










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488On-line version ISSN 2665-4385
Rev. Colomb. Entomol. vol.32 no.1 Bogotá Jan./June 2006
Mediación química entre Allomerus demerarae Wheeler, 1935
(Hymenoptera: Formicidae) y Cordia nodosa Lam. (Boraginaceae)
Chemical mediation between Allomerus demerarae Wheeler, 1935
(Hymenoptera: Formicidae) and Cordia nodosa Lam. (Boraginaceae)
ROXANA ARAUCO A.1,2, JOSE IANNACONE O.1,3, DAVID EDWARDS5, DOUGLAS YU4
1 Laboratorio de Ecofisiología Animal. Facultad de Ciencias Naturales y Matemáticas. Universidad Nacional Federico Villarreal. Calle San Marcos 383, Pueblo Libre, Lima, Perú.
2 rp_arauco@yahoo.com
3 Autor para correspondencia. Facultad de Ciencias Biológicas. Universidad Ricardo Palma. E-mail: joseiannacone@yahoo.es ; Teléfono: ++ 012573227. Telefax: ++ 014600930.
4 Center for Ecology, Evolution and Conservation. University of East Anglia, Norwich. Norfolk. UK.
5 School of Biological Sciences. University of East Anglia, Norwich. Norfolk. UK.
Resumen. Se investigaron las interacciones químicas entre hormigas obreras patrulladoras y de domacio de Allomerus demerarae Wheeler, 1935 y la planta mirmecofila Cordia nodosa Lam. Se determinaron diferencias entre la intensidad de atracción de hormigas hacia los compuestos volátiles de cuatro estímulos químicos: extractos de hojas jóvenes, de hojas maduras, de inflorescencia de C. nodosa, y de huevos-larvas de A. demerarae. En el año 2002 se coleccionaron plantas y hormigas en el albergue turístico Libertador Jungle Lodge ubicado en la zona de amortiguamiento de la Reserva Nacional de Tambopata, Madre de Dios, Perú. En esta simbiosis se ha evidenciado la medición química para hacer efectivo el comportamiento de A. demerarae. Los resultados mostraron que los compuestos volátiles emitidos por las hojas jóvenes son suficientes para desencadenar el comportamiento de patrullaje protectivo en las obreras de A. demerarae. En cambio, las hormigas obreras de domacio mostraron una atracción mayor hacia los huevos-larvas de A. demerarae y las flores de C. nodosa. Los resultados sugieren la existencia de polietismo en las colonias de A. demerarae en las que se han identificado dos grupos de obreras con funciones diferentes. Con base en estos resultados se sugieren dos subcastas; las nodrizas las cuales se encuentran hacia el interior del domacio y son las encargadas del comportamiento de castración de las flores de C. nodosa, y las patrulladoras las cuales fueron observadas sobre las hojas jóvenes.
Palabras clave: Comportamiento de patrullaje, comunicación química, mutualismo, nodriza, parasitismo.
Abstract. The chemical interactions between patrolling and domatia workers of Allomerus demerarae Wheeler, 1935 and the plant myrmecophylic Cordia nodosa Lam were investigated. Differences between the intensity of attraction of ants toward the volatile compounds of four chemical stimuli were determined: extracts of young leaves, of mature leaves, of C. nodosa inflorescences and of A. demerarae eggs-larvae. In 2002, plants and ants were collected at the tourist lodge Libertador Jungle Lodge located in the buffer zone of the Tambopata National Reserve, Madre de Dios, Peru. In this symbiosis, chemical mediation has been demonstrated to make the behavior of A. demerarae effective. The results showed that volatile compounds emitted by young leaves are sufficient to prompt the protective patrolling behavior of A. demerarae workers. In contrast, domatia worker ants showed a greater attraction to A. demerarae eggs-larvae and C. nodosa flowers. Results suggest the existence of polyethism in A. demerarae colonies in which two worker groups with different functions have been identified. On the basis of these results, two subcastes are suggested, nursery ants, which are found inside domatia and are in charge of the behavior of castration of C. nodosa flowers, and patrolling ants, which were observed on young leaves.
Keywords: chemical communication. Mutualism. Nursery. Patrolling behavior. Parasitism.
Introducción
En comparación con cualquier otro ecosistema terrestre los bosques tropicales lluviosos son poseedores de la mayor diversidad de especies (Kricher 1997; du Toit et al. 2004). Además de la diversidad de especies, estos ecosistemas son diversos en las interacciones que los organismos establecen entre ellos para sobrevivir (Thompson 1982; 1999; Chapin et al. 2000; Loreau et al. 2001; Heil y McKey 2003). La búsqueda de los factores y principios que gobiernan el mantenimiento y rompimiento de las interacciones entre especies, sobre todo, de los sistemas de cooperación, es aún un problema intrigante y persistente para la ecología evolutiva (Yu y Pierce 1998; Chapman 2003; Dejean et al. 2004). Aunque existen numerosas referencias en lo que concierne a las interacciones entre plantas y animales aún no es posible tener una visión de los principios que regulan estas interacciones debido a su gran diversidad taxonómica y ecológica (Janzen 1966; Davidson et al. 1990; Bronstein 1998; Davidson y McKey 1993; Hoechberg et al. 2000; Thompson y Cunningham 2002; Gomulkiewickz et al. 2003; Solano y Dejean 2004; Frederickson 2005).
Las interacciones planta – hormiga son sistemas de cooperación importantes para estudios que buscan encontrar los principios que regulan las interacciones interespecificas, por tres razones fundamentales: 1) porque el número de asociaciones simbióticas de este tipo es suficientemente pequeño como para resumirse en una sola tabla y a la vez lo suficientemente grande para probar estadísticamente determinadas hipótesis;2) porque su presencia en regiones tropicales alrededor del mundo facilita las comparaciones entre equivalentes taxonómicos y ecológicos que evolucionaron aisladamente en diferentes continentes; y,3) porque a pesar de su considerable diversidad y amplia distribución, estas relaciones son relativamente uniformes en estructura (Davidson y McKey 1993; Hoechberg et al. 2000; Gaume et al. 2005).
La interacción planta - hormiga más importante es la que se da con las plantas mirmecofilas. Por definición, una verdadera mirmecofila es aquella planta que se ha adaptado a la presencia de las hormigas, modificando alguna de sus estructuras vegetativas, tales como tallos, estípulas foliares, pecíolo, etc., en domacios (del latín domatia que significa «casa pequeña») donde la colonia pueda desarrollarse (Davidson y McKey 1993). En la mayoría de los casos, tanto hormiga como planta no pueden establecerse, sobrevivir, ni reproducirse en ausencia del otro. La remoción de la colonia residente de hormigas conllevaría a un aumento del herbivorismo y/o la disminución del crecimiento, fecundidad y supervivencia de la planta (Janzen 1966; McKey 1984; Gaume et al. 2005).
