva, b) un análisis de varianza y una prueba de Kruskal-Wallis
para determinar el efecto combinado del sustrato de producción masiva y el potencial inductor de virulencia, c) la prueba de diferencia mínima significativa para el rendimiento y actividad biocontroladora cuando se evaluó el efecto combinado del sustrato de producción masiva y el potencial inductor de virulencia seleccionados y d) la prueba de Tukey para evidenciar el efecto de los potenciales inductores de virulencia en la velocidad de crecimiento y actividad biocontroladora de ñ. rileyi, utilizando una significancia de 0,05.
Resultados y Discusión
Selección de un medio de cultivo sólido para la producción masiva de conidios de Nomuraea rileyi Nm005
Germinación. Al evaluar la germinación de los conidios de n. rileyi producidos en los diferentes sustratos de producción masiva, se observó germinación desuniforme, aunque después de 48 h de incubación se obtuvo una germinación mayor al 95% con todos los tratamientos, con valores de 99,33, 97,83, 97,77 y 99,35% para las células provenientes de los sustratos millo, arroz, avena y soya, respectivamente (Fig. 1). Estos resultados indican que los conidios provenientes de todos los sustratos de producción masiva presentaron una germinación alta, la cual no se vio influenciada por el tipo matriz sólida utilizada. La prueba de Kruskal -Wallis (a 0,05) no detectó diferencias significativas entre las germinaciones de los conidios producidos en las diferentes matrices de producción valoradas. Este comportamiento podría atribuirse a que todos los sustratos evaluados le proporcionaron al hongo lo necesario para su desarrollo y la formación de las reservas endógenas necesarias para la posterior germinación de los conidios, corno lo informaron Lañe y Trinci en 1991, quienes afirman que las diferencias en la germinación de esporas de Beauueria bassiana depende de sus respectivas reservas endógenas. Estos autores observaron un aumento de la germinación de los conidios de B. bassiana cuando se produjeron en un medio con fuente de nitrógeno limitado, sugiriendo que al manipular las condiciones de crecimiento se pueden mejorar características de las esporas como longevidad y germinación. Los autores también observaron que las esporas cosechadas en fase estacionaria después de 80 h de incubación en un cultivo con fuente de nitrógeno limitada, contienen más reservas endógenas (glicógeno y lípidos) que las cosechadas en un cultivo con fuente de carbono limitada, lo cual determina su capacidad de germinación.
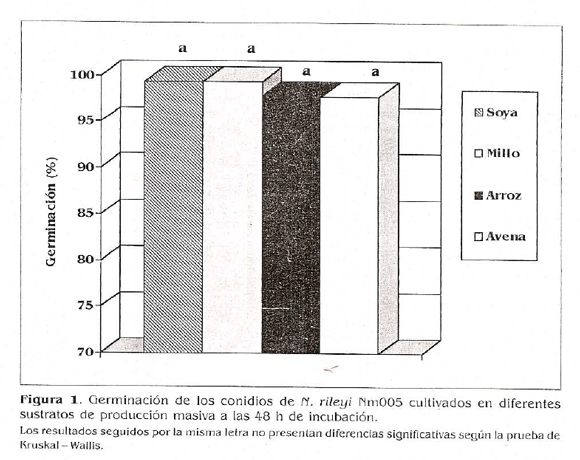
La germinación alta de los conidios de todos los tratamientos también podría atribuirse a un posible lavado de los autoinhibidores de germinación durante la preparación de las diluciones, las cuales se realizaron en todos los casos en una solución de Tween 80 al 0,1%, efecto que ha sido registrado por autores como Kuo (1999), quien observó que al lavar las esporas de Colleto-trichum gloeosporioides previamente a la evaluación de su germinación, obtenía porcentajes superiores a los logrados con las esporas que no habían sido sometidas al proceso de lavado. Este fenómeno lo atribuyó a un posible lavado de los autoinhibidores de germinación: gleosporona y la micosporina.
El Tween 80 también ha sido considerado como un estimulador de la germinación de conidios de Metarhizium anisopliae,actividad que se considera podrían tener diferentes detergentes aniónicos (Milner et al. 1990).
El hecho de que se haya presentado la misma capacidad de germinación en todos los sustratos de producción masiva, podría atribuirse a que los conidios producidos tenían la misma edad al momento de evaluarse la germinación, factor que ha demostrado ser determinante en la capacidad de desarrollo de los mismos (Qriffin 1981). Se ha observado que la germinación de los conidios también puede verse influenciada por factores como la composición del medio de cultivo en el cual se evalúe este parámetro, como lo informó Milner et al. 1990. En el presente estudio la germinación de /Y. rileyi fue valorada en el medio semisintético YM, cuya composición principalmente incluye: extracto de levadura, extracto de malta, dextrosa y peptona, los cuales probablemente le proporcionaron al microorganismo los nutrientes exógenos necesarios para germinar. Milner eí ai. (1990) observaron que los conidios de M. anisopliae germinaban pobremente en ausencia de nutrientes exógenos, y determinaron que la glucosa estimulaba su germinación, al igual que aminoácidos simples como la Lalanina y detergentes aniónicos como el TWeen 80. De igual forma, otros autores corno Dillon y Charnley (1990) observaron un aumento en la capacidad de germinación de los conidios de M. anisopliae, al adicionar al medio de cultivo Lalanina y Nacetylglu-cosamina. Estos autores, determinaron un aumento en la germinación del hongo, al incubar la muestra en agua destilada durante 20 h antes de la inoculación en cajas de Fetri, fenómeno que atribuyeron a una iniciación de la fase de preturgencia de los conidios, previa a la germinación de los mismos.
Bidochka y Khachatourians (1990) también observaron el efecto de fuentes de carbono, nitrógeno o fósforo presentes en el medio de evaluación, sobre la germinación de los conidios y registraron que al adicionar 0,5 mg/ml r\H2P04 a un medio de quitina crudo se incrementa la germinación del hongo del 10 al 43% después de 48 h.
