










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488On-line version ISSN 2665-4385
Rev. Colomb. Entomol. vol.33 no.1 Bogotá Jan./June 2007
Sección Agrícola
Tablas de vida de Hypothenemus hampei (Coleoptera: Curculionidae: Scolytinae) sobre tres introducciones de café
Life tables of Hypothenemus hampei (Coleoptera: Curculionidae: Scolytinae) on three coffee accessions
Juan Vicente Romero1, Hernando Al fonso Cortina G.2
1 I. A., Disciplina de Mejoramiento Genético, CENICAF É. juanvicente.romero@cafedecolombia.com
2 Autor para correspondencia: I.A. M.Sc., Disciplina de Mejoramiento Genético, CENICAF É. A.A. 2427 Manizales. hernando.cortina@cafedecolombia.com
Resumen. Se elaboraron y compararon las tablas de vida de Hypothenemus hampei sobre las introducciones de café Coffea liberica, Coffea arabica CCC 534 y la variedad susceptible Caturra, con el fin de hallar fuentes de resistencia a este insecto bajo condiciones controladas (26ºC ± 1, 75% ± 5 H. R.). Se realizaron dos experimentos: en el primero, se siguió el desarrollo de los estados inmaduros y se estimó la supervivencia. En el segundo, se determinó la supervivencia y fecundidad de los adultos usando hembras de 36 días de edad. La duración del ciclo para CCC 534, C. liberica y Caturra fue similar: 20 días en total, huevo cuatro, primer instar cuatro, segundo instar seis, prepupa dos y pupa cuatro días, aproximadamente. El número de huevos acumulados en las tres introducciones se ajustó a funciones cuadráticas. Los intervalos de confianza (95%), mostraron diferencias significativas entre las introducciones y el testigo: CCC 534 32 ± 2, C. liberica 28 ± 2 y Caturra 42 ± 2 huevos por hembra. Las proporciones sexuales fueron de 1: 9,2, 9,8 y 9,8 macho: hembra, respectivamente. No se encontraron diferencias en la supervivencia (entre 58 y 63%) hasta los 72 días para las tres introducciones. Hubo diferencias en la tasa reproductiva neta (Ro) entre Caturra (25 ± 1 huevos), CCC 534 (18 ± 2) y C. liberica (15 ± 2), en la tasa intrínseca de crecimiento (r) (0,073 ± 0,001; 0,065 ± 0,002; 0,057 ± 0,003, respectivamente) y en el tiempo de duplicación (10 ± 0,2; 11 ± 0,4; 12 ± 0,6 días); en el tiempo generacional solo hubo diferencias entre Caturra y C. liberica (45 ± 0,4; 47 ± 0,8, respectivamente).
Palabras clave. Germoplasma de café. Resistencia a insectos. Antibiosis.
Abstract. The life tables of Hypothenemus hampei were elaborated and compared on the coffee accessions Coffea liberica, Coffea arabica CCC 534 and the susceptible variety Caturra, in order to find sources of resistance to this insect under controlled conditions (26ºC ± 1, 75% ± 5 R. H.). Two assays were performed: in the first, the development of the immature stages was followed and survival was estimated. In the second, adult survival and fecundity were determined using 36- day old females. The duration of the cycle for CCC 534, C. liberica and Caturra was similar: total 20 days, egg four, first instar four, second instar six, prepupa two and pupa four days, approximately. The number of accumulated eggs in the three accessions fit quadratic functions. The confidence intervals (95%) showed significant differences between the accessions and the control: CCC 534 32 ± 2, C. liberica 28 ± 2 and Caturra 42 ± 2 eggs per female. The sex ratios were 9.2, 9.8, and 9.8 male: female, respectively. There was no difference in survival (range 58-63%) up to 72 days for the three accessions. There were differences in the net reproductive rate (Ro) among Caturra (25 ± 1 eggs), CCC 534 (18 ± 2), and C. liberica (15 ± 2); in the intrinsic rate of increase (r) (0.073 ± 0.001, 0.065 ± 0.002, 0.057 ± 0.003, respectively), and the duplication time (10 ± 0.2, 11 ± 0.4, 12 ± 0.6 days). In the generation time there was only a differences between Caturra and C. liberica (45 ± 0.4, 47 ± 0.8, respectively).
Key words. Coffee germplasm. Resistance to insects. Antibiosis.
Introducción
Colombia es el primer productor mundial de café arábigo lavado, con aproximadamente 850.000 hectáreas sembradas. El café contribuyó con el 12% del producto interno bruto agropecuario del país en 2004 (MAD R 2006), y su cosecha generó en 2005 $3.2 billones (Silva 2006). El problema fitosanitario más importante de este cultivo en el país, desde 1988, es la broca Hypothenemus hampei (Ferrari, 1867) (Coleoptera: Curculionidae: Scolytinae), que produce severos daños a la caficultura en más de 700.000 hectáreas (Fedecafé 1999), por la reducción de la producción, la alteración de la calidad, el aumento de los costos de producción, se estima en 7% el costo del control de la broca (Duque et al. 1997), y la disminución del precio de compra.
