










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488On-line version ISSN 2665-4385
Rev. Colomb. Entomol. vol.32 no.2 Bogotá July/Dec. 2006
Sección Medica
Interacción de Leishmania (L) chagasi con la línea celular Lulo en diferentes condiciones ambientales
Interaction of Leishmania (L) chagasi with the Lulo cell line in different environmental conditions
Víctor Acero1, Edna Galeano1, Martha Ayala2, Jaime Castellanos3, Felio Bello4
1Laboratorio de Investigaciones en Entomología, Biología Celular y Genética, Departamento de Ciencias Básicas, Universidad de La Salle, Bogotá, D. C., Colombia. victoraceromv@yahoo.es
2Laboratorio de Parasitología, Instituto Nacional de Salud de Colombia, Bogotá, D.C., Colombia. mayalas@ins.gov.co
3Instituto de Virología, Universidad El Bosque, Bogotá, D.C., Colombia. castellanosjaime@unbosque.edu.co
4Ciencias Básicas, Facultad de Medicina, Universidad del Rosario, Bogotá D.C., Colombia. Calle 63D No. 24-31, Teléfono: (571) 3474570, extensión 257; fax: (571) 3101275. E-mail: fbello@urosario.edu.co
Resumen.Existen eventos desconocidos o pobremente comprendidos en las interacciones Leishmania-célula hospedera que ameritan estudios en nuevos modelos in vitro. El propósito del presente trabajo fue evaluar, teniendo en cuenta variables de temperatura, concentración de CO2 y cambios morfométricos de las células, la interacción de Leishmania chagasi con una línea celular (Lulo), previamente establecida, derivada de tejidos embrionarios de Lutzomyia longipalpisi . La línea celular J774 se utilizó como control positivo de la infección. La cepa MH/CO/84/CI-044B de L. chagasi se usó para los ensayos de infección. Los parásitos se adicionaron a las células adheridas, tanto de Lulo como de J774, en una proporción de 5:1, la incubación se efectuó a 28°C y 37°C, en ausencia y presencia de CO2. Se realizó un estudio morfométrico de las células antes y después de la infección. Las características ultraestructurales, tanto del parásito como de las células, se analizaron por microscopía electrónica. El mayor porcentaje de infección en las células Lulo a 28°C se registró el día seis post-infección (26,8%), mientras que a la temperatura de 37°C, sin atmósfera de CO2, el máximo valor (30,4%) se obtuvo el día nueve. Los análisis morfométricos mostraron un aumento significativo en el tamaño de las células infectadas de Lulo a 28°C y de J774 a 37°C con y sin atmósfera de CO2respectivamente.
Palabras clave: Características ultraestructurales. Temperatura y co2. Promastigote. Porcentaje de infección. Morfometría celular.
Summary.There are unknown or poorly understood events in the Leishmania-host cell interactions that merit studies in new in vitro models. The aim of the present work was to evaluate Leishmania chagasi interaction with a cell line (Lulo) previously established and derived from Lutzomyia longipalpisi embryonic tissues, bearing different temperatures and CO2 concentrations in mind and morphometric changes of the cells. The J774 cell line was used as positive control of the infection. The L. chagasi MH/CO/84/CL-044B strain was used for infection assays. The parasites were added to adherent cells in 5:1 ratio, incubated at 28°C and 37°C with the presence or absence of CO2. A morphometric study of cells was carried out, before and after infection. Both parasite and cell ultrastructure characteristics were analyzed by electron microscope. The greatest percentage of infection in the Lulo cells at 28°C was registered at 28°C (26,8%) on day six, while at 37°C, in CO2 absence, the maximum percentage (30,4%) was obtained the ninth day. Morphometric analysis showed a significant increase in the size of Lulo cells infected at 28°C and J774 cells at 37°C with the presence and absence of CO2 respectively.
Key Words. Ultrastructure characteristics. Temperature and cO2. Promastigote. Percentage of infection. Cell morphometry.
Introducción
Los protozoos flagelados del género Leishmania son parásitos intracelulares obligados que asumen durante su ciclo de vida varios estados morfológicos y funcionales (Abdulrahman et al 1998). Sin embargo, experimentalmente se ha encontrado en el canal alimenticio del flebótomo, después de una segunda comida de sangre y en días avanzados de infección, formas del parásito semejantes a amastigotes (Añez et al 2003) esto plantea nuevos interrogantes sobre el ciclo biológico del parásito, particularmente en el vector. El desarrollo del ciclo biológico de los parásitos del género Leishmania puede ser experimentalmente estudiado in vivo e in vitro. En el primer caso se utilizan hembras de flebótomos para inducir la transformación de especies de Leishmania, particularmente de la forma amastigota a la promastigota, mediante la ingesta de sangre que contiene amastigotes, tomada a través de membranas artificiales. La diferenciación de los promastigotes ocurre en el intestino del insecto, donde además éstos se replican y pueden observarse en diferentes estados morfológicos; luego la transmisión se realiza en condiciones de laboratorio por la picadura a ratones ó hámsters (Bates 1997; Killick-Kendrick 1986; Molyneux y Killick-Kendrick 1986). Experimentos in vitro emplean células de exudado peritoneal de ratones y hámsters, igualmente células macrofágicas tumorales de ratón para reproducir los mecanismos de diferenciación, replicación y maduración de los parásitos (Aikawa et al 1982; Pearson et al 1981; Chang y Dwyer 1976). De forma similar, estos eventos se han podido estudiar en líneas celulares de origen no macrofágico, principalmente en cultivos de fibroblastos (Hérvas-Rodriguez et al 1996; Corte-Real 1995; Schwartzman y Pearson 1985; Dedet et al 1983; Chang 1978, 1979; Mattock y Petters 1975). Por otra parte, los medios axénicos que frecuentemente se emplean para mantener estos parásitos en condiciones de laboratorio se han usado para lograr la transformación in vitro de Leishmania, principalmente de la forma promastigote a la amastigote, mediante temperaturas elevadas y pH ácidos (Aquino et al 2002; Saar et al 1998; León et al 1995; Hodgkinson et al 1996; Bates et al 1992, 1993, 1994; Zilberstein 1991; Pan 1984).