Las interacciones planta – hormiga en la que la señal que desencadena el comportamiento de patrullaje no es la producción de nectarios florales, han llamado la atención de los científicos hacia la comunicación química interespecífica, aunque las observaciones sugieren que los compuestos químicos de las plantas juegan un rol importante en el funcionamiento de las interacciones específicas. La ecología química de las simbiosis planta - hormiga está virtualmente inexplorada (Brouat et al. 2000; Dufay et al. 2003). Existen algunos trabajos sobre el papel que desempeñan los compuestos químicos de determinadas plantas para atraer a hormigas específicas que dispersen y siembren sus semillas, pero los compuestos responsables de esta atracción aún no han sido bien identificados (Davidson et al. 1990).
Empleando bioensayos, Brouat et al. (2000), en el sistema Leonardoxa afri-caca (Baill) Aubrev– Petalomyrmex phylax Snelling, 1979 (Hymenoptera: Formicidae), han proporcionado las primeras evidencias sobre el comportamiento de patrullaje de hojas jóvenes en interacciones planta - hormiga en las que se descartan los nectarios florales como inductores del comportamiento de protección, y que éste es mediado químicamente por compuestos químicos presentes en las hojas jóvenes (McKey 1984).
Uno de los aspectos más interesantes para el éxito de una colonia de insectos sociales es su organización en castas (Wilson 1980; Holldöbler y Wilson 1990; Hoyt 1998). En el caso de las hormigas, las castas son: la casta reproductora compuesta por la reina y los zánganos; la casta no reproductora constituida por las obreras. La casta obrera de las hormigas se subdivide en grupos especializados en realizar determinadas labores, es decir se caracteriza por presentar polietismo (de los términos en latin poly y ethos que significan varios comportamientos) (Holl-döbler y Wilson 1990; Sendova y Franks 1999). Se sugiere que este polietismo puede ser de dos tipos: físico y temporal. En el caso de colonias organizadas por polietismo físico, las castas son morfológicamente distintas (polimórficas), siendo normalmente de diferentes tamaños. En los casos de polietismo temporal, los individuos llevan a cabo diferentes tareas según la edad que tengan (Hölldobler y Wilson 1990; F´en´eron et al. 1996; Hughes y Goulson 2001). Se han llevado a cabo numerosos trabajos sobre la división de labores al interior de la colonia, pero aún existen muchas interrogantes sobre los principios que la regulan (Gautrais et al. 2002).
La mirmecofila Cordia nodosa Lam. (Boraginaceae) es un arbusto o árbol ampliamente distribuido en los bosques tropicales de Madre de Dios, en Perú, y más ampliamente a través de Sudamérica. Esta mirmecófila vive en simbiosis con Allomerus demerarae Wheeler, 1935 (Hymenoptera: Formicidae: Myrmi-cinae), Azteca ulei var. cordiae Forel, 1904 (Hymenoptera: Formicidae: Doli-choderinae), y cuatro especies de Azteca aún no descritas. Una planta individual aloja sólo a una de estas seis especies (Yu y Pierce 1998). La hormiga consigue protección hacia el interior de los domacios los cuales son ensanchamientos a nivel de las ramas donde la planta se beneficia del patrullaje protectivo de sus hojas jóvenes por parte de las hormigas (Yu y Wilson 2001). Cuando C. nodosa se asocia con A. demerarae la interacción es parasítica ya que A. demerarae no permite el desarrollo de frutos ni semillas en las plantas de C. nodosa.
Yu (1997) encontró que las colonias de A. demerarae se beneficiaban de este comportamiento de castración de las plantas de C. nodosa, puesto que la planta canalizaba su energía a la producción de más tejido vegetativo, que obviamente implicaba la producción de más doma-cios. Aun hay muchos estudios por hacer para ver con claridad el proceso de evolución de esta mirmecofila.
En Cordia nodosa no se ha establecido claramente si solo los nectarios u otro tipo de cuerpos alimenticios atraen las hormigas hacia las hojas jóvenes (Solano et al. 2005). Frederickson (2001) ha proporcionado evidencia experimental de que el comportamiento de patrullaje de las hormigas en este sistema también es mediado químicamente. Sin embargo, el uso de técnicas como headspace permitirá determinar si los compuestos naturalmente emitidos por las hojas jóvenes son suficientes para desencadenar el comportamiento de patrullaje. Para el caso de las colonias de A. demerarae no existe evidencia de diferencias morfológicas entre los miembros de la casta obrera, ni se han realizados estudios sobre la organización social al interior de esta colonia. Por esta razón, este estudio tiene por finalidad contribuir a la comprensión del funcionamiento de las interacciones planta-hormiga, explorando la comunicación química entre la colonia de hormigas de A. demerarae y la mirmecofila C. nodosa, para lo cual se determinó si existen diferencias significativas entre la intensidad de atracción hacia los compuestos volátiles de hojas jóvenes y hacia compuestos volátiles de hojas maduras por parte de hormigas de A. demerarae, y si existen diferencian significativas en el patrón de atracción de obreras patrulladoras y el de las obreras de domacio frente a cuatro estímulos químicos (extractos de hojas jóvenes, de hojas maduras, de inflorescencia de C. nodosa y de huevos-larvas de A. demerarae)
Materiales y Métodos
Sitio de estudio. El trabajo se realizó en los alrededores del albergue turístico Libertador Jungle Lodge, ubicado dentro de la zona de amortiguamiento de la Reserva Nacional de Tambopata (9°57´-13°20´ S, 68°39´-72°31´ W; 200 m), Madre de Dios, Perú. La fase preliminar para estandarizar los parámetros se adelantó entre los meses de Junio-Agosto del 2001 y los bioensayos se realizaron en los períodos de Abril-Mayo, Junio-Septiembre y Noviembre del 2002.
Esta región está clasificada como Bosque Tropical Lluvioso según Holdridge et al. (1971). Tiene una precipitación promedio de ~2100 mm año-1, con una estación seca desde Abril hasta Octubre y una estación lluviosa desde Noviembre hasta Marzo.
Sistema de estudio
Cordia nodosa (Boraginaceae): Es un arbusto o árbol de sotobosque conocido con el nombre vernacular de huevo de gato. Se encuentra ampliamente distribuido en la Amazonía Peruana, es una mirmecofila cuyos domacios están constituidos por ensanchamientos a nivel de las ramas (Fig. 1a). Estos domacios se originan con las yemas axilares, produciéndose seis hojas por cada domacio (Yu y Wilson 2001). Florece todo el año, aunque presenta picos entre Abril – Mayo y Agosto – Septiembre.

