










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488On-line version ISSN 2665-4385
Rev. Colomb. Entomol. vol.31 no.2 Bogotá July/Dec. 2005
Tabla de vida de Delphastus pusillus (Coleoptera: Coccinellidae) en la mosca blanca Trialeurodes vaporariorum (Hemiptera: Aleyrodidae)
Life table of Delphastus pusillus (Coleoplera: Coccinellidae) on the whitefly Trialeurodes vaporariorum (Hemiptera: Aleyrodidae)
JAVIER GARCIA GONZALEZ1, EDGAR RICARDO BENITEZ S1, ARISTOBULO LOPEZ-AVILA2
1 Autor para Correspondencia. Ingeniero Agronomo M.Sc.Entomologia.Investigador cooperante, Area de Entomologia Programa Nacional de Manejo Integrado de Plagas MIP. Corpoica C.I. "Tibaitata". jgarcia@corpoica.org.co
2 Ph. D. Programa Nacional de Manejo Integrado de Plagas MIP. Corpoica C.I. "Tibaitata".
Resumen. Se construyo la tabla de vida del depredador Delphastus pusillus (Le Conte) (Coleoptera: Coccinellidae), alimentado sobre su presa la mosca blanca de los invernaderos Trialeurodes vaporariorum (Westwood) (Hemiptera: Aleyrodidae). Apartir de individuos procedentes de una colonia del depredador mantenida en laboratorio. se tomo un grupo de 30 adultos de edad conocida los cuales fueron ubicados en cajas de Petri. A partir de cohortes de 493 y 682 huevos del depredador,se realizaron observaciones de individuos vivos al inicio de cada estado, individuos muertos en cada estado, tiempo de duracion de cada estado, longevidad de la hembra. huevos puestos por hembra, tiempo en que la hembra oviposito y proporcion de sexos. Con la informacion colectada se estimaron los parametros tasa finita de crecimiento (A.), tasa intrinseca de crecimiento natural (rm). tasa neta reproductiva (R()), tiempo gcneracional (Tg), tiempo dc duplicacion de la poblacion original (Dt), tiempo de fecundidad de la hembra, tasa de oviposicion y se obtuvieron las curvas de oviposiciòn y sobre vivencia del depredador. Hubo menor sobrevivencia de D.pusillus en los primeros estados de desarrollo, particularmente en los estados de huevo y larval. Los parametros estimados para individuos de D. pusillus provenientes en una primera y una sexta generacion fueron: R(1 = 5.51 y 3.17 respectivamente, D( = 23.0 y 20.05, A, = 1,03 para las dos generaciones, r,„= 0.03 para las dos generaciones. Se plantea que hembras depredadoras de una primera generacion hacen un mayor aporte de hembras hijas, comparado con el aporte hecho por hembras de una generacion seis mantenida en laboratorio.
Palabras clave: Tasa intrinseca de crecimiento natural. Tiempo generacional. Curva de sobrevivencia.
Summary. The aim of this research was to construct the life table of the predator Delphastus pusillus (LeContc) (Coleoptera: Coccinellidae). feeding on its prey, the greenhouse whitefly Trialeurodes vaporariorum (Westwood) (Hemiptera: Aleyrodidae). A group of 30 adults of known age were taken from among individuals reared in a colony maintained in the laboratory, and put into Petri dishes. Based on cohorts of 493 and 682 eggs of the predator, observations were made on the number of individuals alive at the beginning of each development stage, individuals that died in each stage, duration of each stage, female longevity, eggs laid per female, period of oviposition and sex ratio. The information collected was used to calculate the parameters of finite growth rate (A.), intrinsic rate of natural increase (rm), net reproductive rate (R,.), generation time (Tg), time to population doubling (DO, time of female fecundity, oviposition rate, and the oviposition and survival curves of the predator. There was lower survival of D. pusillus in the early developmental stages, particularly the egg and larva I stages. The estimated parameters obtained from the first and sixth generations were: R<>= 5.51 and 3.17, respectively, Dt= 23.0 and 20.5, A. = 1.03 for both generations and rm = 0.03 for both generations. It is suggested that first generation female predators make a greater contribution to female daughters compared to the contribution made by females of the sixth generation maintained in laboratory.
Key words: Intrinsic natural growth rate. Generation time. Survivorship curve.
Introduccion
Las moscas blancas (Hemiptera: Aleyrodidae) se conocen de tiempo atras y han sido estudiadas por mas de 250 años. De las cerca de 1200 especies de mosca blanca reconocidas e identificadas a nivel mundial, Trialeurodes vaporariorum (Westwood) y Bemisia tahaci (Gennadius) difieren de las otras especies por los graves daños economicos que causan a la agricultura en muchas regiones del mundo (Gerling 2000). En Colombia T. vaporariorum es la especie predominante en el tropico alto y valles interandinos atacando principalmente cultivos de frijol, habichuela, tomate y papa (Quintero et al. 1999).