Los cultivos celulares de insectos vectores se emplean en forma extensiva para estudios in vitro de arbovirus y también, en investigaciones biomédicas y tecnológicas (Crampton et al 1997; Igarashi 1985, 1978; Kuno et al 1985; Singh 1967; Kitamura 1965). En esta última línea de acción, han empezado también a explorarse los usos en estudios de co-cultivos con parásitos (Fampa et al 2003; Leake 1997), cobrando esta situación particular relevancia si se tiene en cuenta que, a pesar de los adelantos obtenidos con la implementación de tecnologías avanzadas para el estudio in vivo e in vitro de los parásitos del género Leishmania, aún existen procesos bioquímicos, inmunológicos y moleculares desconocidos o pobremente conocidos en las interrelaciones parásito-hospedero y parásito-vector que ameritan la utilización de nuevos modelos celulares, que permitan enfocar la atención a los eventos que ocurren entre los parásitos y las células derivadas del vector.
En un trabajo preliminar Bello et al (2005) describieron el proceso de infección experimental de L. chagasi en la línea celular Lulo, sustratos celulares que provienen de tejidos embrionarios de L. longipalpis, el vector primario de este parásito en su ambiente natural; Estos autores compararon este proceso con otro desarrollado paralelamente en las células J774, tomando como referencia de la evaluación de ambos procesos los días post-infección dos, cuatro, seis, ocho y 10. Sin embargo, no se valuaron factores ambientales ni cambios morfométricos de las células. En el presente trabajo se evaluó la interacción de L. chagasi con la línea celular Lulo (Rey et al 2000) teniendo en cuenta variables de temperatura y concentración de co2, para determinar, mediante microscopía de luz, los porcentajes e índices de infección y las modificaciones morfológicas celulares inducidas por la interacción con el parásito. Se analizatron además las características ultraestructurales del parásito y de las células con el apoyo de microscopía electrónica.
Materiales y Métodos
El trabajo se realizó en el Laboratorio de Entomología, Biología Celular y Genética, de la Universidad De La Salle, tuvo una duración de un año y medio (2003-2005).
Parásitos. Se utilizó la cepa MH/CO/84/CL-044 de L. chagasi , suministrada por el laboratorio de Parasitología del Instituto Nacional de Salud de Colombia, la cual se mantuvo en el medio NNN bifásico modificado con Schneider’s y enriquecido con suero fetal bovino (SFB) (Gibco®) inactivado al 10% e incubado a 26°C.
Cultivos celulares. Se empleó la línea celular Lulo (Rey et al 2000) derivada de explantes de tejidosembrionarios del flebótomo L. longipalpis (Lutz y Neiva 1913), principal vector de L. chagasi (Cunha y Chagas 1937). Esta línea celular se mantuvo in vitro en una mezcla de medio L-15 (Sigma®) y Grace (Sigma®), suplementado con 5% de suero fetal bovino (SFB), penicilina 100 unidades/ml y estreptomicina 100 µg/ml (Gibco®), a 28°C. Los subcultivos se efectuaron por remoción mecánica, previa observación de la monocapa confluente. Como control positivo de la infección se utilizó la línea celular J774, aislada de células reticulares de un sarcoma de ratón (Mus musculus) BALB/c, establecida por Ralph et al (1975). Para el mantenimiento de estas células se utilizó el medio RPMI (Sigma®), suplementado con SFB inactivado al 5% y la incubación se realizó a 37°C.
Para los ensayos de infección, conteo y morfometría, una vez se alcanzó la confluencia en la monocapa de ambas líneas celulares, se removieron mecánicamente las células y a continuación se centrifugaron a 900 g por 10 min, el botón de células se resuspendió en el medio L-15/Grace, se hizo conteo y se sembraron aproximadamente 2 x 105 células por cada pozo sobre laminillas de 12 mm de diámetro, en cajas de 24 pozos y se dejaron en crecimiento por 24 horas hasta lograr adherencia completa de las células.
