










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488On-line version ISSN 2665-4385
Rev. Colomb. Entomol. vol.33 no.1 Bogotá Jan./June 2007
Sección Morfología, Comportamiento, Ecología, Evolución y Sistemática
Ciclo de vida y potencial reproductivo de la hormiga depredadora Solenopsis cf. picea
(Hymenoptera: Formicidae)
Life cycle and reproductive potential of the predator ant Solenopsis cf. picea (Hymenoptera: Formicidae)
JANINE HERRERA RANGEL1 e INGE ARMBRECHT1-2
1 Autor para correspondencia: Universidad del Valle, Departamento de Biología, Ciudad Universitaria Meléndez calle 13 No. 100-00, tel: 321 21 00 ext. 2570. A. A. 25360, Cali, Colombia. jahera@gmail.com
2 Universidad del Valle, Departamento de Biología. inge@univalle.edu.co
Resumen. Solenopsis cf. picea, también conocida como “la negrita”, es una hormiga abundante en los sistemas cafeteros de la zona de Risaralda (Colombia), donde anida en el suelo y se encuentra tanto en cafetales de sombra como monogenéricos. Estudios realizados en campo y en laboratorio han demostrado que depreda sobre la broca del café, convirtiéndola en un organismo que podría ser implementado como control biológico. Este trabajo tuvo como objetivo estudiar algunos aspectos de su ciclo de vida, en términos de desarrollo del inmaduro y aspectos reproductivos del organismo. Se encontró que el desarrollo (reconocimiento cronológico y morfológico) de inmaduros correspondió a un promedio de 56 días. Las medidas de cápsula cefálica agruparon a los individuos en tres ínstares larvales. El promedio de la postura de huevos durante 24 horas fue significativamente mayor en colonias monoginas acompañadas de densidades altas de obreras y la fecundidad individual de las reinas disminuyó con la presencia de otras reinas en una misma colonia.
Palabras claveCápsula cefálica. Monogina. Oviposición. Poligina.
Abstract. Solenopsis cf. picea, also known as “the little black”, is an abundant ant in the coffee systems of Risaralda, Colombia, where it nests in the soil and can be found in both shaded coffee plantations and in coffee monocultures. Studies carried out in the field and laboratory have demonstrated that it predates on the coffee berry borer, making it an organism that could be implemented as a biological control. The objective of this research was to study some aspects of its life cycle, in terms of development of the immature and reproductive aspects of the organism. Results showed that immature development (chronological and morphological recognition) took an average of 56 days. The cephalic capsule measures grouped individuals into three larval instars. Mean number of eggs laid in 24 hours was significantly higher in monogynous colonies accompanied by high densities of workers, and the individual fecundity of the queens decreased with the presence of other queens in the same colony.
Key words. Head capsule. Monogynous. Oviposition. Polygynous.
Introducción
Los sistemas cafeteros en Colombia tienen gran importancia no solo económica, sino también social y ecológica. Alrededor de ellos se teje una cultura que se ha mantenido a través de generaciones presentando poblaciones que basan su economía en estos cultivos. El interés ecológico se relaciona con el sistema de manejo, pues los cafetales de sombra pueden ayudar a conservar fauna silvestre (Moguel y Toledo 1999).
En una caracterización de hábitat para fincas cafeteras de la zona de Risaralda, bajo diferentes sistemas de manejo, Armbrecht (2003a) encontró similitudes entre bosque y plantaciones poligenéricas de café en sombra. Basados en una recopilación de datos, Swift et al. (1996) concluyeron que la biodiversidad es frecuentemente más alta en agroecosistemas tradicionales, e incluso significativamente alta en un sistema de intensificación intermedio, pero la transformación a un manejo altamente intensificado o “moderno” resulta en una pérdida sustancial de biodiversidad asociada.
Dependiendo de la situación particular y el lugar involucrado, el policultivo puede promover (mejorando) la utilización de nutrientes, control de plagas, control de malezas y otras funciones agrícolas (Swift et al. 1996). El alto número de especies de plantas y animales hospederas en cultivos de café tradicional bajo sombra indican que este sistema puede jugar un papel de conservación como hábitat antropogénico protector para especies originarias de bosque (Moguel y Toledo 1999). Dado el valor de este agroecosistema, es importante que se preserve y sea utilizado como medio complementario de conservación.