Debido a la distribución de la cosecha en Colombia, la broca encuentra frutos durante todo el año, en la mayor parte de la región cafetera (Arcila et al. 1993). Una vez en el fruto, penetra, realiza galerías, oviposita y permanece en su interior mientras que su progenie se desarrolla, lo que complica su manejo. Actualmente se realizan controles químicos, biológicos y principalmente culturales (Bustillo 2002).
La resistencia varietal es otra herramienta para el control de insectos, que disminuye la población de la plaga al alargar su ciclo de vida y reducir la supervivencia de los individuos o su capacidad reproductiva (Adkinsson y Dyck 1984). A pesar que dentro del género Coffea no se conocen genotipos resistentes a la broca, trabajos previos (Koch 1973; Villagran 1991; Duarte 1992), han mostrado diferencias entre algunos, que aunque pequeñas, pueden ser de interés en un programa de control integrado de la plaga. Romero y Cortina (2004), evaluando la colección colombiana de café, criaron brocas en 18 introducciones etíopes de Coffea arabica L. y una de Coffea liberica Bull ex Hiern., encontrando que cuatro introducciones tuvieron significativamente menos individuos que el testigo, la variedad Caturra.
En este trabajo con el fin de identificar posibles fuentes de resistencia a H. hampei, se compararon la fertilidad y la supervivencia de la broca, al ser criada sobre dos introducciones destacadas y sobre Caturra. Se estudió el crecimiento poblacional utilizando las tablas de vida, que permiten conocer y comparar los principales parámetros poblacionales o demográficos de la broca. Este procedimiento ha sido usado para la identificación de fuentes de resistencia a insectos, especialmente por antibiosis (Trichilo y Leigh 1985; Cardona et al. 1989; Soroka y Mackay 1991).
Materiales y Métodos
En el Centro Nacional de Investigaciones de Café, Cenicafé (Chinchiná, Caldas), en un cuarto con 26ºC ± 1, 75% ± 5 de humedad relativa y con períodos de 12 horas luz artificial (lámparas de 48 vatios, luz día), se evaluaron tres introducciones del género Coffea: una de C. liberica, diploide autoincompatible, y dos de C. arabica, tetraploide autocompatible, la accesión CCC 534 recolectada por la misión de la FAO de 1964-1965 (Meyer 1968), y la variedad susceptible Caturra, que ocupa aproximadamente el 40% del área sembrada en café en Colombia (Fedecafé 1997).
Se realizaron dos experimentos: en el primero se siguió el desarrollo de los estados inmaduros y se estimó su supervivencia; en el segundo se determinó la supervivencia y fecundidad de los adultos a partir de hembras de 36 días de edad.
Evaluación de inmaduros. Se establecieron tres cohortes de 400 huevos de uno a cuatro días de edad, puestos por hembras criadas en las mismas introducciones. La unidad de muestreo (UM), fue un grano de café pergamino con 40% de humedad y un orificio de 1 mm de diámetro y 7 mm de profundidad, donde se colocaron dos huevos (Fig. 1), el grano se depositó en un vial de borosilicato de 9 mm de diámetro por 34 mm de alto. Los granos se trataron con tiabendazol al 1%. La perforación se realizó una con una broca de 1/32”, la cual se esterilizó con flameos frecuentes.
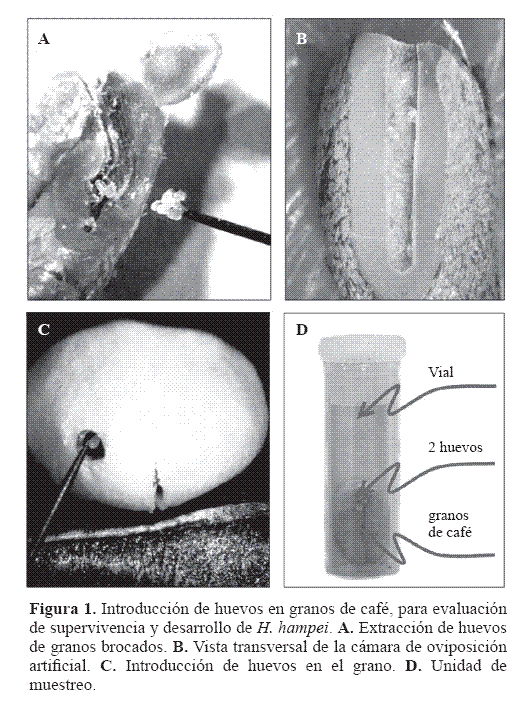
Cada cuatro días, hasta completar 10 evaluaciones (a los 40 días), se tomaron aleatoriamente 20 granos por introducción y se disecaron para determinar las brocas vivas y su estado de desarrollo. Para estimar la duración de cada uno de los estados inmaduros, se consideró que había cambio de estado, en la primera lectura en la que más del 50% de la población se encontraba en un estado más avanzado que el predominante en la evaluación anterior. A partir de la relación entre el número de individuos vivos y el inicial, se estimó la supervivencia, para cada intervalo de edad.