Infección. A cada pozo de la caja de cultivo se adicionó 1 ml de una suspensión de parásitos en forma de promastigote (1 x 106 parásitos/ml), éstos se tomaron en la fase estacionaria y la proporción aproximada parásito/célula fue de 5:1 (Pessotti et al 2004; Fampa et al 2003). Luego, se llevaron a incubación a 28 °C y a 37 °C por 24 horas. Réplicas exactas sin parásitos fueron incubadas al mismo tiempo como controles. Las cajas de cultivo incubadas sin co2 a ambas temperaturas fueron selladas con Parafilm. Las cajas incubadas en ambiente de co2 al 5% no se sellaron, para permitir el intercambio gaseoso. Después de 24 horas de incubación, todas las células fueron lavadas con solución salina para retirar el exceso de parásitos y se adicionó medio fresco L-15/Grace. Los cultivos se observaron diariamente con un microscopio invertido (Olympus CK2). Al tercer día post-infección, se retiró el medio de cultivo de todos los pozos y se sacaron las laminillas correspondientes a ese día, al resto de pozos se les adicionó medio fresco. El mismo procedimiento se realizó en los días seis y nueve post-infección. Las laminillas se dejaron secar al ambiente por una hora y posteriormente se fijaron con metanol absoluto (Merck®) por 24 horas. Luego fueron teñidas con el colorante de Giemsa (Merck®) al 10% durante 10 minutos, se lavaron con agua destilada para retirar los excesos de colorante y se dejaron secar durante 24 horas. Finalmente, se les realizó un montaje permanente sobre láminas portaobjetos con Entellán (Merck®).
Conteos y análisis de infección. Las laminillas con ambos tipos de células se observaron en unfotomicroscopio (Olympus BX60) con objetivo de inmersión (100X) para realizar el conteo de las células infectadas y el número de amastigotes en cada una de ellas. Se contaron en promedio 200 células por cada laminilla y fueron evaluadas cuatro de tres experimentos independientes. Se determinó el porcentaje e índice de infección; este último parámetro se calculó multiplicando el porcentaje de infección celular por el promedio de amastigotes/célula (Lonardoni et al 2000). Las variables que se tuvieron en cuenta en el proceso de infección fueron: a) temperaturas de 28 y 37°C y b) presencia o ausencia de co2.
Morfometría celular. A células no infectadas e infectadas de ambas líneas celulares (J774 y Lulo), tomadas de las monocapas confluentes, se les realizó un análisis morfométrico. Se capturaron imágenes de todas las condiciones experimentales, mediante una cámara Sony Hyper HAD adaptada a un microscopio de luz (Olympus CH30). Se digitalizaron aproximadamente 400 células por cada variable utilizada en los experimentos, con el fin de comparar el área, eje mayor y menor de las células seleccionadas. El proceso de medición se realizó con el software Image Pro-Plus (Media Cybernetics).
Microscopía electrónica de transmisión. Cultivos celulares de Lulo, contenidos en cajas de 35 cm2, se incubaron a 28°C, en ausencia de co2 y se infectaron con promastigotes de L. chagasi . Los cultivos celulares infectados se fijaron con glutaraldehído fresco al 3% disuelto en buffer fosfato 0,1 M con un pH 7,2, durante una hora a 4°C.
Luego, se lavaron por cinco minutos tres veces inmersos en el anterior buffer y fueron post-fijadas por una hora con tetróxido de osmio al 1%. A continuación, se lavaron de nuevo, fueron deshidratadas en una serie ascendente de alcoholes y embebidas en Epon (Polyscience®). Se obtuvieron secciones ultradelgadas usando un ultramicrótomo y se tiñeron con acetato de uranil y citrato de plomo. Las muestras se examinaron en un microscopio electrónico de transmisión marca Zeiss EM 110 (Almeida de Faria et al 1999).
Análisis Estadístico. A los datos de las infecciones se les hizo una transformación angular (arcoseno de la raíz cuadrada de la proporción) y fueron sometidos a un test de ANOVA de una vía y a una prueba de comparaciones múltiples de Student Newman-Keuls, usando el programa Simstat para Windows (Versión 1.31). Las proporciones de infección a 28 y 37°C y los valores obtenidos con y sin atmósfera de co2, se compararon usando la prueba t de Student. La variable de área celular se analizó usando un test Kolmogorov-Smirnov (Young 1977).
Resultados
Infección celular. Se registró infección de las células con el parásito en todos los ensayos. Este se observó en forma de amastigote en las vacuolas parasitóforas, lo cual indica que previamente hubo internalización de los promastigotes, conversión a las formas amastigotes y multiplicación intravacuolar. Sin embargo, la intensidad de este proceso dependió de las variables utilizadas. Hubo resistencia de los promastigotes para desprenderse de las células, al realizarse los lavados con solución salina 24 horas post-infección, indicativo de fuerte adherencia de los parásitos a la membrana celular. En el control positivo con células J774 fue evidente el proceso infectivo, mediante el registro de amastigotes en las vacuolas parasitóforas. En la figura 1a se muestra una célula Lulo con amastigotes intracelulares.