Los himenópteros especialmente de la familia Formicidae se han utilizado en estudios ecológicos y de impacto ambiental, no solo por su abundancia sino por sus funciones en la naturaleza pues son depredadores y simbiontes (Agosti et al. 2000). La variedad de dietas, de sitios para anidar, la duración de sus ciclos de vida y asociaciones con casi cualquier hábitat dado hace de éstas un grupo atractivo para monitorear (Kaspari 2000). Vélez et al. (2001), Armbrecht (2003b) y Gallego (2004) mostraron evidencia en campo y laboratorio que las hormigas de suelo juegan un papel en la depredación de adultos de broca en plantaciones de café de Colombia. En México, Armbrecht y Perfecto (2003) encontraron hormigas pequeñas, como Solenopsis spp., que podían penetrar los hoyos hechos por la broca en la cereza del café.
Solenopsis, al contrario de algunos otros agentes de control biológico usados en agroecosistemas, no es especialista sino generalista (Risch y Carroll 1982). Varios estudios han sugerido que este género puede ser un importante controlador biológico de insectos plagas (Risch y Carroll 1982; Way y Khoo 1992; Varón 2002; Vélez et al. 2003; Gallego y Armbrecht 2005; Armbrecht y Gallego en prensa). Solenopsis geminata mostró ser un predador importante en los agroecosistemas de maíz-calabaza en el sur de México (Risch y Carroll 1982). Solenopsis spp., particularmente la introducida, en el sur de Estados Unidos S. invicta tiene un cierto atributo como controlador biológico, de manera tal que ha sido fomentada en localidades donde el daño es pequeño o ninguno (Way y Khoo 1992).
Solenopsis cf. picea (en adelante S. picea ) (Hymenoptera: Formicidae: Myrmicinae) es una hormiga abundante en los sistemas cafeteros de la zona de Risaralda (Gallego 2004; Armbrecht et al. 2005; Gallego y Armbrecht en prensa). Se ha considerado como hormiga dominante de suelo, en sistemas de sombra, estructurada en modelo de distribución discreta, que pierde complejidad en plantaciones de café de sol (Armbrecht et al. 2005). Rivera (2003) ubicó a esta especie dentro del gremio de hormigas abundantes en hojarasca en un estudio de cafetales en la misma zona de Risaralda, y es una de las especies de hormigas a la cual se le ha comprobado su función depredadora sobre la broca (Gallego y Armbrecht 2005; Armbrecht y Gallego en prensa).
En anteriores estudios se encontró mayor actividad predadora en la estación lluviosa y S. picea resultó ser un significativo componente de la diversidad de hormigas que depredaron la broca. La depredación puede ser un importante mecanismo regulador en granjas cafeteras de sombra en la estación húmeda, siendo esta estación importante para cafeteros en Colombia porque es el tiempo de cosecha, cuando mayor fuente de infestación es generada y quedan residuos de cosecha en el suelo donde confluyen la broca y la hormiga (que anida en el suelo) (Armbrecht 2003b). Teniendo en cuenta el beneficio que puede aportar S. picea a diversos sectores de la agricultura, en especial los relacionados con agroecosistemas tradicionales de café, el objetivo del presente estudio fue obtener información biológica básica del desarrollo y reproducción de la hormiga, que no se conoce, y examinar algunos factores que podrían potenciar su cría artificial (p. ej.: número de reinas y número de obreras).
Materiales y Métodos
Obtención de colonias. Las colonias de hormigas se obtuvieron en el municipio de Apía, departamento de Risaralda (5º 08’ N; 75º 56’ W), ubicado entre un rango altitudinal de 1400-1900 msnm. La región tiene temperaturas entre los 18-20ºC, humedad relativa del 80% y un promedio de precipitación anual de 2.320 mm, con distribución bimodal con picos en mayo y noviembre (Gallego 2004; Armbrecht et al. 2005).
Apía es un pueblo rodeado por fincas cafeteras bajo diferentes tipos de manejo, hasta hace poco tiempo predominaron los sistemas tradicionales bajo sombra. Con el tiempo, la tecnificación aumentó en los cultivos, hasta encontrar hoy en día sistemas de monocultivo completamente bajo sol intercalados con cultivos bajo sombra (Armbrecht et al. 2005).