Allomerus demerarae (Formicidae: Myrmicinae): Localmente conocido con el nombre vernacular de «pucacuro» por el dolor que causa la picadura de su aguijón, aunque cabe mencionar que el término local «pucacuro» agrupa a diversas especies de hormigas pequeñas y rojizas con iguales características de picadura. A. demerarae es simbionte específico obligatorio de C. nodosa, su reina depende enteramente de C. nodosa para el establecimiento y supervivencia de la colonia (Yu y Wilson 2001). Tienen una sola reina por colonia y como toda colonia de hormigas, presenta una organización social en castas. La casta obrera no presenta subcastas morfológicamente diferenciables pero se sugiere que existe división de labores (Yu y Wilson 2001). Los bioensayos se realizaron con obreras que fueron agrupadas en dos categorías: las «patrulladoras» definidas por encontrarse patrullando las hojas jóvenes al momento de la colecta (Fig. 1b) y las «hormigas de domacio definidas así por ser coleccionadas del interior del domacio.
Interacción C. nodosa – A. demerarae:
C. nodosa es un arbusto que sufre altos índices de herbivorismo si no se asocia con alguno de sus seis potenciales simbiontes. Del mismo modo, estas hormigas son simbiontes obligatorios específicos puesto que no pueden desarrollarse sin la presencia de su hospedero (Yu y Wilson 2001). A diferencia de la interacción C. nodosa – Azteca, la interacción C. nodosa - A. demerarae no es mutualista sino mas bien parasítica (Yu y Pierce 1998). Una planta de C. nodosa que tiene A. demerarae como simbionte no llega a producir frutos, o lo hace con rarísimas excepciones. Las obreras de A. demerarae cortan el pedicelo de las diminutas flores de C. nodosa (Fig. 1c), evitando que continúen su desarrollo en la producción de frutos, de este modo redireccionan la energía de la planta hacia la producción de domacios obteniendo así mayor espacio para anidar (Yu y Wilson 2001).
Ubicación de plantas y colonias de hormigas. Para individualizar la plantas de donde se extrajeron los compuestos y las hormigas, se localizaron plantas de C. nodosa en 50 transectos paralelos, de 100 m de longitud, cada 25 m a través de una trocha principal ya existente. Se adjudicó un código a cada individuo de C. nodosa y se registró la siguiente información: lo-calización de la planta, altura (estimación relativa a la estatura personal), número de domacios (como referencia de la edad de la colonia), así como identidad del género de la hormiga que habitaba la planta. La identificación del género se hizo con base en diferencias morfológicas de A. demerarae versus Azteca sp.; hormigas del género Allomerus se diferencian de las del género Azteca en tamaño, coloración y en las forma de sus construcciones de cartón sobre la planta, además Azteca, despide un olor a limonel característico que facilitó su identificación en campo. Los extractos y las hormigas que se emplearon en los experimentos procedieron de individuos de C. nodosa provenientes de los censos. Para los bioensayos se obtuvieron los siguientes extractos de miembros del sistema C. nodosa - A. demerarae: hojas jóvenes, hojas maduras y huevos-larvas. Para obtener extractos de flores se emplearon las flores del sistema C. nodosa-Azte-ca. Los bioensayos fueron conducidos en un albergue que se implementó como Laboratorio.
Extracción de muestras húmedas de hojas, flores y huevos-larvas. Para la extracción de muestras húmedas de hojas jóvenes y de hojas maduras, se colectaron las hojas de un total de diez plantas. En el caso de los extractos de hojas maduras se colectó una hoja por planta y en el caso de los extractos de hojas jóvenes se colectaron todas las hojas jóvenes que tenía dicha planta, repitiéndose este procedimiento para las diez plantas. Este proceso disminuyó el efecto de las diferencias de área foliar entre hojas jóvenes y hojas maduras. Para la extracción de muestras húmedas de huevos - larvas de A. demerarae se colectaron tres domacios de diez plantas, luego se abrieron y se extrajeron los huevos y larvas de hormigas usando un par de pinzas. Para la extracción de muestras húmedas de flores de C. nodosa, se colectaron las inflorescencias de cinco plantas.
Las muestras se colocaron en un vaso de precipitado que contenía 25-50 ml de hexano. Se dejaron reposar por 30 min y luego se decantó el extracto húmedo. Los extractos fueron almacenados en un frasco que se mantuvo a bajas temperaturas (6°C).
Colecta de compuestos volátiles. Para colectar los compuestos volátiles naturalmente emitidos por las hojas jóvenes y hojas maduras, se utilizó la técnica denominada headspace. Esta técnica consistió en aislar el órgano emisor de los compuestos volátiles (hoja joven, hoja madura) con una bolsa para microondas (5 L). A esta bolsa se le insertaron dos tubos en los lados opuestos, uno conectado a un filtro de carbón activado que purificó el aire que se bombearía a través de él y el otro conectado a un filtro (ARS) que atrapó los compuestos volátiles arrastrados por la corriente de succión de aire. Cada filtro estuvo conectado a su vez, a un tubo flexible que se conectó a una bomba de aire. El tubo conectado al filtro de carbón se conectó a una bomba que expele aire, y el tubo conectado al filtro se conectó a una bomba que succiona aire. Cada bomba funciona con la energía proporcionada por una batería de 12 V. El flujo de aire dentro del ambiente aislado por la bolsa se mantuvo constante empleando un flujómetro. Para proteger la instalación de la lluvia se construyó un techo provisional de plástico. Este sistema se mantuvo activo por 24 h. Se colectó el filtro y se le envolvió en papel aluminio para protegerlo de la contaminación y se trasladó al laboratorio. Se usaron 400 uL de hexano para extraer los compuestos atrapados por el filtro, que se conservaron en viales Wheaton® ámbar 320 de 2 mL. Este extracto se mantuvo en refrigeración a 4°C hasta que fue requerido en los bioensayos. El sistema de headspace se instaló en plantas asociadas a A. demerarae para la colecta de compuestos volátiles de hojas jóvenes y hojas maduras. Se diferenció la hoja madura de la joven por el color más claro de esta última, por su mayor fragilidad y su menor tamaño.
Bioensayos. Se realizaron 30 repeticiones como mínimo por cada ensayo, con hormigas patrulladoras y con hormigas de domacio. Los bioensayos se llevaron a cabo en placas Petri, cuyas paredes se rociaron previamente con Fluon® (copolímero de tetrafloroetileno, Asahi Glass Company), para evitar que alguna de las 30 hormigas escape. Para los bioensayos que emplearon hormigas «patrulladoras» se colectaron las mismas directamente de las hojas con la ayuda de un par de guantes, que disminuyen la adherencia de las hormigas hacia los dedos, y se colocaron rápidamente en una placa Petri, tratando de minimizar el estrés provocado. Para los bioensayos que requerían hormigas de domacio, se colectaron domacios, que se abrieron con la ayuda de una tijera podadora, y auxiliados con un par de guantes las hormigas fueron rápidamente trasladadas del interior del domacio a la placa Petri. Los bioensayos se realizaron en un rango de temperatura no controlada de 26 a 31°C y entre las 11:00 a 16:00 h.