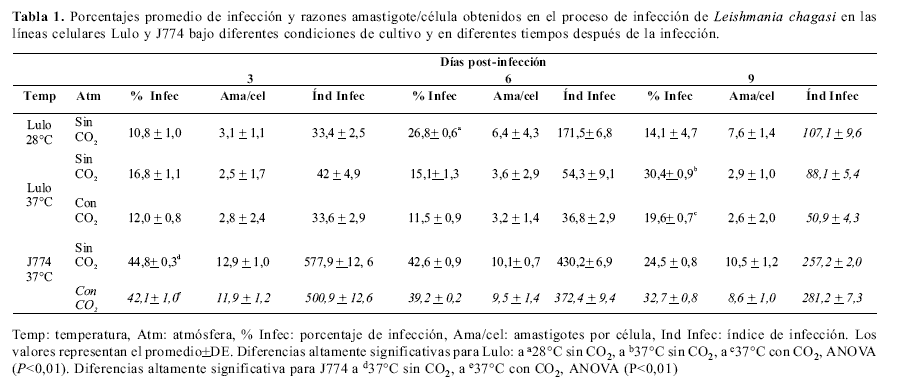
Porcentaje de infección. El día seis se registró el mayor porcentaje de infección (26,8%), en la línea celular Lulo infectada y mantenida a 28°C sin atmósfera de co2, mientras que en los días tres y nueve, los valores correspondieron a 10,8% y 14,1%, respectivamente. A 37°C sin atmósfera de co2, para los días post-infección tres y seis se determinaron porcentajes relativamente altos de infección (16,8% y 15,1% respectivamente); sin embargo, el día nueve mostró el mayor registro de infección (30,4%). El mayor porcentaje de infección (19,6%) en las células Lulo a 37°C con una atmósfera de 5% de co2 se registró el día nueve.
Los experimentos con el control positivo de la infección (células J774), en ausencia de co2, mostraron que a 37°C hubo mayor infección celular en el día tres (44,8%). De forma similar, con 5% de co2 el mayor porcentaje de infección (42,8%) ocurrió al tercer día, presentándose una ligera disminución a medida que el período post-infección se prolongó. El test Student del porcentaje promedio de infección indicó diferencias entre las células Lulo infectadas a 28°C (17,2%) y las células infectadas a 37°C (20,8%) sin atmósfera de co2 (P=0,0002). Igualmente, se encontraron diferencias entre las células Lulo infectadas a 37 °C (20,7%) y los resultados a la misma temperatura de incubación con 5% de co2 (14,4%) (P=0,0000002) (Table 1).
Número de amastigotes por célula. En la línea celular Lulo a 28°C, observamos que al avanzar en los días de infección aumentó progresivamente el número de amastigotes por célula, con el valor promedio más alto (7,6) el día nueve. Contrariamente, en las células Lulo a 37°C sin co2 se registraron 3,6 amastigotes/célula en el día seis; mientras que para los días tres y nueve post-infección, correspondieron a valores de 2,5 y 2,9 respectivamente. Cuando las células Lulo se incubaron en una atmósfera de 5% de co2 el número fue de 3,6 amastigotes/célula, para el día seis al contrario de lo que ocurrió el día nueve, cuyo valor fue el más bajo (2,6). En el control positivo de la infección a 37°C, con y sin atmósfera de co2, se observó mayor registro de parásitos por célula para el día tres; encontrándose 11,9 para el caso de células incubadas con co2 y 12,9 para las cultivadas sin co2.Índice de infección. El mayor índice de infección se registró en la línea celular J774 con un promedio (calculado con los datos de los tres días) de 413,9 en ausencia de co2; en la línea celular Lulo a 28°C se obtuvo un promedio de 104,5, siendo más alto en comparación con Lulo a 37°C con atmósfera de co2 (61,5) y sin atmósfera de co2 (40,4) (Tabla 1).
Morfometría celular. Las mediciones de área celular para la línea Lulo, evidenciaron el pleomorfismo de esta línea, pues las células se agruparon principalmente en tres rangos de área, las más numerosas, pequeñas (menores de 50 µm2), las intermedias (con áreas entre 51 y 90 µm2) y las grandes (con áreas mayores a 90 µm2) (Fig. 2A barras grises). Se comparó entre las células infectadas en las diferentes condiciones y se registró un aumento significativo de la proporción de células pequeñas infectadas a 37 °C con co2 (65,1%) con respecto a los controles de infección sin co2 a 28 °C (54,8%) (Test de Kolmogorov-Smirnoff, p<0,05). El aumento de la infección de células pequeñas ocurrió al tiempo que hubo una disminución en el porcentaje de células grandes infectadas (34,8% en el control de infección vs 21,8% en las células Lulo infectadas a 37 °C en presencia de co2) (Fig. 2A).

En las células J774 se observa que la infección induce un aumento en el área, pues la mayoría de las células en los cultivos no infectados tuvieron áreas entre 16-30 µm2 (48,5%), según las distribuciones calculadas con base en el tamaño celular, y esta población virtualmente desaparece después de la infección. Se observó un aumento en el porcentaje de las poblaciones celulares con un área entre 51 y 90 µm2 en los cultivos infectados. Como se observa en la Figura 2B (barras blancas y grises), la interacción con el parásito produjo un aumento significativo en las células con áreas mayores a 110 µm2, pues a pesar de que la población no infectada en este rango es de 12,0%, se encuentran grandes proporciones de células infectadas, en ausencia (52,1%) o en presencia de co2 (36,2%) en el mismo rango.
Microscopía electrónica de transmisión. Se observó la infección y localización del parásito L. chagasi en las líneas celulares seleccionadas, con clara definición de detalles ultraestructurales, y óptima conservación morfológica. También, se registró la integridad morfológica del parásito en el momento del contacto con las células. Se hizo patente la transformación del parásito L. chagasi en la línea celular Lulo, mediante la observación de amastigotes intracelulares (Fig.1b), demostrando que las células Lulo son un sustrato eficiente para el desarrollo del ciclo biológico del patógeno. En la línea celular J774 se encontraron células con numerosas vacuolas, mitocondrias y elementos organelares característicos y en estrecho contacto con células vecinas a través de sus procesos citoplasmáticos. También, se presentaron amastigotes intracelulares del parásito, confirmándose el proceso de infección.