Las colonias utilizadas fueron obtenidas de dos fincas, “La María” con sistema de monocultivo de café bajo sol a 1.405 msnm, y “La Playita” con sistema tradicional de café orgánico bajo sombra a 1.490 msnm. Dentro del cultivo de café se ubicaron doce eras. Una “era” se considera como el espacio comprendido entre dos franjas de arbustos de café, por lo que asemeja una línea recta o transecto. En cada era se ubicaron cinco puntos o estaciones con cebos de atún, cada uno de ellos separados por 3 m, hasta que el cultivo lo permitiera o hasta llegar a un tope de 10 cebos. Después de 15 o 20 minutos se revisaba cada estación dentro de las eras, en donde el cebo había atraído a S. picea y se realizaba un seguimiento de la pista conformada por las hormigas forrajeando hasta el cebo. En el punto donde el forrajeo, o la pista, era interrumpido se cavaba en el suelo con una pala de mano hasta que se encontraran cámaras con cría del hormiguero. Éstas se extraían con sumo cuidado y se introducían en una bolsa con cierre hermético.
Cada colonia se guardó por separado y con cierta frecuencia se abría la bolsa para suministrar aire, hasta que fueran establecidas en el laboratorio.
Establecimiento de la colonia. Las bolsas fueron trasladadas al Laboratorio de Hormigas en la Universidad del Valle (Cali). El método utilizado fue similar al implementado por Gallego (2004), en el cual las colonias se mantuvieron artificialmente en una cámara de cría a una temperatura entre 22-25ºC y humedad relativa de 65%.
A cada colonia se le retiró el exceso de tierra, estableciéndose cada una en cajas plásticas de 24 x 24 x 10 cm. Los bordes internos fueron impregnados con Fluón® de Bioquip (película deslizante que se aplica húmeda) mediante un pincel o algodón para evitar escape de hormigas. Cada caja contenía un nido artificial en su interior para mantener la humedad. El nido artificial fue elaborado de pasta-yeso y protegido de la luz con un recipiente oscuro para que las hormigas se adaptaran con el tiempo a las condiciones artificiales del laboratorio. La caja estaba cubierta por una tapa plástica con dos agujeros cubiertos con malla metálica muy fina para permitir el intercambio de gases.
La alimentación de las colonias se basó en una dieta descrita por Keller et al. (1989) para criar varias especies de hormigas. Adicionalmente a ésta se les brindó miel y agua en un vial tapado con un poco de algodón que les sirvió como dosificador. Cada semana se renovó el alimento.
Ciclo de vida. Para cumplir este objetivo se determinó primero la duración de los diferentes estados de desarrollo (huevo, larva, pupa) de la hormiga. Para hacer el seguimiento se separó la reina y algunas obreras de una colonia inicial a una subcolonia. Al cabo de un par de días la reina era devuelta a la colonia de origen (inicial), quedando la subcolonia compuesta de obreras al cuidado de la cría. A esta cría, consistente inicialmente de huevos, se le observó su desarrollo hasta la etapa adulta. Se utilizaron cuatro subcolonias.
Luego del reconocimiento cronológico, se procedió a distinguir cada etapa larval mediante la medición y observación de la cápsula cefálica, mandíbulas y longitud de la larva con la ayuda de un microscopio estereoscopio Nikon SMZ 800 (1X) y SMZ 645 (2X), al cual se adaptó un micrómetro (Arcila et al. 2002b). Dado el pequeño tamaño, las larvas y huevos se pueden deshidratar en unos cuantos segundos, por lo cual, durante las mediciones, estos estados se mantuvieron embebidos en solución fisiológica (isotónica) para evitar su muerte; después de tomadas las medidas, estos eran devueltos a la colonia.
La mayoría de las larvas provenían de las colonias más antiguas, establecidas con anterioridad al estudio (aproximadamente dos años), y en menor cantidad de las colonias más recientes (aproximadamente dos meses) que no tenían reinas. Los datos fueron tomados de febrero a abril y de junio a agosto del 2005.
Reproducción
Oviposición de colonias monoginas frente a densidades de obreras. Para estimar la productividad en términos de número de huevos puestos por día, se realizaron pruebas de oviposición como las utilizadas por Arcila et al. (2002a) y Ulloa-Chacón (2003). Las pruebas consistieron en aislar cada reina en compañía de siete u ocho obreras durante 24 horas; al cabo de este tiempo se contaron los huevos puestos.
Se sometieron a prueba seis colonias monoginas (p. ej.: con una sola reina) con alta densidad (300 individuos) de obreras y seis colonias monoginas con baja densidad (20 individuos), para un total de 12 colonias. Cada una de ellas se estableció con un mes de anterioridad antes de ser sometidas a prueba. Se realizaron pruebas de oviposición una vez por mes durante cuatro meses.