Los tratamientos fueron los siguientes: 1) Extracto de hojas maduras obtenido por la técnica de Headspace (M*); 2) Extracto de hojas jóvenes obtenido por la técnica de Headspace (N*); 3) Extracto de hojas maduras obtenido por extracción húmeda (M); 4) Extracto de hojas jóvenes obtenido por extracción húmeda (N); 5) Extracto de flores obtenido por extracción húmeda (F); 6) Extracto de huevos-larvas obtenidos por extracción húmeda (H). Tanto para los extractos obtenidos por extracción con headspace como para los extractos obtenidos por extracción húmeda, el control fue hexano puro, g.a. ©.
El procedimiento general para llevar a cabo los bioensayos fue el siguiente: se colocaron 30 obreras patrulladoras» o 30 obreras de domacio» dentro de una placa Petri. Se tomó un disco de papel filtro (Whatman© N°1, 3 cm de diámetro) y se aplicaron 100 uL del extracto correspondiente. Inmediatamente se colocó el disco de papel filtro en la placa Petri. Se repitió el procedimiento para aplicar el segundo extracto y para aplicar el hexano empleado aquí como control. Para evitar sesgo en los resultados y para minimizar el efecto de los diferentes tiempos de aplicación de los extractos, se procedió a rotar la distribución espacial de los tres discos en sentido horario, en ángulo de 120º. Además el orden de aplicación de los extractos se determinó al azar con el lance de un dado. El tiempo se registró desde el momento en que se colocó el tercer disco de papel filtro, con el extracto ya aplicado. En el primer minuto se registraron todas las hormigas que estuvieron sobre, debajo o tocando con sus antenas el papel filtro para cada tratamiento y para el control. Se repitió lo mismo para el minuto dos y así sucesivamente hasta el minuto 10. En cada bioensayo se enfrentaron dos tratamientos y un control.
Hipótesis analizadas. Se probaron las siguientes hipótesis:
Hipótesis A: Las obreras patrulladoras son atraídas a extractos de hojas jóvenes con mayor intensidad que a los extractos de hojas maduras.
Hipótesis A: Las obreras de domacio son atraídas a extractos de hojas jóvenes con mayor intensidad que a los extractos de hojas maduras.
Para ambos casos, los tratamientos fueron: extracto de hojas jóvenes, extracto de hojas maduras y el control (hexano puro). En los bioensayos con obreras patrulladoras y de domacio se realizaron 37 y 66 repeticiones, respectivamente.
Hipótesis B: Las obreras patrulladoras son atraídas a extractos de hojas jóvenes con mayor intensidad que a los extractos de huevos-larvas.
Hipótesis B: Las obreras de domacio son atraídas a extractos de hojas jóvenes con mayor intensidad que a los extractos de huevos-larvas.
Para ambos casos los tratamientos fueron: extracto de hojas jóvenes, extracto de huevos-larvas de A. demerarae y el control (hexano puro). En los bioensayos con obreras patrulladoras y de domacio se realizaron 30 y 60 repeticiones respectivamente.
Hipótesis C: Las obreras patrulladoras son atraídas a extractos de flores con mayor intensidad que a los extractos de hojas jóvenes.
Hipótesis C: Las obreras de domacio son atraídas a los extractos de flores con mayor intensidad que a los extractos de hojas jóvenes.
Para ambos casos, los tratamientos fueron: extracto de hojas jóvenes, extracto de flores de C. nodosa y el control. En los bioensayos con obreras patrulladoras y de domacio se realizaron 32 y 31 repeticiones, respectivamente.
Análisis estadístico. Para evitar el efecto de las diferencias de actividad a nivel de colonia, el número de obreras atraídas a un determinado tratamiento en cada bioensayo fue expresado como número total de hormigas que respondieron en dicho bioensayo. En todos los casos, se excluyeron las hormigas que no fueron atraídas a ninguno de los tres tratamientos. Los datos fueron transformados a raíz cuadrada de x + 0,5 con el fin de asegurar la homogeneidad de las varianzas mediante la prueba de Levene y la normalidad de los datos mediante la prueba de Kolmogorov-Smirnov (Zar 1996). Los registros de cada bioensayo fueron analizados por minuto, del uno al 10, pero debido a la naturaleza de reclutamiento de las obreras, para el análisis global se excluyeron los cuatro primeros minutos requeridos para que el comportamiento se estabilice. Se promediaron los datos expresados en porcentaje con respecto al total de obreras que respondieron a los tres tratamientos del minuto cinco al 10 y se empleo el ANDEVA (diferencias significativas con α a < 0.05), para determinar las diferencias entre tratamientos. Se usó el test de Tukey para determinar cuáles tratamientos difirieron entre sí. Adicio-nalmente, se usó el análisis de correlación de Pearson (r) para determinar la asociación entre los tiempos de exposición (min) y los diferentes tratamientos empleados en los bioensayos. Todos los cálculos estadísticos se realizaron empleando el programa estadístico SPSS® 12,0 (Norusis 2004).