Discusión
En el presente trabajo se demuestra por primera vez el proceso infectivo en presencia de variables de temperatura y co2 y con repercusiones a nivel de la morfología celular. Los parásitos tuvieron una fuerte adhesión a las células Lulo, independientemente de la variable analizada, demostrando la afinidad entre el parásito y la célula hospedera, lo cual permite inferir que ésta puede presentar receptores de membrana que facilitaron la unión, para luego dar paso a la internalización del parásito y la subsiguiente conversión de la forma promastigote a la amastigote. Además, hay que tener en cuenta la capacidad pluripotencial que probablemente desarrollaron muchas células Lulo para lograr la diferenciación en endocíticas y así poder efectuar el englobamiento de los parásitos. Esta situación, bajo otras circunstancias experimentales, es similar a la observado por Souza et al.(1977), quienes afirmaron que en el proceso endocítico potencial pudieron haber influido factores como el polimorfismo, componentes de membrana y carga eléctrica.
Las células Lulo presentaron capacidad para interactuar con los parásitos, originando modificaciones durante el desarrollo celular con posterior alojamiento de los parásitos; como lo muestra la presencia de vacuolas y formas amastigotas en su interior. Además, se han descrito partículas similares a carbohidratos alrededor de las membranas celulares de los parásitos, identificadas mediante marcadores específicos por microscopía electrónica, las cuales pueden ser considerados favorables al proceso de interacción de los parásitos con las células (Sacks y Kamhawi 2001), similar a como ocurre en el intestino medio del flebótomo.
A 28°C sin co2 se incrementó el porcentaje e índice de infección en las células Lulo del día tres al día seis, disminuyeron ambos parámetros para el día nueve. Esta situación es similar a la reportada por Bello et al. (2005), quienes determinaron incrementos en estos mismos parámetros de infectividad desde el día dos hasta el día seis y luego disminuyeron paulatinamente hasta el día 10 post-infección. Por el contrario, a 37°C en ausencia o presencia de co2, los cultivos tuvieron un alto porcentaje e índice de infección el día tres. La situación de las células J774, mostró alto porcentaje de infección el día tres, comparado con las células Lulo, lo cual permite inferir que este indicador de infección en los días anteriores venía en aumento e incluso se pudo mantener en ese ritmo ascendente hasta el día cinco, disminuyendo ligeramente el día seis, para luego descender hasta el día nueve, donde se registraron los valores más bajos. El efecto del tiempo de infección del parásito en las células J774 coincide parcialmente con los resultados de Bosque et al.(1998), quienes registraron incrementos del índice de infección de L. panamensis en macrófagos derivados de monocitos, en los tiempos de una, 24 y 48 horas post-infección y luego, a las 72 horas el valor descendió. Sin embargo, en este mismo trabajo aumentó el porcentaje de células infectadas y el número de parásitos por célula en fagocitos mononucleares a través de todos los tiempos de exposición tomados en la evaluación del proceso infectivo (dos, 24, 48, 72 y 120 hr).
Lo observado a 37°C sin atmósfera de co2 durante los ensayos de infección de L. chagasi en las células Lulo demostró que, selectivamente, muchas de estas células al multiplicarse, permitieron la internalización y replicación de los parásitos aún con los porcentajes de infección más altos a los registrados a 28°C, presentándose en los análisis estadísticos diferencias significativas. Esto demuestra la capacidad de las células Lulo como hospederas del parásito, en condiciones adversas de temperatura, pero, al mismo tiempo, favorables a los amastigotes si tenemos en cuenta que el promedio de la temperatura en la vacuola parasitófora del hospedero mamífero se encuentra en este valor. Los experimentos en cultivos de Lulo con atmósfera de co2 a 37°C, mostraron valores de infección estadísticamente menores a lo registrado en ausencia de co2.
La alta infección del parásito en las células Lulo a 28°C, posiblemente se debe a que el medio de cultivo de los insectos posee un pH ligeramente ácido con respecto a otros medios de vertebrados y, en ausencia de una atmósfera de co2, éste desciende a niveles críticos de acidez, lo cual favorece la infectividad de los parásitos, siendo concordante con lo reportado por Zilberstein y Shapira (1994), estos autores sometieron promastigotes y amastigotes de Leishmania sp a diferentes pH y temperaturas, encontrando que los parásitos se adaptaron mejor a pH bajos y a temperaturas relativamente altas. El metabolismo del amastigote está adaptado a un pH ácido (Glaser et al 1988) y estos organismos pueden atravesar el gradiente de protones a través de su membrana, formada sobre condiciones acídicas para conducir el transporte activo de glucosa y aminoácidos (Zilberstein 1991; Glaser et al 1988). Este mecanismo puede contribuir al mantenimiento del pH ácido en la vacuola parasitófora.
Con respecto al control positivo del estudio, la línea celular J774, los porcentajes, índice de infección y número de amastigotes por célula fueron más altos comparados con las células Lulo, debido a que estas células presentan características macrofágicas (Papadimitriou y Ashman 1989; Ralph et al 1975) y se encuentran adaptadas fisiológicamente como hospederas del parásito.