Oviposición de colonias monoginas frente a colonias poliginas. Para cumplir con este objetivo, se establecieron colonias con una, dos y cuatro reinas con densidades altas (300 individuos) de obreras. Se emplearon en total nueve colonias y 16 reinas, las cuales fueron sometidas a pruebas de oviposición una vez por mes durante tres meses. De éstas, cuatro fueron monoginas, cuatro poliginas con dos reinas y una poligina con cuatro reinas.
Análisis de datos. Los datos de pruebas de oviposición se analizaron mediante la prueba de homogeneidad de varianzas de Levene y posteriormente una prueba t-student para muestras independientes. Cuando fue requerido, se utilizaron pruebas no paramétricas de Mann-Whitney y Kruskall Wallis. Estos análisis fueron realizados en el programa estadístico PAST versión 0.98c (Hammer et al. 2001). Las gráficas y parte de la estadística descriptiva se realizaron en Excel (2003).
Resultados
Ciclo de vida
Reconocimiento cronológico. El tiempo de desarrollo de S. picea desde huevo hasta estado adulto fue de 56 días promedio bajo condiciones de laboratorio (22-25ºC y H. R. 65%). La etapa de huevo se prolongó en promedio 13 días, con un rango de 11 a 15. El estado de larva, incluyendo sus tres estadíos o ínstares larvales, tuvo una duración promedio de 31 días. Al completar este estado el individuo se encuentra entre el día 43 y 45 de edad total, siendo éste el de mayor duración. La pupa tuvo un tiempo promedio de 12 días, finalizando su desarrollo entre el día 55 y 57.
No se obtuvo información sistemática de la longevidad de adultos, pero se puede asegurar que al menos una colonia colectada durante el estudio de Gallego y Armbrecht (2005) sobrevivió por dos años y ocho meses en cautiverio. Esto significa que la reina mantiene su fecundidad al menos dos años.
Se esperaría que en campo la duración del ciclo de vida difiera un poco de los resultados obtenidos en laboratorio, los cuales son una base preliminar pero confiable. De ahí se sugeriría reproducir las colonias para liberarlas en campo de unas tres a seis veces por año, y liberarlas al suelo, en focos de infestación de cafetales, tanto de sol como de sombra, inmediatamente después de la cosecha cafetera. Por observaciones realizadas en campo, en territorios de S. picea en fincas cafeteras de Apía (Armbrecht 2003b, capítulo 3) se sospecha que la influencia pueda permanecer en territorios estables por meses.
Etapas larvales. De un total de 88 individuos, a 75 se les midió la amplitud de las mandíbulas. No fue posible medir los 13 restantes debido a que las larvas recién eclosionadas no presentaban esclerotización en las mandíbulas.
La longitud de la larva fue la medida de mayor variación (± 0,33), seguida por el diámetro de la cápsula cefálica (± 0,03) y la de menor variación fue la amplitud mandibular (± 0,01). Para la primera, se encontraron datos desde 0,1 mm hasta 1,44 mm de manera continua, con un promedio de 0,89 mm; mientras que los obtenidos para la segunda medida se encontraron en un rango más estrecho, de 0,10 mm hasta 0,34 mm. Finalmente, en la amplitud mandibular se encontraron datos desde 0,05 mm hasta 0,10 mm. La frecuencia de aparición de individuos para las medidas con menor variación de datos (amplitud mandibular y diámetro de la capsula cefálica), mostraron tendencias similares. Aún así, se consideró el diámetro de la cápsula cefálica debido a que abarca el número total de individuos (n = 88). De esta manera, se agruparon tres intervalos de medidas donde se infiere que hay tres grupos o ínstares (Fig. 1).
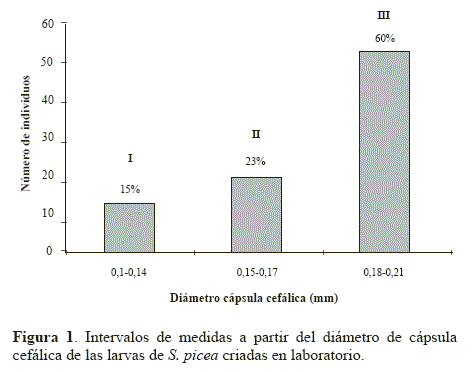
No fue posible detectar la muda de las larvas. Posiblemente las larvas recuperan nutrientes ingiriendo (ayudadas por las obreras nodrizas) su propia muda.