Resultados
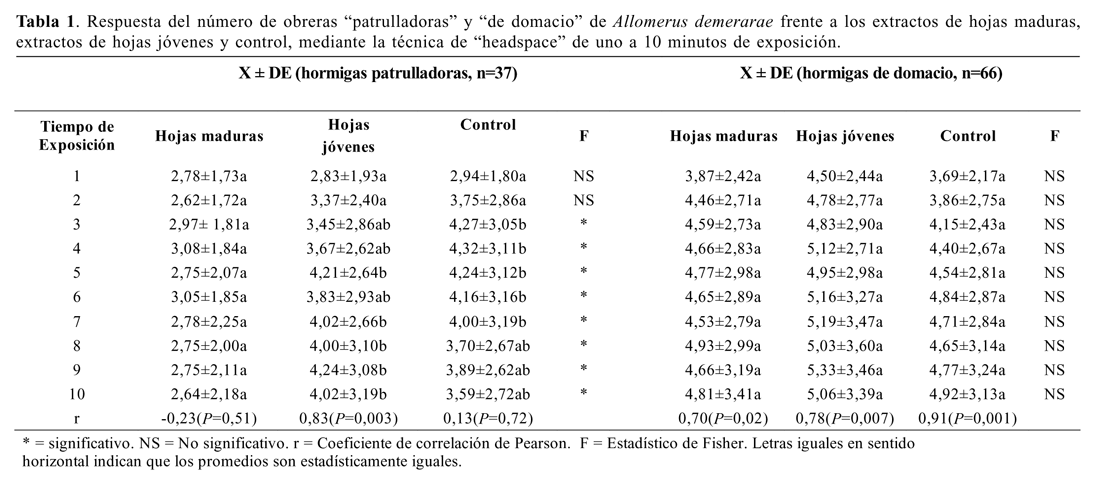
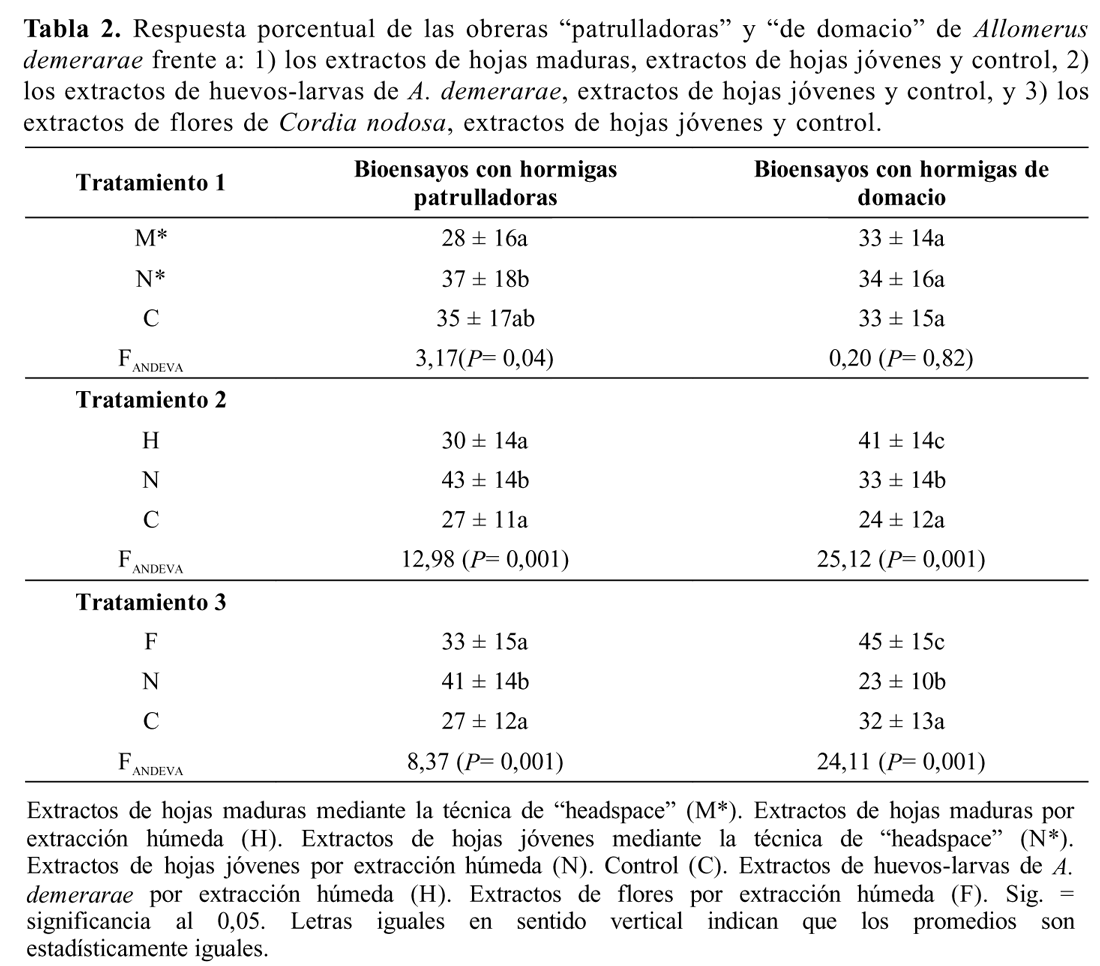
Extractos de hojas maduras (M*), extractos de hojas jóvenes (N*) y control (C). Se llevaron a cabo 37 bioensayos con obreras patrulladoras. El ANDEVA mostró diferencias significativas entre tratamientos del minuto tres al 10 (Tabla 1). La atracción hacia el tratamiento hojas jóvenes (N*) fue significativamente mayor que hacia hojas maduras (M*). No hubo diferencias significativas entre hojas maduras y control, ni entre hojas jóvenes y el control (Tabla 1 y 2). Solo se encontró una correlación significativa entre el tiempo de exposición (min) y la respuesta de las obreras patrulladoras al extracto de hojas jóvenes (Tabla 1).
Se llevaron a cabo 66 bioensayos con obreras de domacio. El ANDEVA no mostró diferencias significativas entre tratamientos (Tabla 1 y 2). Se encontraron correlaciones significativas entre el tiempo de exposición (min) y la respuesta de las obreras de domacio a los extractos de hojas maduras y jóvenes, así como entre tiempo de exposición y el control (Tabla 1).
Extractos de huevos-larvas (H), extractos de hojas jóvenes (N) y control (C).
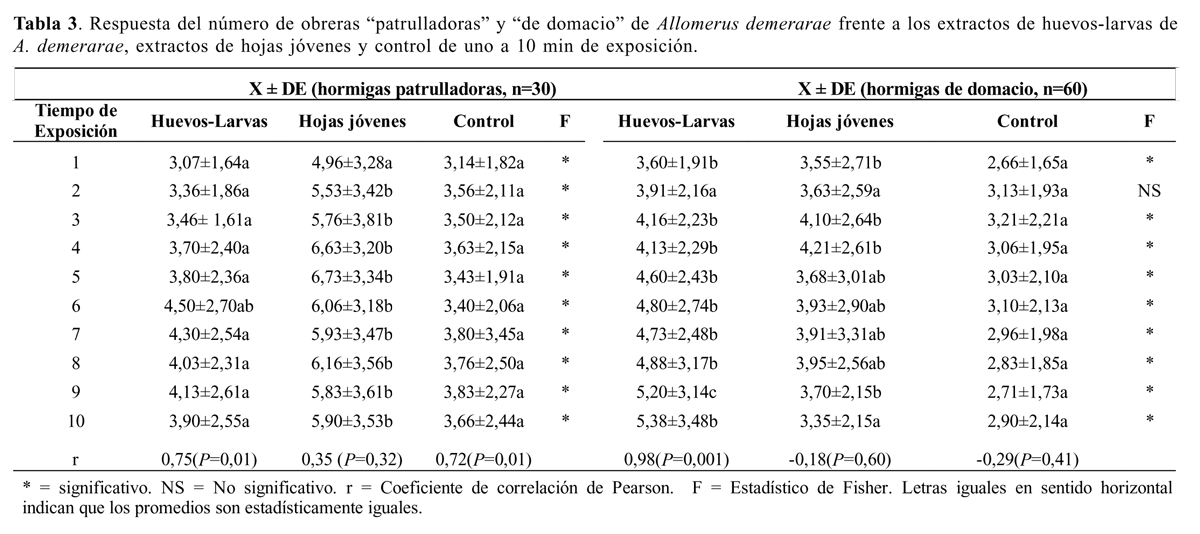
Se llevaron a cabo 30 bioensayos con obreras patrulladoras (Tabla 3). El ANDEVA mostró diferencias significativas entre tratamientos del minuto uno al diez (Tabla 3). La atracción hacia el tratamiento de hojas jóvenes (N) es significativamente mayor que huevos-larvas (H) y el control (C), y no hubo diferencias significativas entre H y C (Tabla 2). Se encontraron correlaciones significativas entre el tiempo de exposición y la respuesta de las obreras patrulladoras al extracto huevos-larvas; así como entre tiempo de exposición y el control (Tabla 3).