Según el análisis morfométrico, la temperatura de incubación no afectó los porcentajes de células no infectadas en cada rango (barra gris en la figura 2). De igual manera, las células infectadas e incubadas a 37 °C en ausencia de co2, no cambiaron los perfiles de área con respecto a los controles (p>0,05), como si ocurrió al incubar las células infectadas a 28°C, cultivos en los cuales se aumentó significativamente el grupo de células mayores de 90 µm2, con una disminución consecuente en los grupos de células pequeñas e intermedias. El mayor cambio en la morfología ocurrió cuando se incubaron las células Lulo infectadas a 37°C con atmósfera de co2, pues se encontró un aumento significativo en el porcentaje de células pequeñas (16-50 µm2), al tiempo que una disminución de las células grandes (mayores de 90 µm2); este hallazgo estuvo relacionado con un mayor número de amastigotes intracelulares. Se podría pensar que la presencia de co2 favoreció la infección en la célula y la capacidad de alojar los parásitos de L. chagasi . La replicación y transformación in vitro de parásitos de Leishmania en las células Lulo y en las J774, originaron cambios en la morfología celular, lo cual finalmente podría haber terminado en lísis, fenómeno que se observa en la infección natural in vivo de Leishmania en las células del hospedero. Esto concuerda con lo reportado en otros estudios quienes comprobaron que la replicación de patógenos en células de mamífero ocasiona cambios en su morfología (Carrasco 1987; Páez y Estebán 1987; Ulug et al.1987; Carrasco et al.1989).
En los cultivos de J774, la infección produjo un aumento en el área de las células, las poblaciones de células más pequeñas desaparecieron y se incrementó el porcentaje de células intermedias y grandes. La población de células mayores a 110 µm2, fue cinco y tres veces mayor en ausencia o presencia de co2 respectivamente, indicando que el parásito podría estar induciendo agravamientos celulares o sincitios.
En el desarrollo del proceso de infección, las células experimentaron aumento en su tamaño hasta alcanzar áreas superiores a 111 µm2, en donde se ubicó el mayor porcentaje de células. Este aumento en el tamaño celular, probablemente, se debió a los cambios metabólicos que indujo el parásito al internalizarse y transformarse en amastigote.
Los cultivos celulares derivados de L. longipalpis permitieron la adhesión, internalización y transformación iológica de los parásitos de L. chagasi . Probablemente muchas células Lulo pudieron expresar su capacidad pluripotencial, como células embrionarias no diferenciadas, al convertirse en hospederas del patógeno configurándose en una herramienta útil de investigación para reproducir algunos procesos de infección del parásito que eventualmente pueden ocurrir en las interacciones parásito-hospedero-vector. Esto puede apoyar estrategias eficaces en el control del parásito.
Agradecimientos
A la doctora María Leonor Caldas y Ladys Sarmiento del laboratorio de análisis y procesamiento de imágenes del Instituto Nacional de Salud, por el apoyo en el procesamiento de las muestras con la microscopía electrónica de transmisión. En la realización del trabajo no se presentó conflicto de interés financiero, político ni académico. Este trabajo fue financiado por Colciencias, Universidad de La Salle e Instituto Nacional de Salud (código: 1243-05-12416).
Literatura citada
ABDULRAHMAN, YI.; GARMSON, IJ.; MOLYNEUX, DH.; BATES, PA. 1998. Transformation, development, and transmission of axenically cultured amastigotes of Leishmania mexicana in vitro and in Lutzomyia longipalpisi . American Journal of Tropical Medicine and Hygiene (59): 421-25. [ Links ]
AIKAWA, M.; HENDRICKS, LD.; ITO, Y.; JAGUSIAK, M. 1982. Interactions between macrophagelike cells and Leishmania braziliensis in vitro. American Journal of Pathology (108): 50-9. [ Links ]
ALMEIDA DE FARIA, M.; FREYMULLER, E.; COLLI, W.; ALVES, MJ. 1999. Trypanosoma cruzi: characterization of an intracellular epimastigote-like form. Experimental Parasitology (92):263-274. [ Links ]
AÑEZ, N.; TANG, Y.; ROJAS, A.; CRISANTE, G.; KILLICK-KENDRICK, M.; KILLICK-KENDRICK, R. 2003. Detection of amastigote-like forms in the valve of Phlebotomus papatasi infected with Leishmania major. Memorias do Instituto Oswaldo Cruz (98): 495-98. [ Links ]
AQUINO, MC.; SANTOS, R.; BARRETO, R.; PONTES DE CARVALHO DOS SANTOS, WLC. 2002. A simple and reproducible method to obtain large numbers of axenic amastigotes of different Leishmania species. Parasitology Research (88): 963-8. [ Links ]