Los resultados obtenidos en el presente estudio no mostraron diferencias significativas en cuanto a la germinación de los conidios provenientes de los diferentes sustratos de producción masiva. Este comportamiento posiblemente se debió a que estas matrices en su composición incluyen fuentes de carbono, nitrógeno, proteína, lípidos, macroy microelementos, que probablemente permitieron al microorganismo realizar la reserva endógena de nutrientes necesarios para formar el tubo germinal; como lo registraron Butt et al. (2001), quienes demostraron que la composición del medio de cultivo determina la formación de reservas endógenas en las células de las cuales depende su germinación, viabilidad y patogenicidad. La importancia de las reservas endógenas en la germinación y posterior desarrollo del tubo germinal se ha atribuido a que éstas son utilizadas como fuente de energía en el proceso de germinación y promueven el subsecuente crecimiento y desarrollo del microorganismo (Pascual 1998 citado por Butt eí ai. 2001). Cotter eí ai. (2000) afirman que sólo se requiere agua y oxígeno para la germinación de esporas que contienen su propia energía en forma de nutrientes endógenos ricos en carbono y nitrógeno.
Rendimiento de conidios de Nomuraea rileyi cultivados en los diferentes medios de producción masiva
El efecto del medio de cultivo en la capacidad de esporulación del hongo se evaluó mediante la cuantificación de la producción de conidios por unidad de área.
Las unidades experimentales se inocularon con 10 mi de una suspensión de conidios ajustada a una concentración de 1 x 10° conidios.mi1; es decir, cada bandeja se inoculó con lxlO7 conidios, lo que equivale a 1,4 x 104 conidios.cm2. Después de 19 días de incubación se obtuvieron rendimientos de 2,3 x 10s conidios.cm2 en el sustrato millo, 4,5 x 10ñ conidios.cm2 y 2,0 x 108 conidios.cm2 en los sustratos soya y arroz, respectivamente y la menor producción de conidios en el sustrato avena, con un recuento de 4,7 x 107 conidios.cm2. Como se puede observar en los resultados obtenidos, con el sustrato avena se obtuvo 1.000 veces el número de conidios inoculados, con los sustratos millo y arroz se obtuvo aproximadamente 10.000 veces más, y con el sustrato soya se alcanzó un rendimiento que equivalió a 20.000 veces lo inoculado.
El análisis no paramétrico de Kruskal-Wallis (a 0,05) indicó que el rendimiento de los conidios del microorganismo producido en sustrato soya fue significativamente superior con respecto al obtenido en el sustrato avena, pero no presentó diferencias significativas con los rendimientos obtenidos en los sustratos millo y arroz. El rendimiento en estos dos últimos sustratos tampoco presentó diferencias estadísticas con el resultado obtenido en el medio a base de avena (Fig. 2). Estos resultados sugieren que la composición del medio de cultivo tiene un efecto determinante en la esporulación del microorganismo
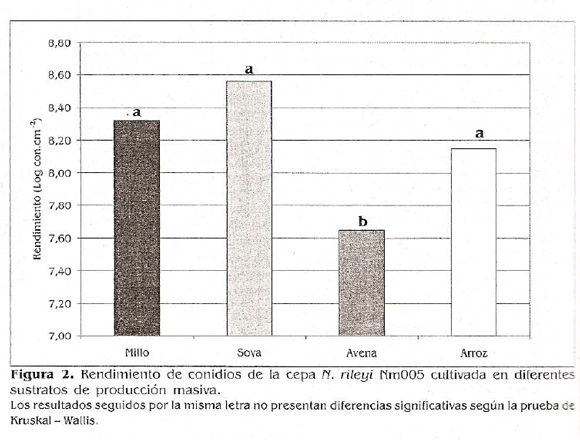
A pesar de no encontrarse diferencias significativas entre los resultados obtenidos con los sustratos soya, millo y arroz, se observa una tendencia numérica a una I" mayor producción en el sustrato soya, además de que en este medio se observó un > crecimiento uniforme en todas las bandejas y una abundante esporulación en las tres unidades experimentales. Adicionalmente, los conidios producidos en el e sustrato soya presentaron un aspecto e polvoso, desprendiéndose fácilmente del :_ sustrato y facilitándose su separación para e la evaluación microbiológica; característica :s deseable en la industria de bioplaguicidas n (Butt eí ai. 2001). Lo contrario ocurrió con n los medios de producción masiva de millo, y arroz y avena, en los cuales se obtuvieron a conidios muy húmedos y adheridos a la )- parte superficial del sustrato, haciendo,difícil su separación.
En el medio avena se observó un crecimiento poco uniforme, debido probablemente a que éste no tiene una estructura )Z de grano sino una consistencia semisólida. Este hecho podría haber generado una ó tasa de evaporación elevada que causó la 0 cristalización del sustrato, afectando el desarrollo del microorganismo obteniéndose una esporulación escasa. En los deis más sustratos millo, arroz y soya se >s mantuvo la humedad y el hongo colonizó completamente la matriz y esporuló abundantemente.
El crecimiento vigoroso de ñ. rileyi en el sustrato soya, probablemente se debió a que este medio en su composición incluía el hidrolizado de proteína, el cual es una fuente de nitrógeno orgánico indispensable para la conidiogénesis de este microorganismo (Riba y Qlandar 1980), además de ser una fuente de vitaminas y lípidos, cofactores de crecimiento que inducen esporulación (Villamizar 1998).
La esporulación menor en los sustratos avena y arroz podría deberse a que estos cereales poseen un alto.contenido de carbohidratos, lo cual favorece el crecimiento micelial del hongo y no su esporulación (Humphereys 1989 citado por Villamizar 1998). Por el contrario, el sustrato millo fue utilizado en forma de grano, el cual está recubierto por la cubierta seminal, que posiblemente ocasionó una escasa disponibilidad de nutrientes que favorecieron la esporulación de N. rileyi, ya que se ha registrado que las condiciones de estrés nutricional favorecen la esporulación (Griffin 1981). Sin embargo, en este último sustrato los conidios presentaron una alta humedad y se encontraron muy adheridos al sustrato, lo que dificultó su separación vía seca. En el caso del sustrato soya, este grano presentó un escaso contenido de carbohidratos (10,10% P/P) y un alto contenido de proteínas (41,33% P/P) que pudieron haber favorecido la esporulación del microorganismo. La importancia del nitrógeno para la conidiación de ñ, rileyi fue señalada por Riba y Clandard (1980), quienes evaluaron medios líquidos que contenían concentraciones de extracto de levadura de 0,5, 1 y 2% y encontraron que al aumentar la concentración de la fuente de nitrógeno se incrementó la esporulación del hongo, ya que dicho compuesto además de aportar fuente de nitrógeno, es una fuente de vitaminas y lípidos.