Como medidas de control de T. vaporariorum el uso de plaguicidas quimicos es casi la unica herramienta empleada. A partir de un diagnostico con agricultores se determino que el 46% de ellos han tenido que abandonar alguna vez el cultivo por mal control de la plaga, por lo que cuestionan el empleo de insecticidas. El numero de ingredientes activos usados contra moscas blanca asciende a 32, siendo los grupos insecticidas mas utilizados los organofosforados, carbamatos, piretroides y los de nueva generacion (Rendon et al. 2000).
En la busqueda de alternativas de manejo de la plaga, se han adelantado estudios con el insecto depredador Delphastus pusillus (Le Conte) (Coleoptera: Coccinellidae), el cual se alimenta de las dos especies de mosca blanca de mayor importancia economica T. vaporariorum y B. tabaci y se encuenlra ampliamente distribuido en el pais en los depatarmentos de Guajira, Cesar, Santander, Boyaca, Cundinamarca, Tolima, y Huila (Lopez-Avila et al. 2001).
Antes de liberar un enemigo natural en campo es necesario conocer algunos atributos biologicos y ecologicos que permitan identificar su efecto regulador. Uno de ellos es la capacidad de crecimiento de la poblacion del depredador con respecto a su presa (Rodriguez del Boque 1994). Algunas caracteristicas de la poblacion en estudio pueden ser obtenidas con la construccion de tablas de vida. La tabla de vida es un componente importante en el entendimiento de la dinamica de poblacion de una especie. Deevy (1947) citado por Soulhwood (1978), fue el primer investigador que enfoco su atencion en la importancia de estas tablas. Las tablas de vida han sido usadas para determinar la expectativa de vida de un candidato biologico y la expectativa de vida dc un estado especifico del organismo. Ya que muchos insectos tienen generaciones discretas y sus poblaciones no son estacionarias, las tablas de vida "de tipo horizontal o especificas por edades" son mas aplicables que las tablas de vida de "temporales o verticales". Las primeras se basan en la mortalidad real de una cohorte en la que los individuos pertenecen a la misma generacion, mientras que las tablas de vida temporales o verticales se basan en la muerte de una cohorte imaginaria encontrada por determinacion de estructura de edad en un punto especifico de desarrollo de la misma (Southwood 1978; Rabinovich 1980).
En la obtencion de una tabla de vida se deben estimar tres parametros de la poblacion para que esta sea lo mas completa posible. El primero es el numero de individuos vivos que entran a cada estado del ciclo de vida del insecto; el segundo la mortalidad a partir de factores especificos en cada estado de vida y el tercero la fecundidad (Bellows et al. 1992). Con el presente trabajo se estimaron parametros de la tabla de vida para Delphastus pusillus y se obtuvieron las curvas de sobrevivencia para el depredador criado en una primera y una sexta generacion mantenidas en laboratorio.
Materiales y Metodos
Con el fin de determinar diferencias en los parametros de la tabla de vida por efecto de las condiciones de cria en laboratorio, se establecieron diferencias para D. pusillus entre una generacion reciente y una generacion antigua, generaciones l y VI.
A partir de individuos procedentes de la colonia del depredador, se tomo un grupo de 30 adultos de edad conocida, los cuales fueron ubicados por parejas en cajas de Petri de vidrio de 5 cm de diametro. Al interior de las que se ubicaron foliolos infestados con inmaduros de mosca blanca; los foliolos fueron renovados cada dos dias, con el fin de retirar las posibles posturas asi como para renovar el alimento al depredador. Las cajas de petri donde se ubicaron las posturas de D. pusillus, fueron debidamente identificadas con la fecha de oviposicion y la pareja de adultos de la cual provenian. Con el fin de determinar el comportamiento biologico del depredador en condiciones de cria, se construyo la tabla de vida para una primera y una sexta generacion; a partir de una cohorte de 493 y 682 huevos del depredador respectivamente. Se realizaron las observaciones en los diferentes estados por los que pasa el insecto. Se registro el numero de individuos vivos al inicio de cada estado, individuos muertos en cada estado, tiempo de duracion de cada estado, longevidad de la hembra, huevos puestos por hembra, tiempo en que la hembra fue fecunda y proporcion de sexos.