BATES, PA.; ROBERTSON, CD.; TETLEY, L.; COOMBS, GH. 1992. Axenic cultivation and characterization of Leishmania mexicana amastigotes-like forms. Parasitology (105): 193-202. [ Links ]
BATES, PA.; TETLEY, L. 1993. Leishmania mexicana: induction of metacyclogenesis by cultivation of promastigotes at acidic pH. Experimental Parasitology (76): 412-23 [ Links ]
BATES, PA. 1994. Complete developmental cycle of Leishmania mexicana in axenic culture. Parasitology (108): 1-9. [ Links ]
BATES, PA. 1997. Infection of Phlebotomine sandflies with Leishmania. En: JM Crampton, CB Beard, C Louis (eds), The Molecular Biology of Inscect Disease Vectors, Chapman & Hall, London. 112-120 p. [ Links ]
BELLO, FJ.; MEJÍA, AJ.; CORENA, M.; AYALA, M.; SARMIENTO, L.; ZUÑIGA, C.; PALAU, M. 2005. Experimental infection of Leishmania (L) chagasi in a cell line derived from Lutzomyia longipalpisi (Diptera: Psychodidae). Memorias do Instituto Oswaldo Cruz (100): 519-525. [ Links ]
BOSQUE, F.; GENEVIEVE, M.; VALDERRAMA, L.; SARAVIA, N. 1998. Permissiveness of human monocytes and monocyte-derived macrophages to infection by promastigotes of Leishmania (Viannia) panamensis. Journal of Parasitology (84): 1250-6. [ Links ]
CARRASCO, I. 1987. Animal virus induced cytotoxity: an overview, p. 1-3. En L. Carrasco, Mecanism of viral toxicity in animal cells, CRC Press, Boca Raton, FL. 19. [ Links ]
CARRASCO, I.; OTERO, MJ.; CASTRILLO, JL. 1989. Modification of membrane permeability by virus infection. Pharmacology & Therapeutics (40): 171-212. [ Links ]
CHANG, KP.; DWYER, DM. 1976. Multiplication of a human parasite (Leishmania donovani) in phagolysosomes of hamster macrophage in vitro. Science (193): 678-80. [ Links ]
CHANG, KP. 1978. Leishmania infection of human skin fibroblasts in vitro: absence of phagolysosomal fusion after induced phagocytosis of promastigotes, and their intracellular transformation. American Journal of Tropical Medicine and Hygiene (27): 1084-96. [ Links ]
CHANG, KP. 1979. Leishmania donovani: Promastigote-macrophage surface interactions in vitro. Experimental Parasitology (48): 175-89. [ Links ]
CORTE-REAL, S.; SANTOS, CB.; MEIRELLSES, MNL. 1995. Differential expression of the plasma membrane Mg2+ ATPase and Ca2+ ATPase activity during adhesion and interiorization of Leishmania amazonensis in fibroblasts in vitro. Journal of Submicroscopic Cytology and Pathology (27): 359-66. [ Links ]
CRAMPTON, JM.; BEARD, CB.; LOUIS, C. 1997. The molecular biology of insect disease vector. A methods manual. 1th Edition. Chapman & Hall. London. 578 p. [ Links ]
CUNHA, AM.; CHAGAS, E. 1937. Nova espécie de protozoário do Gênero Leishmania pathogênico para o homem (nota prévia). Hospital (Rio de Janeiro) (11): 5-9. [ Links ]
DEDET, JP.; RYTER, A.; VOGT, E.; HOSU, P.; PEREIRA DA SILVA, L. 1983. Uptake and killing of Leishmania mexicana amazonensis amastigotes by human skin fibroblasts. Annals of Tropical Medicine and Parasitology (77): 35-44. [ Links ]
FAMPA, P.; CORREA DA SILVA, MS.; LIMA, DC.; OLIVEIRA, SM.; MOTTA, MC.; SARAIVA, E. 2003. Interaction of insect trypanosomatids with mosquitoes, sand fly and the respective insect cell lines. International Journal for Parasitology (33):1019-26. [ Links ]
GLASER, TA.; BAATZ, JE.; KREISHMAN, GP.; MUKKADA, AJ. 1988. pH homeostasis in Leishmania donovani amastigotes and promastigotes. Proceedings of the National Academy of Sciences of the United States of America (85): 7602-6. [ Links ]
HÉRVAS-RODRIGUEZ, J.; MOZOS, E.; MÉNDEZ, A.; PÉREZ, J.; GÓMEZ-VILLAMANDOS, JC. 1996. The Leishmania infection of canine skin fibroblasts in vitro. Veterinary Pathology (33): 469-73. [ Links ]
HODGKINSON, VH.; SOONG, L.; DUBOISE, SM.; MCMAHON-PRATT, D. 1996. Leishmania amazonensis: cultivation and characterization of axenic amastigote-like organisms. Experimental Parasitology (83): 94-105. [ Links ]
IGARASHI, AJ. 1978. Isolation of a Singh’s Aedes albopictus cell clones sensitive to dengue and chikungunya viruses. Journal of General Virology (40):531-44. [ Links ]
IGARASHI, AJ. 1985. Mosquito cell culture and study of Arthropod-Borne Togaviruses. Advances in Virus Research (30): 21-42. [ Links ]
KITAMURA, S. 1965. The in vitro cultivation of tissues from the mosquito Culex pipiens var. molestus. II. A improved culture medium useful for ovarian tissue culture. Kobe Journal of Medical Sciences (11): 315 – 433. [ Links ]
KILLICK-KENDRICK, R. 1986. The transmission of Leishmaniasis by the bite of the sandfly. Journal of the Royal Army Medical Corps (132): 134-40. [ Links ]