Se llevaron a cabo 60 bioensayos con obreras de domacio (Tabla 3). Se observaron diferencias altamente significativas entre los tratamientos, excepto en el minuto dos. La atracción hacia el tratamiento huevos-larvas (H) es significativamente mayor que la atracción hacia hojas jóvenes (N) y el control (C); y hojas jóvenes (N) es significativamente mayor que el control (C) (Tabla 2 y 3). Solo se encontró una correlación significativa entre el tiempo de exposición y la respuesta de las obreras de domacio al extracto huevos-larvas (Tabla 3.
Resultados extractos de flores (F), extractos de hojas jóvenes (N) y control (C).
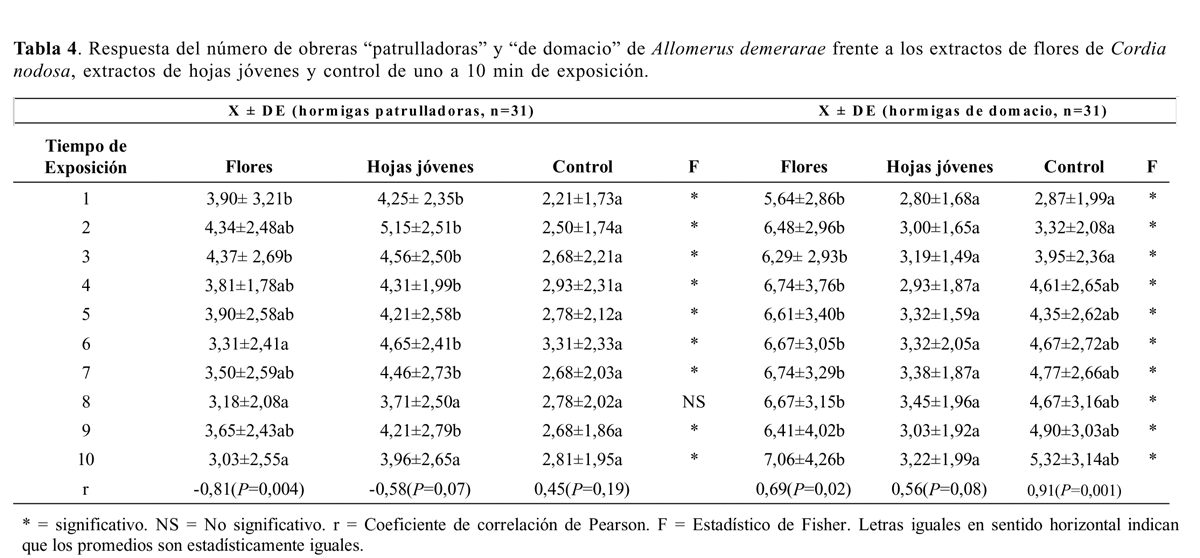
Se llevaron a cabo 31 bioensayos con obreras patrulladoras. Se encontraron diferencias altamente significativas entre tratamientos (Tabla 4). La atracción hacia el tratamiento de hojas jóvenes (N) fue significativamente mayor que hacia flores (F) y que control (C), mientras que no hubo diferencias entre flores (F) y control (C) (Tabla 2). Solo se encontró una correlación significativa negativa entre el tiempo de exposición y la respuesta de las obreras patrulladoras al extracto flores (Tabla 4). Se llevaron a cabo 31 bio-ensayos con obreras de domacio. Se encontraron diferencias altamente significativas entre tratamientos (Tabla 4). La atracción hacia el tratamiento flores (F) fue significativamente mayor que hacia hojas jóvenes (N) y que hacia el control (C). N fue significativamente menor que C (Tabla 2 y 4). Solo se encontró una correlación significativa entre el tiempo de exposición y la respuesta de las obreras de domacio al extracto flores y al control (Tabla 4).
Discusión
La intensidad de la respuesta de las obreras, en términos de número de obreras para cada tratamiento, fue en general bastante baja, en comparación con la actividad mostrada en su medio natural en la que normalmente actúan con decenas de individuos. Esto puede deberse a las bajas concentraciones de los químicos estimulantes presentes en los extractos aplicados a los discos de papel filtro. Por otro lado, para que una obrera realice determinada tarea (patrullaje, forrajeo, defensa, cuidado de la prole, etc.), su rol dependerá de la interacción con las otras obreras (Gordon 1999), es decir que, los resultados pueden explicarse sugiriendo que el comportamiento de reclutamiento de las obreras requiera de más señales que proporcionen el contexto adecuado para desencadenar la respuesta esperada. Sin embargo, las condiciones ofrecidas en este medio artificial han sido suficientes para evidenciar diferencias etológicas.
Los resultados de los bioensayos con obreras patrulladoras de A. demerarae frente a extractos de hojas nuevas y extractos de hojas maduras, verifica que de todos los compuestos secundarios emitidos por estas hojas, la porción volátil de los extractos de hojas jóvenes o nuevas es suficiente, por sí sola, para desencadenar una respuesta diferencial en el comportamiento de las obreras (Tablas 1 y 2).
Los resultados de los bioensayos llevados a cabo con obreras de domacio, por otro lado, no mostraron una mayor preferencia por alguno de los tres tratamientos (extractos de hojas jóvenes, extractos de hojas maduras y control) (Tablas 1 y 2). Sin embargo, esto no debe considerarse como evidencia en contra de la hipótesis que sugiere la potencia de los compuestos volátiles como señal suficiente para desencadenar el comportamiento de patrullaje, sino como evidencia de la diferente sensibilidad que poseen las obreras de domacio por ser un grupo especializado en otras labores.