Al expresar los resultados del presente trabajo como conidios producidos por gramo de sustrato, se obtuvieron rendimientos de 1 xlO9 conidios.g_l para millo, de 1,5 x 109 conidios.g-' para soya, de 1,5 x 10e conidios.g-1 para avena y 8,8 x 108 conidios. g-1 cuando la producción se hizo en arroz, resultados similares a los obtenidos por Arriero (2001), quien evaluó el rendimiento expresado como conidios producidos por gramo de medio de cultivo esporulado, empleando como sustrato de producción salvado de trigo suplementado con diferentes fuentes de proteína. El autor obtuvo un rendimiento de 1,2 x 109 conidios.g-' para el medio suplementado con el hidrolizado de soya, 8,9 x 108 conidios.g"1 cuando utilizó hidrolizado de levadura y 1,4 x 10° conidios.g~' en el salvado enriquecido con solución de malta. Vimala-Devi (1994) evaluó diferentes sustratos para la producción masiva de fí. rileyi, en los cuales los rendimientos fueron similares a los obtenidos en el presente estudio, con una producción de 1,4 x 10a conidios.g-1 de sustrato, en un medio de sorgo triturado y suplementado con una solución de extracto de levadura al 1%.
Evaluación del efecto de los diferentes sustratos de producción masiva en la actividad biocontroladora de los conidios de Nomuraea rileyi Nm005
Al evaluar la actividad biocontroladora de los conidios de n. rileyi provenientes de los diferentes sustratos de producción masiva sobre larvas de S. frugiperda, se obtuvo un 100% de mortalidad a los 10 días después de iniciado el ensayo, con los conidios provenientes de todas las repeticiones de los tratamientos, utilizando una concentración de 1 x 107 conidios, mi, concentración letal 90 determinada para el aislamiento de N. rileyi (Fig. 3) (Bosa et al. 2002). El testigo absoluto no presentó mortalidad, confirmando que la muerte de los insectos en los tratamientos se debió a la actividad biocontroladora del microorganismo y no a condiciones ambientales o estrés en los individuos.
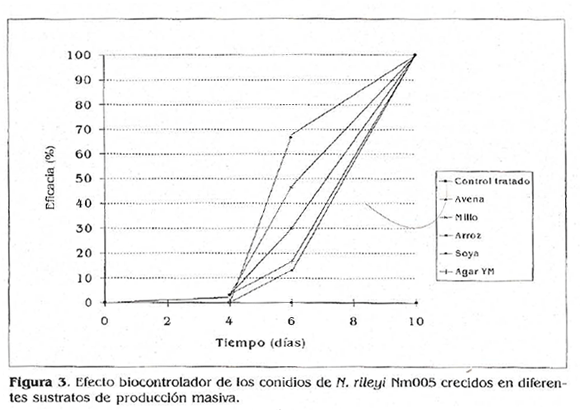
Todos los sustratos utilizados tuvieron el mismo efecto sobre la actividad biocontroladora del microorganismo. Sin embargo, se observó una cierta tendencia de los conidios producidos en el sustrato soya a ejercer más rápidamente su efecto biocontrolador.
Debido a la falta de variabilidad entre los resultados de las réplicas de cada tratamiento, no fue necesario aplicar una prueba estadística para determinar diferencias entre los mismos. Se observó que los conidios cultivados en los cuatro sustratos evaluados presentaron la misma virulencia y el medio de cultivo no tuvo efecto sobre la patogenicidad, pudiéndose utilizar cualquiera de ellos para la multiplicación del hongo. Este resultado se podría atribuir a que posiblemente todos los sustratos le proporcionaron al hongo los nutrientes necesarios para su desarrollo.
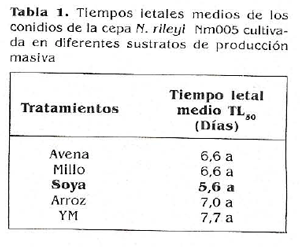
Con el propósito de seleccionar un sustrato de producción masiva se determinó el tiempo letal medio (TLM) de los conidios de ri. rileyi cultivados en los diferentes sustratos de producción masiva, obteniéndose valores de 5,6 días para soya, 6,6 días para avena y millo, 7 días para arroz y 7,7 días para los conidios cultivados en medio YM.
La prueba estadística de Kruskal-Wallis no detectó diferencias significativas entre los tiempos letales de todos los tratamientos (Tabla 1), confirmando que los conidios producidos en todos los sustratos son igualmente virulentos. Sin embargo, el tiempo letal medio más bajo fue obtenido con los conidios provenientes del cultivo en sustrato soya, lo cual indica que posiblemente en este sustrato se obtienen conidios que ejercen su actividad biocontroladora más rápidamente en comparación con los conidios de los demás tratamientos.
El tiempo letal menor obtenido con los conidios cultivados en el sustrato soya posiblemente se debió a una activación por parte del sustrato de algunas de las etapas determinantes en el mecanismo de acción de este microorganismo. Por ejemplo, pudo haber mejorado la capacidad de adherencia de las células, ya sea por la modificación en la carga de la pared celular de los mismos o por un aumento en la producción de mucílago que media el proceso de adhesión. También el sustrato podría haber aumentado la capacidad de germinación de los conidios, sus sistemas enzimáticos o la producción de toxinas, que determinan la virulencia del microorganismo (Lezama 1994).