Con estos registros se estimaron los siguientes parametros de fecundidad del depredador: tiempo de fecundidad de la hembra definido por el tiempo que la hembra oviposito dividido por su longevidad, tasa de crecimiento reproductivo (GRR), tasa de oviposicion promedio dia, tasa neta reproductiva (R()), tasa intrinseca de crecimiento natural (rm) y tasa finita de crecimiento (A). Posteriormente se obtuvieron parametros adicionales no relacionados con la fecundidad de D. pusillus: tiempo generacional (Tg), tiempo de duplicacion de la poblacion inicial (Dt) y longevidad de la hembra. Finalmente se determino el comportamiento de sobrevivencia y mortalidad de las poblaciones del depredador en estudio.
La informacion se analizo con ayuda del procedimiento propuesto por Maia et al. (2000) y el paquete estadistico SAS® (1999). La comparacion de los parametros estimados para las dos generaciones en estudio se realizo con la prueba t de una cola.
Resultados y discusion
La prueba t de una cola presento diferencias significativas entre las generaciones uno y seis para las variables tiempo de fecundidad, tasa de crecimiento reproductivo (GRR), tasa neta reproductiva (R()) y tasa de oviposicion hembra/dia (Tabla 1).
El tiempo de fecundidad, definido como el tiempo de vida en que la hembra oviposito dividido por su longevidad, para hembras de D. pusillus en la primera generacion fue de 69,3 dias. En este tiempo, la hembra depredadora presento una tasa de crecimiento reproductivo (GRR) de 185,3 huevos y una tasa de oviposicion diaria de 0,84 (Tabla 1). Las hembras en la sexta generacion mostraron un tiempo de fecundidad de 49,0 dias, una GRR de 84,2 huevos y una tasa de oviposicion hembra/dia de 0,61 (Tabla 1).Para la variable tasa neta reproductiva (R()) se obtuvo un valor estimado de 5,51 hembras hijas/hembra madre para la primera generacion; para la generacion seis el aporte de hembras hijas/hembra madre fue de 3,17. Los resultados muestran que hembras del depredador en la primera generacion hacen un mayor aporte de hembras hijas respecto al aporte hecho por hembras de una generacion seis mantenida en laboratorios.
El significado del valor de R()en ecologia de poblaciones, señala el crecimiento, disminuciòn o estabilidad de una poblacion de generacion en generacion; valores de R() mayores de la unidad indican que la poblacion en estudio se encuentra en estado de crecimiento, caso observado para la poblacion de D. pusillus, donde dichos valores son mayores que la unidad (Southwood 1978; Rabinovich 1980).
La cria de insectos en condiciones de laboratorio puede generar adaptaciones no geneticas temporales, o cambios geneticos permanentes como resultado de seleccion, deriva y endocria. Ambos tipos de cambios pueden influenciar el comportamiento del insecto benefico al ser liberado en campo y puede afectar su desempeño en programas de control biologico. Por ello la implementacion de un esquema de control de calidad es importante en un programa de manejo de plagas cuando se utilizan enemigos naturales producidos en laboratorio (De Clercq et al. 1998). A partir de la informacion obtenida se hace necesario determinar a partir de que generacion del depredador hay mayor reduccion del valor R(), para introducir poblaciones de D. pusillus de campo y mitigar la disminucion de hembras hijas.
La prueba t no delecto diferencias significativas para las variables tasa intriseca de crecimiento (rm) y tasa finita de crecimiento (A), en las generaciones de D. pusillus evaluadas. La variable tasa intrinseca de crecimiento (r,()), para las dos generaciones presento un valor de 0,03 hembras/hembra/dia; mientras que para la variable tasa finita de crecimiento (k) el valor fue 1,03.
La igualdad en los paramelros rm y A estimados para las dos generaciones en estudio se podria explicar por que estos son considerados intrinsecos dc la especie y estan regidos por factores geneticos. Al respecto Rabinovich (1980), menciona que rm, como una tasa intrinseca de crecimiento natural, es un parametro geneticamente determinado y refleja una capacidad potencial de multiplicacion de la poblacion.
En condiciones naturales los factores que afectan el equilibrio de una poblacion estan dados por el nacimiento, muerte. inmigracion y emigracion de los individuos. En condiciones controladas los dos ultimos factores no son considerados. En condiciones controladas el parametro rm esta regido por la natalidad y la mortalidad de individuos de la poblacion, las cuales para las condiciones de estudio fueron similares.
En terminos demograficos el valor de A esta determinado por el ambiente y por el ciclo de vida de las especies. Este parametro caracteriza la dinamica potencial de una poblacion de una forma simplificada y sirve para discutir efectos de escenarios mas complejos. Ademas sirve como punto de partida de manejo de poblaciones ya sea para reducir la diseminacion de la plaga o para incrementar la dispersion de una especie introducida (Ncuberty Caswell 2000).