El principal factor no genético que afecta el desarrollo de la broca en el grano, es la humedad del endospermo (Fernie et al. 1986). Por tal razón, en cada evaluación, se determinó la humedad de una muestra de tres granos de cada una de las introducciones, se calculó la diferencia entre el peso de la muestra antes y después de ser secada en una estufa a 105°C durante 24 horas, se dividió entre el peso inicial de la muestra y se llevó a porcentaje. De acuerdo con Álvarez et al. (2001), la humedad del grano debe ser superior al 20% durante el experimento.
Evaluación de hembras adultas. Cohortes de 360 hembras adultas de aproximadamente 35 días, criadas en cada una de las introducciones, se ubicaron individualmente en viales con un grano de café pergamino de la respectiva introducción, en la cual se criaron (UM). La humedad de los granos fue de 40% aproximadamente. Cada cuatro días, durante nueve evaluaciones, se tomaron aleatoriamente 40 UM de cada introducción, y se registró: el número de hembras vivas y el número de huevos. Se midió la humedad de tres granos de cada introducción a través del tiempo.
Se estimaron las funciones de oviposición acumulada a través del tiempo en las tres introducciones y los intervalos de confianza para la media en cada tiempo de evaluación. Se calculó la proporción de sexos de la progenie de las hembras fundadoras, hallando la relación entre hembras y machos adultos encontrados en la última evaluación. Todos los ajustes de curva se realizaron con el programa estadístico SAS versión 8.0 (SAS 1999).
Como las supervivencias de inmaduros y adultos fueron estimadas en las mismas condiciones (75 ± 5% de humedad y 26 ± 1oC), se calcularon las funciones de supervivencia desde huevos hasta adultos a los 72 días, cuando ya había ocurrido la mayor parte de las posturas, y se construyeron los intervalos de confianza para la media en cada fecha de evaluación. Esta integración se usa especialmente cuando el seguimiento de los individuos es difícil o sus ciclos se encuentran traslapados (Harcourt 1967; Rabinovich 1980; Price 1997), como ocurre con la broca y sus hábitos crípticos en el grano de café
Parámetros demográficos. Con la supervivencia (lx), y la fecundidad especifica (mx= N° huevos x proporción sexual), por edad (x), se elaboraron las tablas de fecundidad de cada población y se calcularon los siguientes parámetros demográficos (Birch 1948; Andrewartha y Birch 1954):
Tasa reproductiva neta, que mide el efecto de la introducción sobre la reproducción de las hembras:
Tasa intrínseca de crecimiento, indica el efecto de la introducción sobre la capacidad innata de crecimiento de la población de broca. Inicialmente se calculó como:
Y este valor se tomó como base para resolver, por iteración, el valor de r en la ecuación de Lotka:
Luego se recalculó el tiempo generacional, que mide la influencia de la introducción sobre la edad promedio en la cual una broca tiene su descendencia:
Estos parámetros son puntuales y no hay un estimativo de sus errores típicos, lo que impide comparaciones estadísticas. Esta limitación se superó con la técnica de remuestreo de Jackknife, que permite realizar estimaciones de error y asociar intervalos de confianza a los parámetros demográficos. Para ello se formaron submuestras, ignorando por ciclos una de las observaciones de los datos originales. Para cada una de las submuestras, se calcularon los parámetros demográficos, y con su variabilidad se estimó el error estándar (Sokal y Rohlf 1995). Basado en esta técnica y con los algoritmos computacionales para las tablas de vida desarrollados por Maia et al. (2000) en lenguaje SAS, se calcularon los límites de confianza para los parámetros y se realizaron las pruebas de t.
Resultados y Discusión
Durante la evaluación de inmaduros y de hembras adultas, la humedad de los granos fue en general superior al 30% en todos los tiempos de evaluación; no hubo diferencia entre las introducciones. Al ser superior al control del 20% propuesto por Álvarez et al. (2001), para estos experimentos. Hay evidencia de que estas condiciones son apropiadas para el desarrollo de la broca.
Inmaduros. Álvarez et al. (2001), propusieron la evaluación de introducciones a partir del estudio de las crías de hembras adultas, de esta forma la supervivencia y ciclo de vida se estiman indirectamente a partir del número de posturas, el número total de individuos y el tiempo en que predomina un determinado estado. Aunque se puede estimar la mortalidad, principalmente de los estados inmaduros, ésta queda confundida por la natalidad que ocurre en diferentes días y no permite la construcción precisa de las tablas de vida.
Al adicionar el experimento del desarrollo de estados inmaduros a la metodología de Álvarez (2001), se logró una mejor estimación de la duración de los estados inmaduros y de la mortalidad, debido a que se conoce el momento de inicio del ciclo y la mortalidad en cada grano. Por otra parte, este experimento permite determinar en qué momento se estaría manifestando el efecto de antibiosis sobre el desarrollo.
El primer instar ocurrió aproximadamente a los cuatro días, y a los ocho predominaron las larvas del segundo instar (Tabla 1), en Caturra y en las dos introducciones. A los 12 se presentaron en Caturra larvas II, prepupas y pupas en proporciones similares, mientras que en las dos introducciones predominaron las larvas II. El estado de prepupa fue muy rápido, como lo habían indicado Bergamin (1943), Bartra et al. (1982) y Montoya (1993), por lo que la frecuencia de las evaluaciones no permitió identificar claramente su duración.