El fríjol soya tiene un contenido proteico de 41,3% que sumado al 49,46% de proteína contenido en el hidrolizado, constituyeron la fuente de nitrógeno orgánico para el microorganismo, la cual se encontró en alta concentración, mientras que la disponibilidad de carbohidratos en este medio de producción masiva fue limitada. Esta mayor disponibilidad de nitrógeno y escasa de carbono, además de favorecer la esporulación del hongo, pudo haber jugado un papel importante, en la patogenicidad. Las enzimas fúngicas que han sido señaladas como determinantes en el mecanismo de acción, tienen un mecanismo de represión catabólica por exceso de fuente de carbono (St Leger. et al. 1986 a,b,c) y por lo tanto con este medio con fuente de carbono limitada, podrían haberse activado dichos sistemas enzimáticos, obteniéndose conidios más virulentos. En el caso de los demás sustratos, los mayores tiempos letales obtenidos posiblemente se debieron a que en su composición incluyen una mayor cantidad de carbohidratos que de proteína, ya que por ejemplo el arroz contiene un 8,22% de proteína y un 55,15% de carbohidratos, la avena 12,5% de proteína, 53,84% de carbohidratos, el millo 13,35% de proteína y 46% de carbohidratos (Caro 2003). Esla disponibilidad mayor de carbono y menor de nitrógeno con respecto a la soya, posiblemente generó una represión catabólica de enzimas en los conidios producidos en estos sustratos, obteniéndose en consecuencia que el proceso infeccioso se desarrollara más lentamente (St. Leger et a/. 1986 a, b, c).
El contenido endógeno de las células depende de la composición del medio de cultivo y es inversamente proporcional a la disponibilidad cié nutrientes en el mismo (Griffin 1981). En el presente trabajo posiblemente el sustrato soya, el cual ocasionó el menor tiempo letal, pudo haber generado una mayor reserva endógena en los conidios, permitiendo que el proceso de infección se desarrollara más rápido. Este efecto ha sido registrado para Ulocladium atrum, hongo para el ctial se manipularon sus reservas endógenas, pudiéndose modificar su capacidad de establecimiento sobre las hojas y su actividad biocontroladora de Botrytis cinérea (Frey y Magan 1998; registrados por Butt et al. 2001).
Los resultados obtenidos permitieron seleccionar el sustrato soya como el más adecuado para la producción de conidios del aislamiento de /Y. rileyi rlmOOS, ya que con este medio se obtuvo el mayor rendimiento de conidios, los cuales mostraron una alta germinación y actividad biocontroladora, además de ser el único sustrato que se obtuvo una esporulación abundante y de apariencia pulverulenta que facilitaría su separación vía seca, características deseables en la industria de bioplaguicidas.
Efecto de los potenciales inductores de virulencia en la velocidad de crecimiento de N. riíeyi Nm005
La capacidad de desarrollo del microorganismo es un factor determinante de la virulencia y por lo tanto cualquier manipulación que permita mejorar la velocidad de crecimiento de un hongo entornopatógeno, podría también mejorar su actividad biocontroladora. Con el fin de evaluar el fecto de diferentes potenciales inductores de virulencia sobre la actividad biocontroladora de /Y. rileyi y relacionar dicha actividad con parámetros determinantes del mecanismo de acción, como la capacidad de desarrollo del hongo, se evaluó el efecto de dichas sustancias en la velocidad de crecimiento del microorganismo.
En los resultados obtenidos en este estudio se observó la mayor tasa de crecimiento cuando el microorganismo se cultivó en medio YM suplementado con el inductor polímero 1. Para este tratamiento la velocidad fue de 1,29 rnm/día, seguido por el inductor proteico 2 con 1,20 mm/día, el homogeneizado de larvas con 1,11 mm/ día, el medio YM utilizado como tratamiento control con 0,91 mm/día, el inductor proteico 1 con 0,88 mm/día y por último el inductor no específico que alcanzó una velocidad de 0,87 mm/día (Tabla 2; Fig. 4).
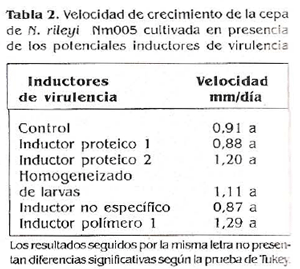
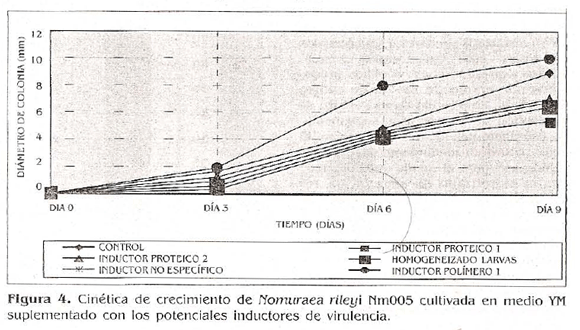
La comparación de medias de Tükey (o 0,05) no detectó diferencias en la tasa de crecimiento del hongo en medio YM suplementado con los diferentes potenciales inductores de virulencia, resultado que sugiere que ninguna de las sustancias evaluadas tuvo un efecto Inductor en la capacidad de crecimiento del microorganismo.
Este resultado probablemente se debió a la baja concentración de los inductores (0,5% P/F) utilizada para suplementar el medio YM. Dicha concentración no hizo un aporte nutricional significativo en los diferentes medios suplementados, por lo que no hubo un efecto en la velocidad de crecimiento, ya que como lo señaló Qriffin (1981) este parámetro depende de la composición del medio de cultivo.
A pesar de no encontrarse diferencias estadísticas entre los tratamientos evaluados, se observó una tendencia numérica a una mayor tasa de crecimiento cuando el medio se suplemento con el inductor polímero 1 con respecto a los demás tratamientos, lo que indica que el polímero 1 posiblemente estimuló el crecimiento del hongo. La tasa de crecimiento de muchos hongos depende del microorganismo y de las condiciones ambientales. Se ha indicado que la disminución en la tasa de crecimiento es causada probablemente por el suministro limitado de nutrientes y es directamente proporcional al tiempo (Qriffin 1981).