En la mayoria de las hembras evaluadas se observo que la primera postura no fue viable, pero fue de importancia para ubicar sitios de oviposicion seguros; algunos sitios usados por la hembra de D. pusillus para proteger sus posturas fueron las nervaduras de las hojas, debajo del cuerpo de adultos de mosca blanca muertos y en el interior de exuvias pupales de mosca blanca; al parecer, este comportamiento reduce la probabilidad de que los huevos sean atacados por otros enemigos naturales.
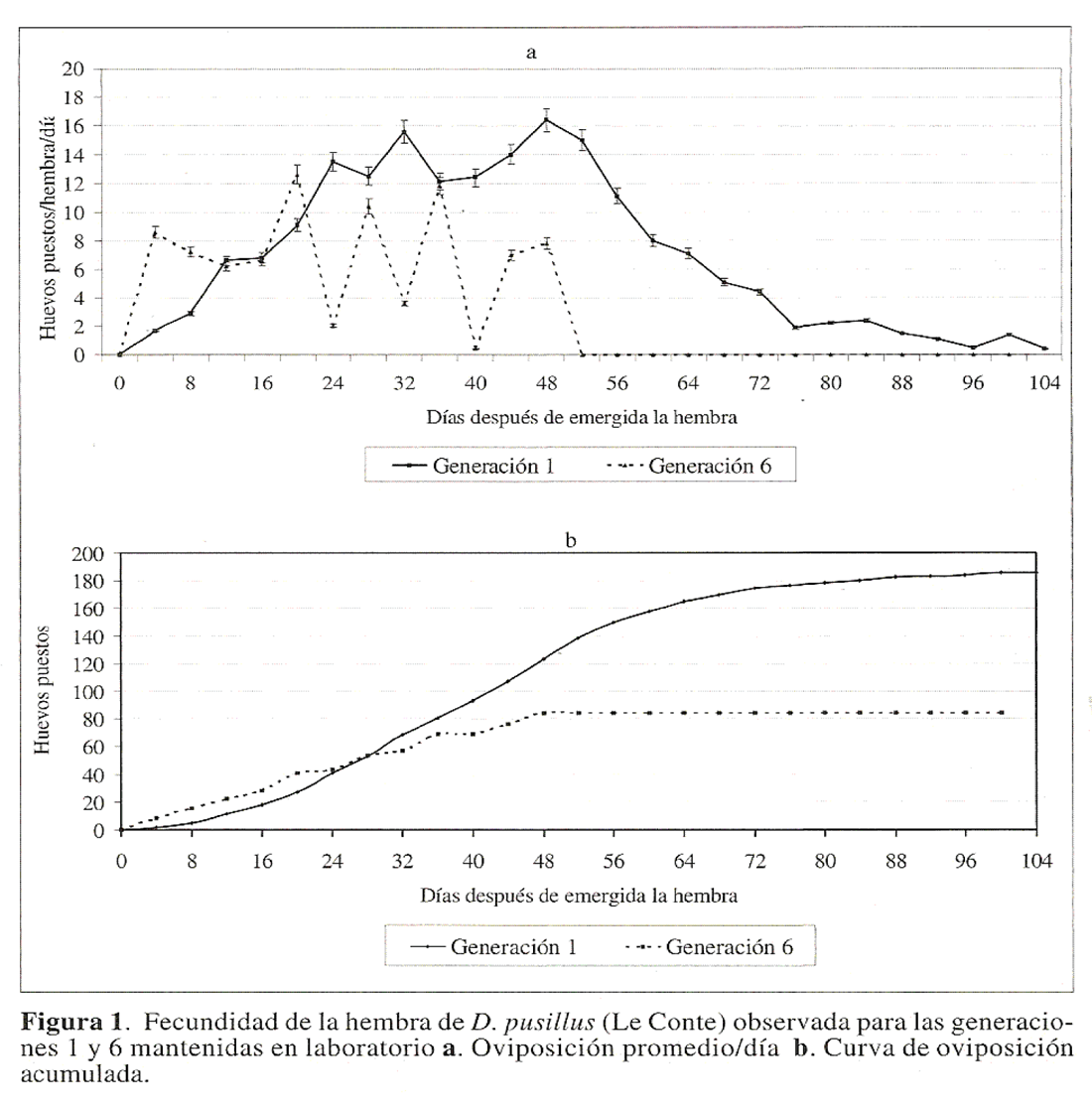
La curva de oviposicion hembra/dia (Fig. 1a), para hembras en la primera generacion presento dos picos. El primero a los 32 dias y el segundo a los 48 dias de vida de la hembra con un promedio de 15,6 y 16,4 huevos/hembra/dia respectivamente. Despues del dia 48 disminuyo la cantidad de huevos puestos, hasta ser cero al ultimo dia de vida de la hembra (dia 104)
La curva de oviposicion acumulada para hembras de la primera generacion mostro una tendencia sigmoide. Desde el dia cero hasta el dia 24 de vida de la hembra, se observo un leve incremento de oviposicion. Entre los dias 30 y 68 de vida de la hembra se presento un incremento acelerado de oviposicion (Fig. 1b), determinado por los mayores picos de oviposicion. Despues del dia 68 de emergida la hembra, se redujo la tasa de oviposicion que concuerda con los periodos finales de vida de la hembra (Fig. 1b).