Más del 50% de individuos en el estado de pupa se presentó a los 16 días después de la infestación (DDI), en Caturra y en la introducción CCC 534, pero en C. liberica 32% de los individuos aún eran larva II. En esta fecha también aparecieron los primeros adultos no esclerotizados en todas las introducciones. Los adultos emergieron alrededor de los 20 DD I, y el 100% de adultos se presentaron a los 24 DDI en Caturra y CCC 534, mientras que en C. liberica ocurrió a los 32 días.
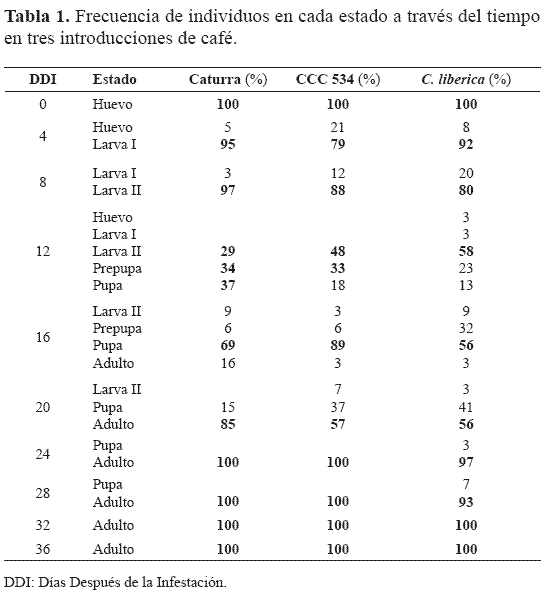
De acuerdo con esto, la duración del ciclo para CCC 534, C. liberica y Caturra fue similar: 20 días; los tiempos de desarrollo estimados son los siguientes: huevo cuatro días, primer instar cuatro días, segundo instar seis días, prepupa dos días y pupa cuatro días. Los cuales concuerdan con la duración hallada bajo condiciones similares por Bergamin (1943), Borbon (1989) y Montoya (1993).
Hembras adultas
Oviposición. Durante la evaluación de fecundidad, en los granos de las tres introducciones se presentaron dos períodos de oviposición (Fig. 2). El primero corresponde a la hembra fundadora, con un pico entre los ocho y 12 DD I, disminuyendo hasta los 24 DD I en Caturra y CCC 534, y hasta los 28 en C. liberica. Bergamin (1943), a 27°C, obtuvo resultados similares, encontrando la mayor oviposición entre siete y 13 DDI y una disminución de la intensidad de oviposición hasta los 27 DD I.
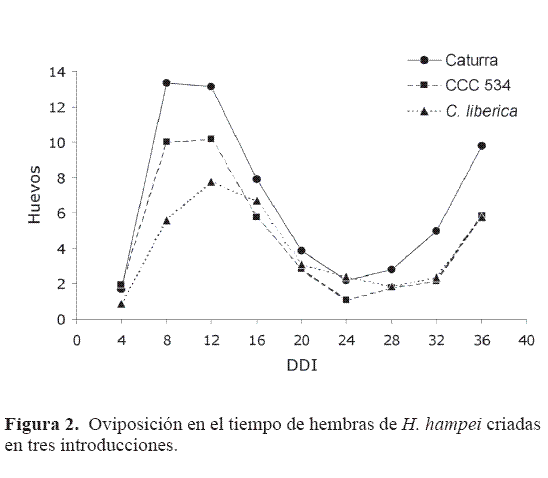
A unque es probable que las fundadoras hayan puesto algunos huevos después de los 24 DD I, para el análisis se tomó hasta esta fecha, debido que la nueva generación de adultos se desarrolló a partir de los 20 DDI, y como lo señalan Bergamin (1943) y Baker et al. (1992), el segundo periodo de oviposición corresponde principalmente a las hembras adultas recién emergidas, enmascarando las posturas finales de la fundadora.