La velocidad de crecimiento micelial continuamente presenta variaciones que presumiblemente se deben a un sistema de control poligénico. ya que se ha observado que la tasa de crecimiento está asociada con diferentes regiones cromosomales (Larraya et al. 2002). En el presente estudio los resultados obtenidos no presentaron diferencias entre sí, posiblemente porque en todos los tratamientos los genes implicados con la tasa de crecimiento se expresaron de la misma forma, lo cual podría deberse a las diferencias mínimas en la composición de los medios.
La velocidad de crecimiento de algunos hongos se ha relacionado con las reservas endógenas presentes en las células, particularmente polioles como el glicerol que pueden servir como fuente de energía y ser utilizados como promotores del crecimiento y desarrollo (Butt et al. 2001). Las tasas de crecimiento obtenidas en este trabajo con los potenciales inductores de virulencia no presentaron diferencias estadísticas entre sí, posiblemente debido a que el fragmento colonizado con el hongo y utilizado para la inoculación de los medios suplementados provenía del mismo cultivo en caja de Petri con medio YM; razón por la cual los propágulos utilizados para inocular posiblemente tenían unas reservas endógenas similares que les permitieron crecer a velocidades similares.
Evaluación del efecto de potenciales inductores de virulencia sobre la actividad biocontroladora de los conidios de Nommraea rileyi NmOO5
Los conidios de ñ. rileyi crecidos en presencia de potenciales inductores de virulencia ocasionaron los mayores porcentajes de mortalidad sobre larvas de S. frugiperda con los tratamientos inductor polímero 1 e inductor proteico ], con 93,10 y 96,55%, respectivamente; seguidos por los obtenidos con el homogeneizado de larvas con un 89,65%, el inductor proteico 2 con 89,65 %, el inductor no específico con 75,86% y por último, la menor mortalidad se obtuvo con el tratamiento control YN con un valor de 65,46% (Fig. 5).
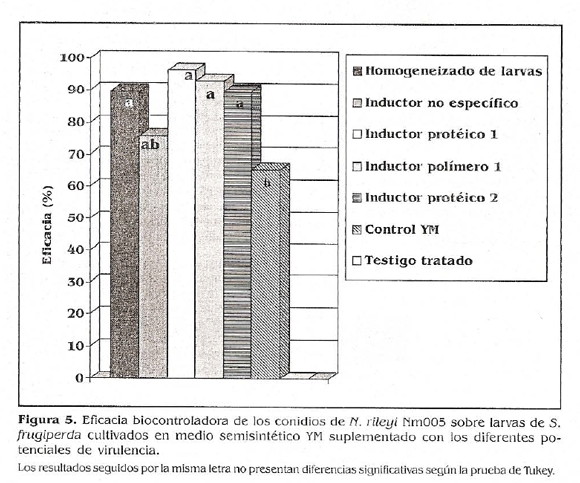
Los datos se sometieron a un análisis de varianza (AMAVA) y a una comparación de medias de Tukey (u 0,05). La actividad biocontroladora de los conidios provenientes del tratamiento control YM sin inductor fue significativamente inferior a las obtenidas con el microorganismo cultivado en el medio suplementado con el inductor proteico 1, el inductor polímero 1, el homogeneizado de larvas y el inductor proteico 2, no encontrándose diferencias entre estos últimos. Los resultados obtenidos con estos inductores no presentaron diferencias estadísticas con el presentado por el inductor no específico, el cual tampoco fue significativamente diferente del tratamiento control (YM) sin suplementar. Estos resultados sugieren que los inductores proteico 1, proteico 2, polímero 1 y homogeneizado de larvas tuvieron un efecto inductor sobre la actividad biocontroladora, lo cual posiblemente podría atribuirse a una activación por parte del sustrato de alguna de las etapas del mecanismo de acción, como los sistemas enzimáticos que son indispensables en la penetración de la cutícula o en la producción de toxinas.
La relación de la actividad enzimática con la patogenicidad ha sido registrada por autores como El- Sayed et al. (1989) citado por Lezama (1994), quienes compararon la virulencia de Í1. rileyi sobre la base de la actividad quitinolítica de dos cepas virulentas y una no virulenta contra Trichoplusia ni, trabajo en el que encontraron de 10 a 17 veces más actividad enzimática en las cepas virulentas que en las no virulentas.
El efecto del inductor proteico 1 podría deberse a una inducción de enzimas proteolíticas que han sido establecidas como determinantes en el mecanismo de penetración de la cutícula, debido a que está compuesta en un 70% de proteína (Clarkson y Charnley 1996).
En el caso del inductor polímero 1 pudo ocurrir una inducción de enzimas hidrolíticas como proteasas, esterasas y quitinasas, como M-acetilglucosamlnidasa, la cual es muy importante en el proceso de penetración. Algunos autores han relacionado la inducción de quitinasas con la composición del medio de cultivo, es el caso de Bosa et al. (2001) quienes observaron un aumento en la actividad de esta enzima al cultivar la bacteria Serratia marcescens en presencia de un homogeneizado de su hospedero Tecia sotanivora, insecto que tiene un alto contenido de quitina proveniente de su cutícula. Los autores adicionalmente informaron que al aumentarse la actividad de esta enzima también se incrementó la patogenicidad del microorganismo.
El inductor proteico 1 mejoró la velocidad de crecimiento del microorganismo, lo que posiblemente se podría relacionar con el aumento en la actividad biocontroladora del mismo, debido a que éste podría haber desarrollado más rápidamente el proceso de infección.
Los inductores proteico 1 y el polímero 1 ptidieron tener efectos en otras etapas del mecanismo de acción como la adherencia y la germinación. Este efecto fue registrado por Villamizar et al. (2001), quienes observaron un aumento en la germinación, adherencia e hidrofobicidad de los conidios de Metaihizium anisopliae cuando éste se cultivó en un medio suplementado con potenciales inductores de virulencia, viéndose relacionado con la actividad biocontroladora de la langosta llanera Rhatnmatocerus schistocercoides.
El inductor no específico no tuvo un efecto significativo en la actividad biocontroladora del hongo, lo que podría atribuirse a que este inductor tiene un bajo contenido proteico que no permitió que se activaran los sistemas enzimáticos implicados en la hidrólisis de estos componentes, los cuales como se mencionó anteriormente están estrechamente relacionados con la virulencia de los hongos entornopatógenos (Clarkson y Charnley 1996).