KUNO, G.; GUBLER, DJ.; VELEZ, M.; OLIVER, A. 1985. Comparative sensitivity of three mosquito cell line for isolation of dengue viruses. Bulletin of the World Health Organization (63): 279 – 86. [ Links ]
LEAKE, CJ. 1997. Establishing primary cell culture from disease vectors and maintenance of continuous cell line. En: JM Crampton, CB Beard, C Louis (eds), The Molecular Biology of Insect Disease Vectors, Chapman & Hall, London. 487 p. [ Links ]
LEÓN, LL.; SOARES, MJ.; TEMPORAL, RM. 1995. Effects of temperature on promastigotes of several species of Leishmania. Journal of Eukaryotic Microbiology (42): 219-223. [ Links ]
LONARDONI, MVC.; RUSSO, M.; JANCAR, S. 2000. Essential role of platelet-activating factor in control of Leishmania (Leishmania) amazonensis infection. Infection and Immunity (68): 6355-61. [ Links ]
LUTZ, A.; NEIVA, A. 1913. Contribuicoes para a biolojia das megarininas com descricoes de duas especies novas. Memorias do Instituto Oswaldo Cruz (5): 129-41. [ Links ]
MATTOCK, NM.; PETTERS, W. 1975. The experimental chemotherapy of Leishmaniasis. I: Techniques for the study of drug action in tissue culture. Annals of Tropical Medicine and Parasitology (69): 349-56. [ Links ]
MOLYNEUX, DH.; KILLICK-KENDRICK, R. 1986. Morphology, ultrastructure and life cycles in The Leishmaniases. En: Peters, W.; Killick-Kendrick, R. (Eds.), Biology and Medicine, vol. I. Academic Press, London .140–161 p. [ Links ]
PAN, AA. 1984. Leishmania mexicana. Serial cultivation of intracellular stages in a cell-free medium. Experimental Parasitology (58): 72-80. [ Links ]
PAEZ, E.; ESTEBÁN, M. 1987. Inhibition of host protein synthesis by DNA viruses. 59-89 p. In Carrasco, L. 1987. Mechanism of viral toxicity in animal cells, CRC, Press, Boca Raton, Florida. [ Links ]
PAPADIMITRIOU, JM.; ASHMAN, RB. 1989. Macrophages: current views on their differentiation, structure, and function. Ultrastructural Pathology (13): 343-372. [ Links ]
PEARSON, RD.; ROMITO, R.; SYMES, PH.; HARCUS, JL. 1981. Interaction of Leishmania donovani promastigotes with human monocyte-derived macrophages: parasite entry, intracellular survival, and multiplication. Infection and Immunity (32): 1249-53. [ Links ]
PESSOTTI, JH.; ZAVERUCHA DO VALLE, T.; CORTE-REAL, S.; GONCALVES DA COSTA, SC. 2004. Interaction of Leishmania (L) chagasi with the Vero cell line. Parasite (11): 99-102. [ Links ]
RALPH, P.; PRICHARD, J.; COHN, M. 1975. Reticulum cell sarcoma: an effector cell in antibody-dependent cell-mediated immunity. The Journal of Immunology (114): 898-905. [ Links ]
REY, G.; FERRO, C.; BELLO, F. 2000. Establishment and characterization of a New Continuous cell line from Lutzomyia longipalpisi (Diptera: Psychodidae) and its susceptibility to infections with Arboviruses and Leishmania chagasi . Memorias do Instituto Oswaldo Cruz (95): 103-10. [ Links ]
SAAR, Y.; RANSFORD, A.; WALDMAN, E.; MAZAREB, S.; AMIN-SPECTOR, S.; PLUMBLEE, J.; et al. 1998. Characterization of developmentally-regulated activities in axenic amastigotes of Leishmania donovani. Molecular and Biochemical Parasitology (95): 9-20. [ Links ]
SACKS, DL.; KAMHAWI, S. 2001. Molecular aspects of parasite – vector and vector – host interactions in Leishmaniasis. Annual Review Microbiology (55), 453-83. [ Links ]
SINGH, K. 1967. Cell culture derived from larvae of Aedes albopictus (Skuse) and Aedes aegypti (L). Current Science (36): 506 – 8. [ Links ]
SCHWARTZMAN, JD.; PEARSON, RD. 1985. The interactions of Leishmania donovani promastigotes and human fibroblasts in vitro. American Journal of Tropical Medicine and Hygiene (34): 850-55. [ Links ]
SOUZA, WC.; ARGUELLO, A.; MARTINEZ–PALOMO, D.; GONZALEZ–ROBLES, T.; CHIAU, E. 1977. Surface charge of Trypanosome cruzi. Binding of cationized ferritin and measurement of cellular electrophoretic mobility. Journal of Protozoology (24): 411-15. [ Links ]
ULUG, ET.; GARRY, RF.; BOSE, HR. 1987. Cell killing by enveloped RNA viruses, 92-113 p. In Carrasco, L. Mechanisms of viral toxicity in animal cells, CRC Press, Boca Raton, FL. [ Links ]
YOUNG, IT. 1977. Proof without prejudice, use of the Kolmogorov, Smirnov test for the analysis of histograms from floe systems and other sources. Journal of Histochemistry and Cytochemistry (25):935-41. [ Links ]
ZILBERSTEIN, DA. 1991. Adaptation of Leishmania species to an acidic environment. En: Biochemical Protozoology (G. Coombs & M. North, eds) London: Taylor and Francis. 349-358 p. [ Links ]
ZILBERSTEIN, DA.; SHAPIRA, M. 1994. The role of pH and temperature in the development of parasites. Annual Review of Microbiology (48): 449 470. [ Links ]
Recibido: 29-mar-06 • Aceptado: 16-sept-06


{kind=link}