La curva de huevos puestos/hembra/dia para hembras de D. pusillus en una sexta generacion mostro picos de alta oviposicion seguidos por perfodos de baja oviposicion (Fig. 1a). Este comportamiento junto al hecho de que la hembra presento un bajo tiempo de fecundidad respecto a hembras de la primera generacion, redujo en gran medida la GRR para la sexta generacion (Tabla 1). La curva de oviposicion acumulada para hembras de la sexta generacion mostro un incremento en la cantidad de huevos puestos desde el dia 4 hasta el dia 48 de vida de la hembra. Despues de esta fecha no se observaron posturas, por lo que la curva se eslabilizo hasta la muerte de la ultima hembra producida al dia 100 (Fig. 1b).
A partir de las curvas de oviposicion acumulada se evidencia la diferencia que existe en la fecundidad de hembras de la primera y la sexta generacion de D. pusillus. La oviposicion total acumulada para hembras de la primera y la sexta generacion respectivamente fue de 185,3 y 84,2 con un promedio de 1,8 y 0,8 huevos puestos/hembra/dia respectivamente. El periodo de finalizacion de la oviposicion, representado por el tiempo en que la curva se. estabiliza, se observo en los dias 84 y 48 de emergida la hembra para las generaciones uno y seis respectivamente (Fig. 1b).Algunos de los factores descritos como limitantes de la fecundidad de especies mantenidas en laboratorio son la calidad y cantidad del esporma producido por el macho (Jervis y Copland 1996). Otro factor limitante es la endocria, en la que algunas caracteristicas biologicas de la especie como la fecundidad y la proporcion de sexos se afectan negativamente, al cruzar individuos con algun parentesco (De Clercq et al. 1998).
El procedimiento seguido para calcular parametros de la tabla de vida permitio obtener estimativos para otras variables diferentes a la fecundidad y que correspondieron a longevidad. tiempo generacional (Tg) y tiempo de duplicacion de la poblacion original (Dt).
La prueba t mostro diferencias significativas para el tiempo de duplicacion de la poblacion original, pero no evidencio diferencias para el tiempo generacional y longevidad de la hembra, en las generaciones evaluadas (Tabla 2). De acuerdo con los parametros estimados D. pusillus en una primera generacion requiere 23,0 dias para duplicar su poblacion inicial; mientras que el depredador en la generacion seis, requiere 20,5 dias (Tabla 2). Es probable que D. pusillus trate de remediar la reduccion en la fecundidad con la disminucion en el tiempo necesario para duplicar la poblacion original. La variable longevidad en las hembras de D. pusillus fue de 82,5 dias para hembras en la primera generacion. Para hembras de la sexta generacion la longevidad de la hembra fue de 79,8 dias en promedio (Tabla 2).
De los parametros estimados, el tiempo de duplicacion de la poblacion inicial (Dt) fue el unico que se vio afectado por las crias sucesivas en laboratorio. Posiblemente existe una relacion directa entre (Dt) con los parametros de fecundidad tasa neta reproductiva (R()), tiempo de fecundidad, tasa de crecimiento reproductivo (GRR) y tasa de oviposicion hembra/dia, los cuales fueron significativamente diferentes en las dos generaciones evaluadas (Tabla 1). Como lo muestran los datos, la cria sucesiva del depredador en laboratorio reduce algunos componentes de fecundidad por lo que se debe determinar la generacion a partir de la cual se reduce de manera significativa la fecundidad para introducir poblaciones del depredador en campo.Algunas investigaciones han mostrado la efectividad de la cria continuada de insectos en laboratorio. Cohen y colaboradores (1999), demostraron casos de crias continuadas de insectos en laboratorio por mas de 25 años. Se cita el ejemplo del insecto depredador Geocorys punctipes (Hemiptera: Lygaeidae) depredador generalista con el cual se ha mantenido una cria continuada por mas de 150 generaciones y la colonia se ha llevado por cerca de 15 años en laboratorio. Para que esta colonia se estabilizara en el tiempo, fue necesario realizar una seleccion de individuos que permitiera obtener aquellos con capacidades de sobrevivencia en laboratorio. Una vez superado este paso, la colonia del depredador se ha mantenido.