En el caso del homogeneizado de larvas, este inductor tuvo un efecto significativo sobre la virulencia de 11. rileyi, ya que el sustrato consistió en el insecto hospedero de dicho hongo, como lo registraron Bidochka y Khachatourians (1992), quienes observaron un aumento en la virulencia de B. bassiana cuando se cultivó en presencia de componentes cuticulares de su hospedero Melanoplus sanguinipes (Orthoptera: Acrididae).
Debido a que con el inductor polímero 1 y el inductor proteico 1 se alcanzaron los mayores porcentajes de mortalidad, estos inductores se seleccionaron para utilizar en combinación con el medio de producción masiva seleccionado (soya) para la producción de conidios de N. rileyi.
Efecto combinado del sustrato de producción masiva y ei potencial inductor de virulencia sobre las características de N. rileyi Mm005
Se evaluó el efecto de cultivar el hongo en el medio de producción masiva seleccionado, supíementado con los potenciales inductores de virulencia que presentaron los mejores resultados. Para tal fin se utilizó el sustrato soya supíementado con el inductor polímero 1 y con el inductor proteico 1.
Después de 40 h de incubación en medio YM, los conidios provenientes del sustrato soya supíementado con inductor polímero I presentaron porcentajes de germinación de 99,49%, siendo superiores a los porcentajes de germinación obtenidos con los conidios provenientes del medio suplementado con el inductor proteico 1 y del sustrato sin suplementar, los cuales presentaron una germinación del 76,54 y del 61,31%, respectivamente (Fig. 6). Estos resultados no se ajustaron a una curva normal y no presentaron homogeneidad de varianzas, razón por la cual se realizó un análisis no paramétrico utilizando la prueba de Kruskal-Wallis con una significancia de 0,05.
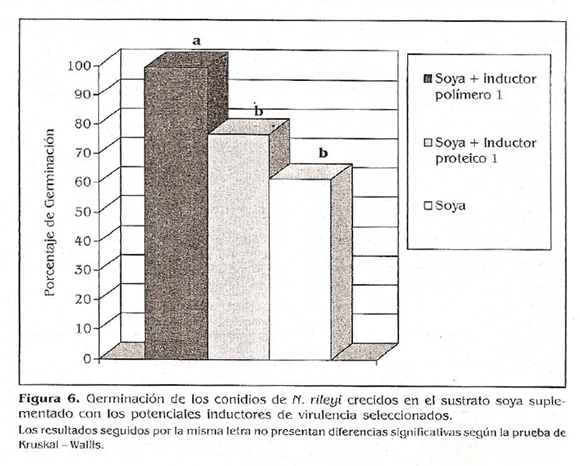
El análisis estadístico encontró que los conidios provenientes del sustrato soya supíementado con el inductor polímero 1 presentaron una germinación significativamente superior a la obtenida con los conidios provenientes del sustrato suplementado con el inductor proteico y sin suplementar, lo que indica que esta sustancia tuvo un efecto inductor de la germinación de los conidios producidos. Lo que podría deberse, como se mencionó anteriormente, a la activación por parte del polímero 1 de genes relacionados con la germinación (Kothe y Free 1998).
La germinación de los conidios cultivados en la soya suplementada con el inductor proteico 2 no presentó diferencias estadísticas con la germinación de los conidios cultivados en el sustrato soya sin suplementar. Resultado que sugiere que este inductor al ser combinado con el medio de producción masiva no tiene un efecto inductor de la germinación de N. rileyi.
Al evaluar el rendimiento de conidios en el sustrato soya supíementado con los potenciales inductores de virulencia seleccionados, se encontraron valores de 3,9 x 108 conidios.cm2 para el sustrato soya suplementado con el inductor proteico 1; 1,0 x ios conidios.cm2 para el sustrato soya supíementado con ei inductor polímero 1 y de 4,0 x 108 conidios.cm2 para el sustrato soya sin suplementar (Fig. 7). Los resultados se sometieron a un análisis de varianza (ANAVA) y a una comparación de medias de diferencia mínima significativa (a 0,05). Se encontró que el rendimiento de conidios en el sustrato soya supíementado con el inductor polímero 1 fue significativamente inferior a los obtenidos con los otros tratamientos, resultado que indica que el polímero 1 al ser combinado con el medio soya, disminuyó la capacidad de esporular del microorganismo.
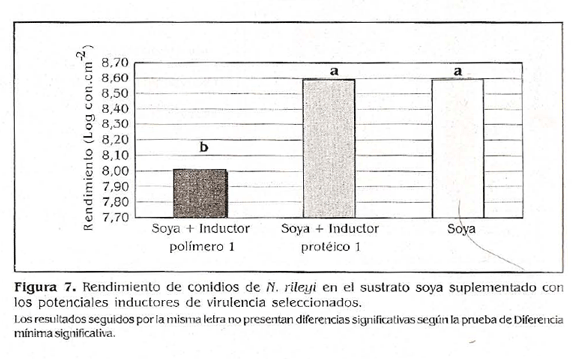
Los resultados de germinación obtenidos con el sustrato soya supíementado con el inductor proteico 1 y el sustrato soya sin suplementar no presentaron diferencias estadísticas entre sí, lo que indica que el inductor proteico 1 no afectó la conidio génesis del microorganismo. En el presente trabajo a pesar de haberse utilizado una concentración baja de inductores (0,5%), si se observó un efecto del polímero 1, posiblemente porque éste fue distribuido homogéneamente sobre toda la superficie del medio de producción donde fue cultivado el hongo.
La eficacia biocontroladora obtenida con una concentración subletal de los conidios cultivados en el medio de producción masiva a base de soya supíementado con el inductor polímero 1 fue de 62,07%, para los cultivados en el sustrato de soya sin suplementar, que constituye el tratamiento control, fue de 48,27 y de 44,83% para aquellos crecidos en el sustrato soya supíementado con el inductor proteico 1 (Fig. 8).