La oviposición acumulada de las brocas fundadoras, hasta los 24 días en Caturra y CCC 534 y hasta los 28 DD I en C. liberica, se ajustaron a una funciones cuadráticas (Fig. 3); los intervalos de confianza del 95%, para la media en cada evaluación, mostraron diferencias significativas entre las introducciones y el testigo. La oviposición en las introducciones fue menor que en Caturra desde los ocho DD I, cuando comienza la máxima oviposición (Fig. 2).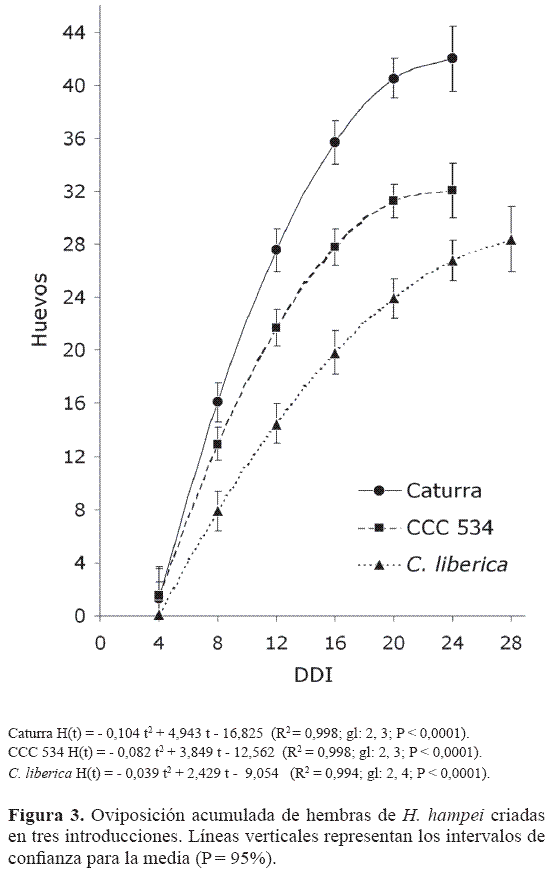
Sobre Caturra, hasta los 24 DD I, las hembras ovipositaron en promedio 42 huevos, Bergamin (1943) encontró 41.3 huevos a los 27 DD I, mientras que en CCC 534 y C. liberica se encontraron respectivamente 32 y 28 huevos por grano, lo que equivale a 24% y 33% menos que en Caturra, confirmando las diferencias obtenidas por Romero y Cortina (2004).
En la última evaluación los intervalos de confianza del 95% para el promedio de huevos acumulados en CCC 534 y en C. liberica se traslaparon, lo que significa que aunque el período de oviposición en la especie diploide fue más prolongado que en la CCC 534, no se hallaron diferencias en el número de huevos puesto por las hembras sobre ambos materiales.
Proporción de sexos. No se encontraron diferencias en la proporción de sexos entre poblaciones de brocas criadas en Caturra, CCC 534 o C. liberica. Sus proporciones fueron 9,8, 9,2 y 9,8 hembras por cada macho, respectivamente, las cuales concuerdan con los reportes de la literatura (Bergamin 1943; Bartra et al. 1982; Montoya 1993).
Supervivencia. Las curvas de supervivencia para la broca, cuando se crió y fue mantenida sobre las tres introducciones, se ajustaron a funciones de segundo grado (p = 95%; gl. 2, 15, t = 2.13). Estas curvas fueron similares entre Caturra y las introducciones; sus intervalos de confianza para la media, en cada fecha de evaluación se traslaparon, lo que muestra que las accesiones no afectan la supervivencia de la broca. Las menores pendientes se observan al inicio por la mortalidad de los estados inmaduros, que estuvo entre el 22 y el 25%, mientras que la mortalidad de los adultos, entre los 24 y 72 días, fue menor (Fig. 4).
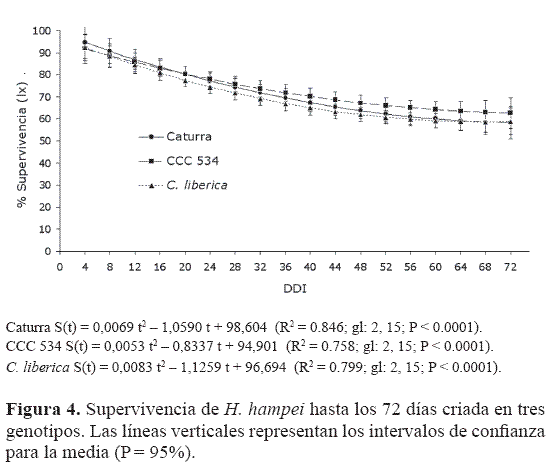
Bergamin (1943), también encontró una mortalidad baja de la broca una vez llega al grano, por la protección que este le ofrece. Este autor midió la mortalidad de 54 hembras en laboratorio, encontrando que las primeras murieron a los 81 días. Baker et al. (1992) encontraron en campo una mortalidad similar para los estados inmaduros de broca (30%). Sin embargo, esta es una estimación baja, si se considera que en el laboratorio la broca está libre de sus enemigos naturales y no sufre las adversidades del clima; también calificaron como muy baja la mortalidad de los adultos una vez penetran al fruto. Ruíz (1996), encontró supervivencias superiores del 80% para las hembras fundadoras.
Todos estos trabajos indican que el grano de café, donde ocurre la mayor parte del ciclo biológico de la broca, no solo le ofrece alimento, sino una excelente protección, por lo que las tasas de mortalidad son pequeñas. Silveira et al. (1976), dividen en tres las etapas de la vida de los insectos: pre-reproductiva, reproductiva y pos-reproductiva. La supervivencia de las dos primeras etapas corresponde a una curva parcial como las que se muestran en la figura 4. Al cambiar la escala a logarítmica (información no incluida), corresponden a la curva de supervivencia de tipo I, que presenta una baja tasa de mortalidad en la mayor parte del ciclo y muy alta al final (Deevey 1947; Slobodkin 1962).