Al analizar la proporcion de individuos vivos de D. pusillus durante su tiempo de vida, los resultados mostraron una reduccion significativa desde los estados iniciales de desarrollo del insecto; la disminucion en la sobrevivencia fue mas acelerada en individuos de la sexta generacion del depredador respecto a aquellos obtenidos en la primera generacion (Fig. 2).
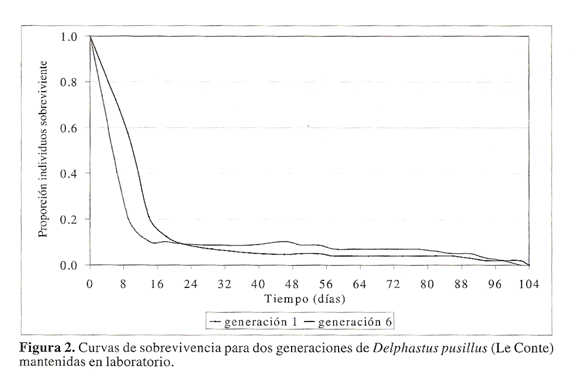
En la primera generacion se observo una reduccion en la proporcion de individuos vivos al pasar de 1,0 a 0.13 desde el dia cero hasta el dia 18 de puesto el huevo. Despues del dia 18 y hasta el dia 104 la probabilidad de individuos vivos presento disminuciones minimas con 0,13 individuos vivos al dia 18. con 0,09 individuos vivos al dia 22, con 0,05 al dia 54, con 0,04 al dia 86 y con 0,02 sobrevivientes al dia 102 (Fig. 2). Igual tendencia se observo con individuos de la generacion seis, aunque la reduccion en la probabilidad de sobrevivientes fue mas acelerada al inicio del tiempo de vida de los individuos. En los primeros 14 dias, la probabilidad de vida del depredador se redujo de 1,0 a 0,10. Despues del dia 14 y hasta el dia 54, la probabilidad de vida del depredador se redujo de 0,1 a 0,09 individuos vivos. Desde el dia 54 al dia 78 la probabilidad de vida se redujo de 0,09 a 0,07. A partir de esta fecha la probabilidad de vida se redujo hasta ser cero al ultimo dia de vida del adulto.
Los resultados de sobrevivencia de los diferentes estados de D. pusillus, mostraron una reduccion importante durante los primeros dias de vida del depredador. Este periodo coincidio con los estados de huevo y larval. Individuos del depredador en una primera generacion tienen mayor probabilidad de sobrevivir en los primeros estados de desarrollo del insecto comparado con aquellos de una sexta generacion. Individuos de la primera generacion tuvieron una probabilidad de vida de 0,21 al dia 14; mientras que individuos de una sexta generacion tuvieron una probabilidad de vida de 0,10.
La reduccion de la sobrevivencia en los primeros dias de vida de D. pusillus, se debio a la cantidad de huevos que no fueron fertilizados. Otro factor de mortalidad observado en el estado de huevo se debio en menor grado a huevos depredados por larvas del mismo depredador. Cuando ocurrieron oviposiciones en grupo, las larvas que primero emergieron tuvieron la posibilidad de consumir huevos de su especie.
En el estado de larva 1 las causas de mortalidad observadas fueron intrinsecas y extrinsecas. Aquellas larvas que no tienen alimento en su vecindad salen en busqueda de presas. Si en el transcurso de 20 a 30 minutos la larva no encuentra alimento, muere. En estos casos la caracteristica de la hoja juega un factor esencial. En el caso de foliolos con gran cantidad de tricomas se interfiere el proceso de busqueda y encuentro de presas por el depredador ya que se dificulta el contacto entre depredador y presa. Estudios para determinar el efecto de la cantidad de tricomas presentes en la superficie de la hoja sobre el comportamiento reproductivo y alimenticio de D. pusillus, han demostrado que el depredador presenta mayor fecundidad y consumo de ninfas de mosca blanca cuando la hoja posee baja cantidad de tricomas (Heinz et al. 1994).
Aquellos individuos que superaron los estados mencionados tuvieron mayor probabilidad de vida. lo que se evidencio con la estabilidad de las curvas de sobrevivencia (Fig. 2). Despues del dia 22 de vida y hasta la muerte del ultimo adulto al dia 104, individuos de la primera generacion presentaron menor probabilidad de vida respecto a la probabilidad observada en individuos de la sexta generacion. Este resultado se tradujo en una menor sobrevivencia de los estados de larva2, larva3, larva4, pupa y adulto para individuos de la primera generacion.
De acuerdo con los principales tipos de curvas de sobrevivencia propuestos en la literatura D. pusillus asemeja una curva tipo IV, (Fig. 3). Una curva de sobrevivencia tipo IV se caracteriza por que la mortalidad natural afecta fundamentalmente a los individuos jovenes de la poblacion.