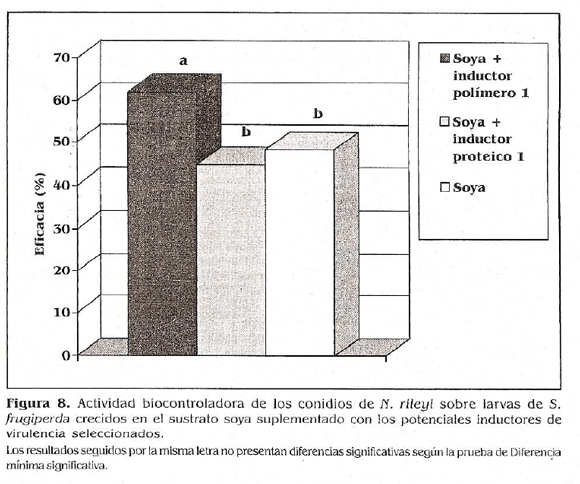
La comparación de medias mediante la prueba de diferencia mínima significativa (LSD) con (a 0,05) determinó que la actividad biocontroladora de los conidios cultivados en el sustrato soya supíementado con el inductor polímero 1 fue significativamente superior a la obtenida con el hongo crecido en el sustrato soya sin suplementar y en el sustrato soya suplementado con el inductor proteico 1. Este resultado confirma que el polímero 1 sí ejerce un efecto Inductor de la virulencia de este microorganismo, el cual no se vio afectado por la combinación con el medio de producción masiva. Los resultados obtenidos con el inductor proteico 1 no presentaron diferencias significativas con respecto a los obtenidos con el tratamiento control, sugiriendo que el inductor proteico i no tiene un efecto sobre la virulencia del microorganismo cuando éste es combinado con el sustrato de producción masiva a base de soya, mientras que si hubo una inducción de virulencia cuando fue combinado con el medio YM. Este resultado podría deberse a una represión catabólica de las enzimas implicadas en el mecanismo de acción por exceso de fuente de carbono en el medio soya. Posiblemente si fueron activadas por el inductor proteico 1 eri el medio YM que tiene un menor contenido de carbohidratos.
El aumento en la actividad biocontroladora del microorganismo cuando se cultivó en el medio soya con el inductor polímero 1, podría estar relacionado con el incremento en la capacidad de germinar de los mismos, ya que como lo registró Lezama (1994), estos dos factores están estrechamente relacionados. El autor determinó que la agresividad de una cepa de Conidiobolus obscurus estuvo en función de la velocidad con la cual fue capaz de germinar el microorganismo.
Los resultados de este trabajo permitieron seleccionar el sustrato soya suplementado con el inductor polímero 1 para la producción eficiente de conidios de ri. rileyi Nm005 de alta calidad, con un alto nivel patogénico que permitiría elaborar un bioplaguicida de calidad que permitiría reducir las dosis de aplicación, minimizando el impacto ambiental y disminuyendo los costos de producción al agricultor, ya que se requeriría menos producto para la aplicación.
Conclusiones
• Los conidios de riomuraea rileyi (Nm005) obtenidos en todos los sustratos de producción masiva evaluados presentaron alta actividad biocontroladora bajo condiciones de laboratorio.
• La composición del medio de cultivo afectó las características microbiológicas y de actividad biocontroladora de riomuraea rileyi <Nm005).
• Se seleccionó el sustrato soya por producir el mayor rendimiento de conidios, los cuales presentaron una germinación alta y ocasionaron el tiempo letal menor sobre larvas de Spodoptera frugiperda.
• El inductor proteico 1 y el polímero 1 mejoraron la velocidad de crecimiento y la actividad biocontroladora de riomuraea rileyi Mm005.
• La producción de conidios de riomuraea rileyi en el medio soya suplementado con el polímero 1 mejoró la germinación y la actividad biocontroladora.
Literatura citada
ARRIERO, C. 2001. Producción y caracterización de principios activos de un bioplaguicida a base de Nomuraea rileyi para el control de Spodoptera frugiperda. Bogotá. Trabajo de grado (Microbióloga Industrial). Pontificia Universidad Javeriana. Facultad de Ciencias. Carrera de Microbiología industrial. 122 p. [ Links ]
BIDOCHKA, M.; KHACMATOURIANS, G. 1990. Beaiweria bassiana submerged conidia production in a defined médium containing chitin, two hexosamines or glucose. Applied Microbiology and Biotechnology 33:641-647. [ Links ]
BIDOCHKA, M.; KHACHATOURIANS, G. 1992. Growth of the entomopathogenic fungus Beauueria bassiana on cuticular components from the migratory grasshopper, Melanoplus sanguinipes. Journal of lnvertebrate Pathology 59:165-173. [ Links ]
BOSA, F.; COTES, A. 2001. Efecto de las condiciones de cultivo sobre la actividad enzimática de Serrada marcescens contra Tecia solaniuora (Lepidoptera: Gelechiidae). Revista Colombiana de Entomología 30 (1): 79-85. [ Links ]
BOSA, E; CHAVEZ. D.; TORRES, L.; PARÍS, A.; VILLAMIZAR, L.; COTES, A. 2004. Evaluación de aislamientos nativos de tiomuraea rileyi para el control de Spodopiera frugiperda J. E. Smith (Lepidoptera: lioctuidae). Revista Colombiana de Entomología 30 (1): 93-97. [ Links ]
BÜTT, T.; JACKSON, C: MAGAN, N. 2001. Fungi as biocontrol agents. CABI publishing. p. 239-287. [ Links ]
CARO, L. F. 2003. Efecto de la composición del medio de cultivo en la virulencia de riomuraea rileyi sobre el gusano cogollero del maíz Spodoptera frugiperda (Lepidoptera: Moctuidae). Trabajo de grado (Microbiología industrial). Pontificia Universidad Javeriana. 149 p. [ Links ]
CIBA - GEIGY. 1973. Como realizar un bioensayo. Manual de ensayos de campo, p. 2-10. [ Links ]