Dadas las diferencias en el patrón de respuestas de hormigas procedentes del interior de los domacios y las de hormigas que están patrullando hojas jóvenes, frente al mismo conjunto de estímulos químicos, se evidencia la existencia de un sistema de asignación de roles regulado por mediación química al interior de la casta obrera de A. demerarae. Las obreras patrulladoras muestran una preferencia por los extractos de hojas jóvenes frente a extractos de hojas maduras, extractos de huevos-larvas y extractos de flores. Mientras que las obreras colectadas del interior del domacio mostraron preferencia por los extractos de huevos y de flores frente a los extractos de hojas nuevas, no mostrando preferencia por los extractos de hojas nuevas cuando están frente a los extractos de hojas maduras (Tabla 2).
Las hormigas que se colectaron de las hojas nuevas estarían encargadas de velar por el buen estado de salud de las hojas nuevas de la planta y en este caso el término patrulladora estaría correctamente asignado; las hormigas procedentes del interior del domacio tendrían la función de tareas al interior del nido, es decir de cuidar los huevos, larvas y pupas, y por esto podríamos denominarlas nodrizas.
La carencia de diferencias morfológicas sugiere que la división de labores en las colonias de A. demerarae esté basada en la asignación de roles según la edad de las obreras, es decir que estaríamos frente a un caso de polietismo temporal, en el que las obreras mas jóvenes serían las nodrizas y las adultas serían las patrulladoras. Sin embargo, se requiere de un análisis morfométrico comparativo más exhaustivo de estos dos grupos de hormigas para determinar si existen o no diferencias morfológicas. Para determinar la existencia de polietismo temporal hace falta determinar las edades de las obreras o por lo menos su estado fisiológico y comparar las actividades que realizan los estadíos juveniles versus los estadíos adultos. Pie (2002) utilizó la coloración corporal como indicador de la madurez de las hormigas, en las que las de color más pálido serían juveniles y las de color más oscuro serian las más viejas.
Frederickson (2001) sugirió que es posible que las obreras de A. demerarae estén colectando sustancias presentes en la flores de C. nodosa para luego aplicarlas a sus huevos, basándose en las observación de que todos los compuestos químicos encontrados en los extractos de huevos de A. demerarae también estaban presentes en los extractos de las flores de A. demerarae. Los resultados de esta investigación refuerzan esta teoría al reportar que las obreras encargadas de castrar a C. nodosa, es decir, de cortar las flores, son las mismas que las encargadas del cuidado de la prole.
La mayoría de investigaciones que evalúan la existencia de grupos especializados de obreras parten de la observación de diferencias morfológicas en las obreras y se basan en observaciones etológi-cas llevadas a cabo en nidos artificiales (Traniello 1978; Traniello y Jayasurika 1985; Pie 2002). En estas investigaciones se usan químicos para determinar la diferencia entre grupos especializados. Sabiendo que el medio principal de comunicación entre hormigas es el olfato, sin descartar el tacto, se utiliza señales químicas para identificar los grupos especializados (Dicke 2000). Por lo pronto, aislando los compuestos químicos de los huevos se han identificado a las obreras especializadas en el cuidado de los estadios juveniles, aislando los compuestos químicos de las hojas jóvenes se han identificado a las obreras especializadas en el patrullaje de las hojas jóvenes y finalmente aislando los compuestos químicos de las flores se ha determinado el grupo de obreras especializado en el comportamiento de castración. Este protocolo es de gran utilidad sobre todo porque aún no se han desarrollado técnicas eficientes para reproducir un nido de hormigas que viven en asociación con mirmecofilas.
Conclusiones
El conjunto de compuestos volátiles emitidos por las hojas jóvenes de C. nodosa constituyen una señal suficiente para dirigir el comportamiento de patrullaje en las colonias de A. demerarae. En cambio, las hormigas obreras de domacio de A. demerarae, la atracción fue significativamente mayor hacia los huevos-larvas de A. demerarae y las flores de C. nodosa. Los resultados muestran evidencia de la existencia de polietismo en las colonias de A. demerarae, en la que se han identificado dos grupos de obreras con funciones diferentes. Se sugieren dos subcastas, las del interior del domacio serian las nodrizas y las colectadas de las hojas jóvenes serian las patrulladoras. Las nodrizas serían además las encargadas del comportamiento de castración de las flores de C. nodosa.
Literatura citada
BRONSTEIN, J. L. 1998. The contribution of ant – plant protection studies to our understanding of mutualism. Biotropica 30: 150-161. [ Links ]
BROUAT, C.; McKEY D.; BESSIERE, J.M.; PASCAL, L.; HOSSAERT – McKEY, M. 2000. Leaf volatile compounds and the distribution of ant patrolling in an ant – plant protection mutualism: Preliminary results on Leonardoxa (Fabaceae: Caesal-pinoideae) and Petalomyrmex (Formicidae: Formicinae). Acta Oecologica 21: 349-357. [ Links ]
CHAPIN, F.S.; ZAVALETA, E.S.; EVINER, V.T.; NAYLOR, R.L.; VITOUSEK, P.M.; REYNOLDS, H.L.; HOPPER, D.U.; LAVOREL, S.; E.SALA, O.; HOBBIE, S.E.; MACK, M.C.; DIAZ, S. 2000. Consequences of changing biodiversity. Nature 405: 234-242. [ Links ]
CHAPMAN, R.F. 2003. Contact chemo-reception in feeding by phytophagous insects. Annual Review of Entomology 48: 455-484. [ Links ]
DAVIDSON, D. W.; SEIDEL, J. L.; EPS-TEIN, W. W. 1990. Neotropical ant gardens I. Chemical constituents. Journal of Chemical Ecology 16: 1791-1815. [ Links ]
DAVIDSON, D. W.; McKEY, D. 1993. The evolutionary ecology of symbiotic ant – plant relationships. Journal of Hymenop-tera Research 2: 13-83. [ Links ]
DEJEAN, A.; QUILICHINI, A.; DELABIE, J.C.H.; ORIVEL, J.; CORBARA, B. ; GIBERNAU, M. 2004. Influence of its associated ant species on the life history od the myrmecophyte Cordia nodosa in French Guiana. Journal of Tropical Ecology 20: 701-704. [ Links ]