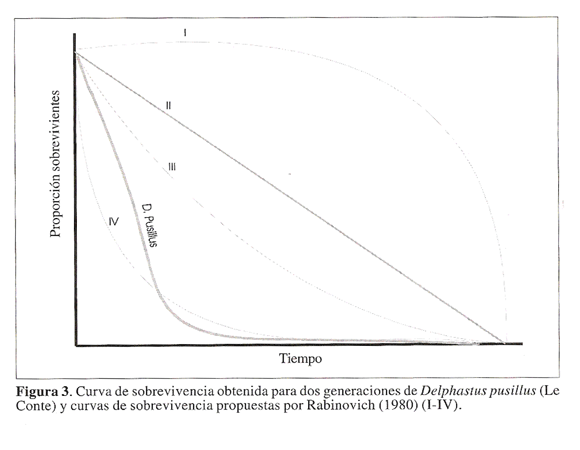
Cuando se han superado las etapas juveniles, la mortalidad natural se reduce en forma considerable y la sobrevivencia de los individuos de la poblacion se estabiliza (Rabinovich 1980; Southwood 1978). En condiciones de cria se hace necesario suministrar suficiente alimento y de buena calidad nutricional al depredador en los primeros instares larvales y facilitar la consecucion de presas sanas para la larva recien emergida con el fin de incrementar la probabilidad de sobrevivencia de estos estados.
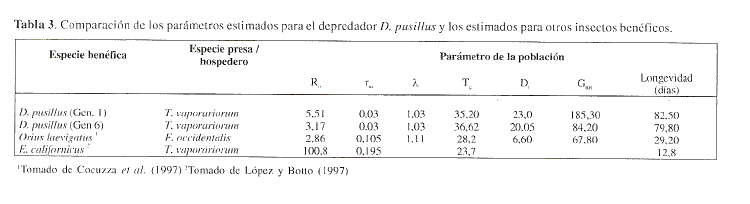
Al comparar los parametros estimados para el depredador D. pusillus con los registros obtenidos para otros insectos entomofagos se puede tener una idea del comportamiento del enemigo natural sobre la especie hospedera/presa. Se compararon los parametros de poblacion de D. pusillus con los registrados en la literatura para los insectos Orius laevigatus (Hemiptera: Anthocoridae) y Eretmocerus californicus (Hymenoptera: Aphelinidae), respectivamente un insecto depredador y un insecto parasitoide (Cocuzza et al. 1997; Lopez y Botlo 1997). Los parametros tasa intrinseca de crecimiento rm, tasa finita de crecimiento A y tiempo generational Tg. fueron menores para el depredador en estudio D. pusillus respecto a los enemigos naturales mencionados. Para las variables longevidad. tasa reproductiva de crecimiento (GRR), tiempo de duplicacion (Dt y tasa neta reproductiva R(), D. pusillus presento valores mayores comparado con O. laevigatus (Tabla 3). Como lo muestran los parametros estimados y presentados en la Tabla 3, insectos depredadores como D. pusillus presentan valores de R(), rm y A bajos comparados con los observados por insectos parasitoides, caso especifico E. californicus. Estos resultados se explican en el hecho quc los parametros mencionados se fundamcntan en la produccion de hembras hijas por unidad de tiempo o por generacion. Los insectos depredadores requieren del consumo de una cierta cantidad de presas para poder alcanzar su estado adulto y asi producir, de acuerdo a la proporcion de sexos, una hembra (Hassell 1980).
El hecho de requerir mas de una presa para desarrollarse, incrementa el riesgo de mortalidad del depredador en los diferentes estados inmaduros. El efecto biocontrolador de los depredadores se fundamenta en el hecho que tanto larvas como adultos del depredador requieren consumir mas de una presa para cumplir con sus funciones vitales. En el caso de la larva para completar su ciclo de vida, mientras que el adulto lo utiliza para funciones reproductivas. Caso contrario ocurre en los parasitoides, dondc la relacion parasitoide/huesped es por lo menos 1:1. es decir por cada huesped parasitado se asegura el alimento al inmaduro y la formacion de minimo un adulto (Hassell 1980).
Conclusiones
De los resultados obtenidos en los diferentes experimentos se puede concluir que:
- Los primeros estados de desarrollo de Delphastus pusillus (Le Conte) son los mas sensibles a los factores de mortalidad natural. Cuando se superan dichos estados la mortalidad de la poblacion tiende a equilibrarse.
- Caractcristicas intrinsecas del depredador reflejadas por los valores similares de rm y A en las dos generaciones evaluadas permiten proponer la cria continuada en laboratorio del depredador.
- Hembras de una primera generacion respecto a hembras de una sexta generacion hacen un mayor aporte a la poblacion de hembras hijas.