Las etapas reproductiva y pos-reproductiva fueron halladas por Bergamin (1943), en condiciones de laboratorio similares, y, como ya se dijo, muestran baja mortalidad en la época reproductiva y alta después de los 80 días de infestación por las hembras, confirmando la pertenencia al tipo I. Esta curva de supervivencia es típica de insectos con estrategia biológica “K”. Las características de los insectos con esta estrategia que están presentes en la broca son: ser especialista, en este caso del género Coffea, y no sufrir elevada mortalidad en los estados inmaduros, debido a la protección materna. Aunque también tienen algunos elementos de la estrategia “r”, como el tamaño reducido, la madurez precoz y su alta capacidad reproductiva.
Parámetros demográficos. La tasa reproductiva neta (Ro), para la población criada sobre Caturra fue de 25 ± 1 hembras, la cual coincide con la calculada por Baker et al. (1992), y por Ruíz (1996), en campo, al infestar frutos de 210 días de edad (cuando infestó frutos de menor edad la tasa fue menor), pero superior a los 19 individuos reportados por Portilla (1999), en café pergamino. La tasa de reemplazo en CCC 534 fue menor en un 30% y en C. liberica en un 43% (Fig. 5).
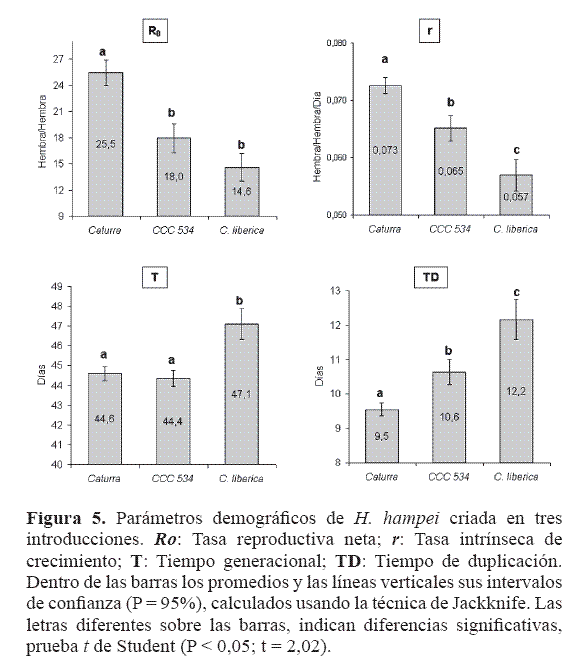
El tiempo generacional fue similar entre Caturra y CCC 534 (44,6; 44,4 días) y tres días más prolongado en C. liberica, debido a un período de oviposición más largo. Las tres introducciones tuvieron valores similares con relación a los hallados por Baker et al. (1992), Ruíz (1996), y Portilla (1999), que corresponden a 45,2, 51,4 y 41,9 días respectivamente.
De acuerdo con Rabinovich (1980), la tasa intrínseca de crecimiento es un carácter genético, que refleja la capacidad de multiplicación de la población en condiciones óptimas. Las evaluaciones de campo de Baker et al. (1992), y Ruíz (1996), arrojaron valores de r = 0,065, en variedades de C. arabica susceptibles. Portilla (1999), en granos de café en laboratorio, la estimó en 0,071, pues no se presentan condiciones ambientales extremas, ni controladores biológicos, permitiendo un mayor crecimiento de la población.
Si se considera que en Caturra se expresa el máximo potencial de crecimiento (rm= 0,073 ± 0,001), los estimados de la tasa intrínseca de crecimiento en CCC 534 (r = 0,065 ± 0,002) y C. liberica (r = 0,057 ± 0,003), muestran que estas introducciones redujeron significativamente (P < 0,05), la expresión de ese potencial (Fig. 5). La tasa intrínseca de crecimiento depende del tiempo generacional, la supervivencia y la fecundidad (Birch 1948), en la primera variable no se encontraron diferencias entre Caturra y las dos introducciones, pero en el número de huevos sí. Entre CCC 534 y C. liberica la significancia se debió a pequeñas diferencias en desarrollo y a una oviposición más tardía sobre la última. De tal manera que fue el efecto sobre la fecundidad el que determinó principalmente las diferencias en el crecimiento de las poblaciones. De igual manera y como consecuencia de lo anterior, los tiempos de duplicación fueron también diferentes en cada una de las introducciones, lógicamente la broca sobre CCC 534 o sobre C. liberica tarda más en doblar su población (Fig. 5). La menor oviposición se debe a la existencia de antimetabolitos en el grano, o a que éste carece del balance de nutrientes adecuados para una buena reproducción del insecto (Maxwell 1972; Nation 2001), lo que se podría determinar a partir del estudio del desarrollo de ovocitos. Otra causa del menor número de huevos en las introducciones y de un período más prolongado de oviposición en C. liberica, puede ser la retención de los mismos por la broca al encontrar un medio desfavorable para su descendencia, lo cual es una expresión de antixenosis (Maxwell 1972). Por otro lado, a pesar que las diferencias no son muy altas, si el mecanismo que reduce la oviposición se transfiriera a variedades comerciales, este efecto acumulado en varias generaciones reduciría notablemente las poblaciones del insecto. Esta variedad haría más eficiente y económico el manejo del insecto, al retardar el tiempo en que la población alcanza los niveles en que se hacen necesarias otras medidas de control.Conclusiones