DICKE, M. 2000. Chemical ecology of host-plant selection by herbivorous arthropods: a multitrophic perspective. Biochemical and Systematic Ecology 28:601-617. [ Links ]
DUFAY, M.; HOSSAERT-McKEY, M.; ANSTETT, M.C. 2003. When leaves act like flowers: how dwarf palms attract their pollinators. Ecology letters 6: 28-34. [ Links ]
Du TOIT, J.T.; WALKER, B.H. ; CAMPBELL, B.M. 2004. Conserving Tropical nature: current challenges for ecologists. Trends in Ecology and Evolution 19: 12-17. [ Links ]
F´EN´ERON, R.; DURAND, J. L.; JAISSON, P. 1996. Relation between behaviour and physiological maturation in a ponerine ant. Behaviour 133: 791-806. [ Links ]
FREDERICKSON, M. 2001. Chemical ecology of a Neotropical ant – plant system: Interactions between Cordia nodosa, mutualistic symbionts Azteca spp., and the cheater species, Allomerus demerarae. Thesis. Harvard University. Boston. 66 p. [ Links ]
FREDERICKSON, M. 2005. Ant species confer different partner benefits on two neotropical mymecophytes. Oecologia 143: 387-395. [ Links ]
GAUME, L.; ZACHARÍAS, M.; GROS-BOIS, V.; BORGES, R.M. 2005. The fitness consequences of bearing domatia and having the right ant partner: experi-ments with protective and non-protective ants in a semi-myrmecophyte. Oecologia 145: 76-86. [ Links ]
GAUTRAIS, J.; THERAULAZ, G., DENEU-BOURG, J.L.; ANDERSON, C. 2002 Emergent polyethism as a consequence of increased colony size in insect societies. Journal of Theoretical Biology 214: 1-11. [ Links ]
GOMULKIEWICZ, R.; NUISMER, S.L.; THOMPSON, J.N. 2003. Coevolution in variable mutualisms. The American Naturalist 162: S80-S93. [ Links ]
GORDON, D.M. 1999. Ants at work: how an insect society is organized. Simon and Schuster Ed. New York, USA. 182 p. [ Links ]
HEIL, M.; McKEY, D. 2003. Protective ant-plant interactions model systems in ecological and evolutionary research. Annual Review of Ecology, Evolution and Systematics 34: 425-553. [ Links ]
HOECHBERG, M.E.; GOMULKIEWICZ, R.; HOLT, R.D.; THOMPSON, J.N. 2000. Weak sinks could cradle mutualistic symbioses-strong sources should harbour parasitic symbioses. Journal of Evolutio-nary Biology 13: 213-222. [ Links ]
HOLDRIDGE, L.R.; GRENKE, W.C.; HATHEWAY, W.H.; LIANG, T.; TOSI, J.A. 1971. Forest Environments in Tropical Life Zones: A Pilot Study. Pergamon Press. Oxford, England. 735 p. [ Links ]
HOLLDÖBLER, B.; WILSON, E.O. 1990. The Ants. The Belknap Press of Harvard University Press. Cambridge. Massa-chusetts. 732 p. [ Links ]
HOYT, E. 1998. Superorganism. p. 66-100. En: Hoyt E. (Ed.). Earth dwellers: Adven-tures in the Land of Ants. Mainstream Publishing Company. Edinburgh, UK. 301 p. [ Links ]
HUGHES, W.O.H.; GOULSON, D. 2001. Polyethism and the importance of context in the alarm reaction of the grass-cutting ant, Atta capiguara. Behavioral Ecological and Sociobiology 49: 503-508. [ Links ]
JANZEN, D. H. 1966. Coevolution of mutualism between ants and acacias in Central America. Evolution 20: 249-275. [ Links ]
KRICHER, J. 1997. A Neotropical companion: An introduction to the animals, plants and ecosystems of the New World Tropics. Princeton University Press. Princeton. NJ. USA. 451 p. [ Links ]
LOREAU, M.; ÑAME, S.; INCHAUSTI, P.; BENGTSSON, J.; GRIME, J.P. ; HEC-TOR, A.; HOOPER, D.U.; HUSTON, M.A.; RAFFAELLI, D.; SCHMID, B.; TILMAN, DF.; WARDLE, D.A. 2001. Biodiversity and ecosystem functioning: current knowledge and future challenges. Science 294: 804-808. [ Links ]
McKEY, D. 1984. Interaction of the ant- plant Leonardoxa africana (Caesalpinoideae) with its obligate inhabitants in a rainforest in Cameroon. Biotropica 16: 81-99. [ Links ]
NORUSIS, M. 2004. SPSS 12.0 Guide to data analysis. Prentice Hall. New Jersey. USA. 647 p. [ Links ]
PIE, M. R. 2002. Behavioral repertoire, age polyethism and adult transport in Ecta-tomma opaciventre (Formicidae: Poneri-nae) Journal of Insect Behavior. 15: 25-35. [ Links ]
SENDOVA, A.B.; FRANKS, N.R. 1999. Self-assembly, self-organization and division of labour. Philosophical Transactions of the Royal Society of London 354: 1395-1405. [ Links ]
SOLANO, P.J.; DEJEAN, A. 2004. Ant-fed plants comparison between three geophitic myrmecophytes. Biological Journal of the Linnean Society 83: 433-439. [ Links ]
SOLANO, P.J.; BELIN-DEPOUX, M.; DEJEAN, A. 2005. Formation and structure of food bodies in Cordia nodosa (Boraginaceae). Comptes rendus biologies 328: 642-647. [ Links ]
THOMPSON, J. 1982. Interaction and coevolution. John Wiley & Sons Editors. NY. USA. 179 p. [ Links ]
THOMPSON, J. 1999. Coevolution an escalation: are ongoing coevolutionary meandering important?. The American Naturalist 153: S92-S93. [ Links ]
THOMPSON, J.; CUNNINGHAM, B.M. 2002. Geographic structure and dynamics of coevolutionary selection. Nature 417: 735-738. [ Links ]
TRANIELLO, J. F. A. 1978. Caste in a primitive ant: absence of age polyethism in Amblyopone. Science 202:770-772. [ Links ]
TRANIELLO, J. F. A.; JAYASURIYA, A. K. 1985. The biology of the primitive ant Aneuretus simoni (Emery) (Formicidae: Aneuretinae). II. The social ethogram and division of labor. Insectes Sociaux. 32: 375-388. [ Links ]
WILSON, E.O. 1980. Caste and division of labor in leaf-cutter ants (Hymenoptera: Formicidae: Atta). Behavioral Ecology and Sociobiology 7:157-165. [ Links ]
YU, D. W. 1997. Parasitic castration of the ant – plant Cordia nodosa and the consequen-ces for a metapopulation model of species coexistence. Tesis Ph. D. University of Harvard. Boston. USA. [ Links ]
YU, D. W.; PIERCE, N. E. 1998. A castration parasite of an ant – plant mutualism. Proceedings of the Royal Society of London 265: 375-382. [ Links ]
YU, D. W.; WILSON, H. B. 2001. The competition-colonization trade-off is dead: Long live the competition-colonization trade-off. The American Naturalist 158: 49-63. [ Links ]
ZAR, J. H. 1996. Bioestatistical analysis. 3th Ed. Prentice-Hall. Inc. Upper Saddle River, New Jersey. USA. 662 p. [ Links ]
Recibido: 20-ene-05 Aceptado: 10-ene-06


{kind=link}
{kind=link}
{kind=link}
{kind=link}