- La ubicacion de posturas por la hembra de D. pusillus en areas con disponibilidad de presas sanas, incrementa la probabilidad de sobrevivencia de las larvas recien emergidas.
Literatura citada
BELLOWS JR TS. VAN DRISCHE RG, ELKINTON JS 1992. Life-Table construction and analysis in the evaluation of natural enemies. Annual Review of Entomology. 37: 587-614. [ Links ]
COCUZZA GE: DE CLERG P; LIZZIO S; VAN DE VIERE M; TIRRY L; DE-GHEELE D: VACANTE V 1997. Life tables and predation activity of Orius laevigatus and O. albidipennis at three constant temperatures. Entomologia Experimentaliset Applicala. 85: 189-197. [ Links ]
COHEN C.A.; NORDLUND D; SMITH R. 1999. Mass rearing of entomophagous insects and prcdaceous mites: are the bottlenecks biological, engineering, economic, or cultural? Biocontrol, news and information. 20 (3): 85-90. [ Links ]
DE CLERCG P; VANDEWALLE M-:. L. TIRRY. 1998. Impact of inbreeding on periormance of the predator Podisus maculiventris. Biocontrol. Netherlands. 43. 299-310. [ Links ]
GERLING D. 2000. Whiteflies revisited. XXI international congress of entomology. Abstract book. Brazil. August 20 -26. pp. 64-67. [ Links ]
HASSELL M.P. 1980. The Dynamics of Competition and Predation. Department of Zoology and Applied Entomology. Imperial College of Science and Technology. Study in Biology No. 72. 68. [ Links ]
HEINZ K.; BRAZZLE J.; PICKETT CH.: NATWICK E.; NELSON J.: PARR ELLA M. 1994. Predatory Beetle may suppress silverleafwhitefly. California Agriculture. (United States) 48 (2): 35-41. [ Links ]
JERVIS M. A.: COPLAND M. J. 1996. The life cycle. Insect Natural enemies. Practical approaches to their study. ED. Chapman and Hall, 63-160. [ Links ]
LOPEZ-AVILA A.: CARDONA MEJIA C: GARCIA GONZALEZ J.: RENDON F.; HERNANDEZ P. 2001. Reconocimiento e identificacion de enemigos naturales de
moscas blancas (Homoptera: Aleyrodidae) en Colombia y Ecuador. Revista Colombiana de Entomologia. 27(3-4): 137-141. [ Links ]
LOPEZS.N.; BOTTON.E. 1997. Biology of South American population of Eret-mocerussp. (Hymenoptera: Aphclinidae) attacking the greenhouse withefly. Biological control. 9(1): 1-5. [ Links ]
MAIA A.; LUIZ A.; CLAYTON C. 2000. Statistical inference on associated fertility table parameters using Jackknife technique: computational aspects. Journal of Economic Entomology. 93 (2): 511-518. [ Links ]
NEUBERT M.; CASWELL H. 2000. Demography and dispersal: Calculation and sensitivity analysis of invasion speed for structured populations. Ecology 8 (16): 1613-1628. [ Links ]
QUINTERO C; RENDON F.: GARCIA J.; CARDONA C: LOPEZ-AVILA A.: HERNANDEZ P. 1999 Especies y biotipos de moscas blancas (Homoptera: Aleyrodidae) en cultivos semestrales de Colombia y Ecuador. Revista Colombiana de Entomologia. 27 (1 -2): 27-32. [ Links ]
RABINOVICH J. E. 1980. Introduccion a la ecologia de poblaciones animales. Capitulo 5. Editorial CECSA. Caracas (Venezuela). 313. [ Links ]
RRENDON F; GARCIA J.: VALERAZO O.: CARDONA C; LOPEZ-AVILA A.: BUENO J.: RAMIREZ J. 1999. La problemalica de las moscas blancas (Homoptera: Aleyrodidae) en cultivos anuales en el tropico alto, valles interandinos y costas dc Colombia y Ecuador: 1. Caracterizacion y diagnostico general. Resumenes XXVI Congreso de la Sociedad Colombiana de Entomologia. Santafe de Bogota (Colombia), 78. [ Links ]
RODRIGUEZ DEL BOSQUE, L. 1994. Teorias y bases ecologicas del control biologico. Memorias V curso de control biologico. Institute tecnologico agropecuario Oaxaca. Mexico, 6-15. [ Links ]
SAS INSTITUTE 1999. Sas Language. Reference. Version 8. Sas institute. Gary NC SOUTHWOOD T.R. 1978. Ecological Methods with particular reference to the study of insect population. Ed. Chapman and Hall. London. [ Links ] [ Links ]
Recibido: 08-oct-03 - Aceplado: 01-jun-05


{kind=link}
{kind=link}
{kind=link}
{kind=link}