No se hallaron diferencias en supervivencia, ni en la duración de estados biológicos de la broca, cuando fue criada sobre Caturra, la introducción CCC 534 de C. arabica o sobre C. liberica, pero si se encontró menor oviposición sobre estas introducciones, lo que incidió en una menor tasa reproductiva neta e intrínseca. Estas introducciones pueden utilizarse como fuentes de resistencia en programas de mejoramiento genético.Agradecimientos
A las disciplinas de Entomología y Mejoramiento Genético de CENICAF É, especialmente al Ingeniero Agrónomo Gabriel Alvarado A., la Dra. Pilar Moncada Botero y al señor John Esteban Quintero.Literatura Citada
AD KINSSON, P. L.; DYCK, V. A. 1984. Variedades resistentes en los sistemas de manejo de plagas, pp. 253-271. En: Maxwell, F. G.; Jennings, P. R. Manejo de plantas resistentes a insectos. Primera edición. Editorial Limusa. México. [ Links ]
ÁLVAREZ S., J. H.; CORTINA G., H. A.; VILLEGAS M., J. F. 2001. Métodos para evaluar antibiosis a Hypothenemus hampei (Ferrari), en café, bajo condiciones controladas. Cenicafé 52 (3): 205-214. [ Links ]
ANDREWARTHA, H. G.; BIRCH, L. C. 1954. The distribution and abundance of animals. University of Chicago Press. pp. 31-54. [ Links ]
ARCILA P., J.; JARAMILLO R., A.; BALDION R., J. V.; BUSTILLO P., A. E. 1993. La floración del cafeto y su relación con el control de la broca. Avances Técnicos Cenicafé (Colombia) No. 193:1-6. [ Links ]
BAKER, P. S.; BARRERA, J. F.; RIVAS, A. 1992. Life-history studies of the coffee berry borer (Hypothenemus hampei, Scolytidae) on coffee trees in Southern Mexico. Journal of Applied Ecology 29 (3): 656-662. [ Links ]
BARTRA, C.; URRELO, R.; ROD RIGUEZ, R. 1982. Biología de la broca del café Hypothenemus hampei (Ferrari) (Coleoptera: Ipidae) en Tingo María - Perú. Tropicultura (Perú) 2 (1): 17-31. [ Links ]
BERGAMIN, J. 1943. Contribuição para o cohnecimento da biología da broca do café Hypothenemus hampei (Ferrari 1867) (Col. Ipidae). Arquivos do Instituto Biológico (Brasil). 14: 31-72. [ Links ]
BIRCH, L. C. 1948. The intrinsic rate of natural increase of an insect population. Journal of Animal Ecology 17: 15-26. [ Links ]
BORBON, M. O. 1989. Bioecologie d'un Ravageur des Baies de Cafeier, Hypothenemus hampei Ferr. (Col: Scol.) et de ses parasitoides au Togo. Toulouse, Francia. L'Universite Paul Sabatier de Toulouse. 185 p. (Tesis de Doctorado). [ Links ]
BUSTILLO P., A. E. 2002. El manejo de los cafetales y su relación con el control de la broca del café en Colombia. Boletín técnico N° 24, Cenicafé. Chinchiná, Colombia. 40 p. [ Links ]
CARDONA M., C.; POSSO, C. E.; KORNEGAYS, S.; VALOR, S.; SERRANO, M. 1989. Antibiosis effects of wild dry bean accessions on the mexican bean weevil and the bean weevil (Coleoptera: Bruchidae). Journal Economic Entomology 82 (1): 310-315. [ Links ]
DEEVEY, E. 1947. Life tables for natural populations of animals. Quaterly Review of Biology 22: 283-314. [ Links ]
DUARTE N., M. T. 1992. Determinación de la atractividad de frutos de varios cultivares de café a la broca del fruto del cafeto Hypothenemus hampei (Ferrari), utilizando el método de olfatometría a nivel de laboratorio. Guatemala, Universidad de San Carlos, Facultad de Agronomía, 55 p. (Tesis: Ingeniero Agrónomo). [ Links ]
DUQUE, O. H.; CHAVES, C. B.; HERNÁNDEZ, S. M. 1997. Costos del manejo de la broca Hypothenemus hampei (Ferrari) en lotes comerciales de café del departamento del Risaralda. Federación Nacional de Cafeteros de Colombia. Centro Nacional de Investigaciones de Café. Cenicafé. Chinchiná, Colombia. 37 p. [ Links ]
FEDERACIÓN NACIONAL DE CAF ETEROS DE COLOMBIA (Fedecafé). 1997. Sistema de información cafetera, SICA. Encuesta nacional cafetera. Estadística cafetera, informe final. Federación Nacional de Cafeteros. Bogotá, Colombia. 164 p. [ Links ]
FEDERACIÓN NACIONAL DE CAF ETEROS DE COLOMBIA (Fedecafé). 1999. Informe anual de la gerencia técnica 1997-1998. Gerencia técnica. Bogotá, Colombia. 106 p. [ Links ]