CLARKSOM, J.; CHARNLEY, A. K. 1996. New insights into the mechanisms of fungal pathogenesis in insects. Trends in Microbiology. 4 (5): 197-202. [ Links ]
COTTER, D. A.; MAHADEO, D.; CERVI. D.; KISHI, Y.; SANDS, T; SAMESHIMA, M. 2000. [On line] Environmental regulation of pathways controlling sporulation, dormancy and germination utilizes bacterial - like signaling complexes in Dictyostelium discoideum. Protist 151: 111-126. [Consulta Septiembre 2003] [ Links ]
DILLON, R. J.; CHARNLEY, A. K.1990. Initiation of germination in conidia of the entomopathogenic fungus, Metarhizium anisopliae. Mycological research 94 (3): 299- 304. [ Links ]
FERRER, F. 2001. Biological control of agricultural insect pests in Venezuela; Advances, Achievements, and Future Perspectives. Biocontrol News and Information 22 (3): 67-74. [ Links ]
GARCÍA, F. 2000. Manejo integrado del gusano cogollero del maíz, Spodoptera frugiperda. En: Memorias I Curso Taller Internacional Control Biológico. Componente fundamental del MIP en agricultura sostenible. Programa MIP Bogotá - Colombia. p. 156- 161. [ Links ]
GARCÍA R.; PULIDO, F. J. 1984. El gusano cogollero del maíz Spodoptera frugiperda (J. E. Smith), su control y perspectivas para su manejo. En: XI Reunión de maiceros de la zona andina. II Reunión latinoamericana de maíz. Ministerio de Agricultura. Instituto Colombiano Agropecuario. Programa nacional del Maíz y Sorgo. Colombia diciembre 2-7. [ Links ]
GRIFFIN, D. 1981. Fungal Physiology, A while -Interscience Publication, USA. New York. p. 102-127. p. 260-279. [ Links ]
KOTHE, G.; PREE, S. J. 1998. The isolation and caharacterization of nrc - I and nrc -2, two genes encoding protein kinases that control growth and developmenl in rieurospora crassa. Genetics. Endocrinologyand Metabolism 149: 117- 130. [ Links ]
KUO, K. 1999. Germination and appressorium formation in Colletotrichum gloesporioides. Proc. Nati. Sci. Counc. Vol 23 (3): 126-132 [ Links ]
LANE, B.; TRINCI, A. 1991. Influence of cultural conditions on the virulence of conidia and blastospores of Beauveria bassiana to the green leafhopper, Hephotettix virens. Mycol. Res. Gran Bretaña. 95 (7): 829-833. [ Links ]
LARRAYA, L. M.; IDARETA, E.; ARANA, D.; RITTER, E.; PISABARRO, A. G. 2002. Quantitative trait loci controlling vegetative growth rate in the edible Basidiomycete Pleurotos ostreatus. Applied and Environmental Microbiology 68: 1 109-1 I 14. [ Links ]
LEÓN M., G. A.; PULIDO, J. L. 1991. Importancia del control natural del cogollero Spodoptera frugiperda J. E. Smith. En: Memorias seminario Spodoptera frugiperda (El gusano cogollero) en sorgo, maíz y otros cultivos. Cali, Colombia, p. 78-82. [ Links ]
LEZAMA, R. 1994. Patogenicidad de hongos parásitos de insectos. En: I Seminario Patología. PCBA. U de C. Tecomán, Colima. p. 47-69 . [ Links ]
MILNER, R.; NUPFATZ, R.; SWARIS, S. 1990. A new method for assessment of germination of Metarhizium conidia. Journal of Invertebrate Pathology Notes, p. 121. [ Links ]
PÉREZ, M. 2003. Control biológico de Spodoptera frugiperda Smith en maíz. Departamento de manejo de plagas, INISAV. [On line] Playa Ciudad de la Habana, Cuba. http://www.cied.org/bae/bae71/b71e.htm - Consulla Marzo 10, 2003. [ Links ]
RIBA, G.; GLANDARD, A. 1980. Mise au point d' un inilieu nutritif pour la culture profonde du champignon entomopathogene Nomuraea rileyi. Entomophaga 25 (3): 317-322. [ Links ]
St. LEGER, R.; CMAMLEY, A.; COOPER, R. 1986a. Cuticle - degrading enzymes of entomopathogenic fungi: synthesis in culture on cuticle. Journal of Invertebrate Pathology 48: 85-95. [ Links ]
St. LEGER, R.; CHAMLEY, A.; COOPER, R. 1986b. Cuticle - degrading enzymes of entomopa-thogenic fungi: mechanism of interaction between pathogen enzymes and ¡nsect cuticle. Journal of Invertebrate Pathology 47: 295-302. [ Links ]
St. LEGER, R.; CHAMLEY, A.; COOPER, R. 1986c. Cuticle degradation by fungal enzymes. Journal of Invertebrate Pathology 47:167-177. [ Links ]
VILLAMIZAR, L. P. 1998. Efecto de la composición del medio de cultivo en la virulencia de Metarhizium anisopliae sobre la langosta llanera Rhammatocerus schisto-cercoides. Santafé de Bogotá. Tesis (M. Se. Microbiología). Universidad Nacional de Colombia, p. 11 -37. [ Links ]
VILLAMIZAR. L. F.; COTES, A. M.; URIBE, D. 2001. Relación entre la actividad enzimática y la virulencia de rletharhizium anisopliae sobre la langosta llanera Rliammatocerus schistocercoides (Orthoptera: Acrididae) Revista Colombiana de Entomología 27 (3 - 4): 107-1 14. [ Links ]
VILLAMIZAR, L.; ARRIERO, C; COTES, A. 2004. Desarrollo de preformulados a base de nomuraea rileyi para el control de Spodoptera frugiperda. Revista Colombiana de Entomología 30 (1): 99-105. [ Links ]
VIMALA - DEVI. P S. 1994. Conidia production of the entomopathogenic fungus Nomuraea rileyi its evaluation for control of Spodoptera íiíura (Pab) on Ricinus communis. Journal Invertebrate Pathology 63: 145-150. [ Links ]
Recibido: Dic. 15/2003 Aceptado: Oct. 10/2004