FERNIE, L. M.; LANGLEY, C. I. 1986. Arabica coffee storage. Part II. A review of the problem in Tangayika. Kenya Coffea 31 (367): 297-299. [ Links ]
HARCOURT, D. G. 1967. The development and use of life tables in the study of natural insect populations. Annual Review of Entomology, 12: 175-196. [ Links ]
KOCH, V. J. M. 1973. Abondance de Hypothenemus hampei Ferr., scolyte des graines de café, en fonction de sa plant-hote et de son parasite Cephalonomia stephanoderis Betren, en Cote d'Ivoire. Wageningen, Holanda, Veenman and Zonen, 84 p. (Tesis de Doctorado). [ Links ]
MAIA, A. de H.; LUIZ, A. J.; CAMPANHOLA, C. 2000. Statistical inference on associated fertility life table parameters using jackknife technique: computational aspects. Journal Economic Entomology 93 (2): 511-518. [ Links ]
MAXWELL, F. G. 1972. Host plant resistance to insectsnutritional and pest management relationships, pp. 599-609. En: Rodríguez, J. G. Insect and mite nutrition. Significance and implications in ecology and pest management. North- Holland publishing company. Netherlands. [ Links ]
MEYER, F. G. 1968. FAO Coffee mission to Ethiopia 1964- 1965. Roma, Italia, FAO . 200 p. [ Links ]
MINISTERIO DE AGRICULTURA Y DESARROLLO RURAL (MAD R). La Cadena Del Café En Colombia. 2006. www.agrocadenas.gov.co Fecha última revisión: enero 2006. Fecha último acceso: [octubre 2006]. [ Links ]
MONTOYA O., S. A. 1993. Ciclo de vida de la broca del café (Hypothenemus hampei (Ferrari)) sobre frutos en diferentes estados de desarrollo. Bogotá, Colombia, Universidad Nacional de Colombia, Facultad de Ciencias Agropecuarias, 77 p. (Tesis Ingeniero Agrónomo). [ Links ]
NATION, J. L. 2001. Insect physiology and biochemistry. CRC Press. Boca Raton. 485 p. [ Links ]
PORTILLA R., M. 1999. Mass production of Cephalonomia stephanoderis on Hypothenemus hampei reared using artificial diet. Ascot Berkshire, Inglaterra, University of London, 253 p. (Tesis de Doctorado). [ Links ]
PRICE, P. W. 1997. Insect Ecology. 3. ed. John Wiley & Sons, New York, 874 p. [ Links ]
RABINOVICH, J. E. 1980. Introducción a la ecología de poblaciones animales. México. Ed. Continental. 313 p. [ Links ]
ROMERO, J. V.; CORTINA G., H. 2004. Fecundidad y Ciclo de Vida de la broca Hypothenemus hampei F. (Coleoptera: Curculionidae: Scolytinae) en introducciones silvestres de café. Cenicafé 55 (3): 221-231. [ Links ]
RUÍZ C., R. 1996. Efecto de la fenología del fruto de café sobre los parámetros de la tabla de vida de la Broca del Café Hypothenemus hampei (Ferrari). Universidad de Caldas. Facultad de Ciencias Agropecuarias Tesis: Ingeniero Agrónomo, 87. p. 62. [ Links ]
SAS. 1999. SAS/STAT User's Guide. 8.0 version. SAS Institute. Inc., Cary, NC, EUA. [ Links ]
SILVA, G. 2006. Cuatro Años Cosechando Futuro: Resultados de Política Gremial Cafetera 2002-2006. www.cafedecolombia.com/documents/cartilla_version_web.pdf "Fecha última revisión: agosto 2006. Fecha último acceso: [octubre 2006]. [ Links ]
SILVEIRA N., S.; NAKANO, O.; BARBIN, D.; NOVA, N. A.V. 1976. Manual de ecología dos insetos. São Paulo (Brasil), Editora Agronomica Ceres, 419 p. [ Links ]
SLOBOD KIN, L. B. 1962. Growth and regulation of animal populations. Holt, R & W, New York. [ Links ]
SOKAL, R. R.; ROHLF, F. J. 1995. Biometry. Third edition. New York: W. H. Freeman & Co. 887 p. [ Links ]
SOROKA, J. J.; MACKAY, P. A. 1991. Antibiosis and antixenosis to pea aphid (Homoptera: Aphididae) in cultivars of field peas. Journal of Economic Entomology 4 (6): 1951- 1956. [ Links ]
TRICHILO, P. J.; LEIGH, T. F. 1985. The use of life tables to assess varietal resistance of cotton to spider mites. Entomologia Experimentalis et Applicata 39: 27-33. [ Links ]
VILLAGRAN G., W. 1991. Atractividad relativa y susceptibilidad de varias especies y cultivares de café Coffea spp. a la broca del fruto Hypothenemus hampei Ferr. en condiciones de laboratorio. Guatemala, Universidad de San Carlos de Guatemala. Facultad de Agronomía, 43 p. (Tesis: Ingeniero Agrónomo). [ Links ]
Recibido: 17-jun-2005- Aceptado: 034-feb-2007