Reproducción
Oviposición en colonias monoginas. El promedio de huevos puestos por reina durante las pruebas fue consistentemente mayor en alta densidad de obreras. La primera prueba de oviposición presentó los valores más altos a lo largo del experimento, con un promedio de nueve huevos por reina en condiciones de alta densidad de obreras y un promedio de 3,5 huevos por reina en condiciones de baja densidad de obreras. La postura de huevos se vio negativamente afectada en ambas densidades disminuyendo su número con el tiempo, los resultados más bajos se obtuvieron en la tercera prueba, en donde el promedio de huevos en alta densidad fue de 2,4 y de un huevo para baja densidad por cada reina presente. Para la cuarta prueba se observaron signos de recuperación en ambos casos.
El promedio de huevos puestos por día y obtenidos para cada reina en su colonia también demostró ser mayor en aquellas reinas atendidas por un mayor número de obreras (alta densidad). El promedio de huevos puestos por reina acompañadas por altas densidades de obreras fue de 5,1 huevos en 24 horas con un máximo de 35 huevos por día y el promedio de huevos por reina asociadas a bajas densidades de obreras fue de dos huevos en 24 horas con un máximo de 14 huevos por día. Se presentaron diferencias significativas (U = 94, N1: 22, 2 reina bajo diferentes densidades (Fig. 2).
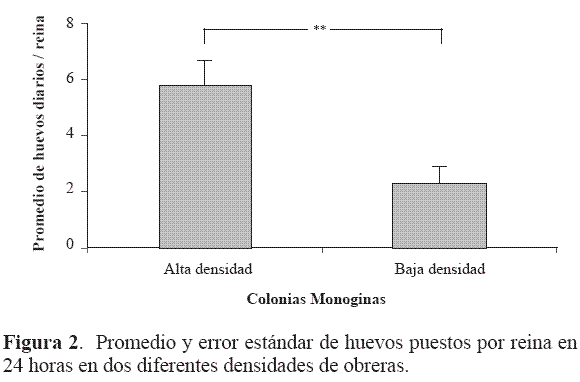
Oviposición en colonias poliginas. Teniendo en cuenta la productividad de las reinas como el número de huevos puestos en 24 horas, se estableció que las reinas de colonias monoginas muestran una tendencia a ser más productivas que las reinas que se encuentran acompañadas por otras reinas en una misma colonia. Dentro de las colonias poliginas se observó ligeramente que dos reinas son más ponedoras que cuatro reinas acompañadas. El promedio de huevos por día en las reinas solitarias fue de 6,5, en colonias poliginas con dos reinas el promedio fue de 4,5 huevos y con cuatro reinas 2,4 huevos puestos (Fig. 3).

No se encontraron diferencias entre los distintos tipos de colonia (Kruskal-Wallis, H: 5.106, H : 5,274, P = 0,07) aunque la no significancia fue marginal (P = 0,05). Se realizaron pruebas que relacionaran colonias monoginas con cada una de las poliginas, y se encontró que entre las colonias poliginas (dos y cuatro reinas) no existen diferencias significativas (t: 1,15, gl: 27, P = 0,25). De igual forma, no existen diferencias en la postura entre reinas únicas y dos reinas (t: 0,83, gl: 24, P= 0,41). Lo contrario ocurrió con reinas únicas y poliginas de cuatro reinas, las cuales presentaron diferencias significativas en la oviposición por día (U: 23,5, N : 9 N : 12, P = 0,03). Aún así, se puede apreciar claramente una relación negativa (R2: 0,9) entre el promedio de huevos puestos en 24 horas y el número de reinas presentes en una colonia (Fig. 3).
Discusión Ciclo de vida y etapas larvales. Al igual que S. picea , otros ejemplos de hormigas de interés son Paratrechina fulva Mayr, 1862 que bajo condiciones de laboratorio (27ºC, 80% H. R.) en su desarrollo de huevo a adulto, comprende de 23 a 50 días, considerado un tiempo corto comparado con otros miembros de Formicinae (Arcila et al. 2002b). El tiempo límite en días para larvas de obreras en la subfamilia Myrmicinae es: huevo 6-29, larva 5-44, pupa 8-28 (Wheeler y Wheeler 1976). Para la “hormiga roja de fuego”, Solenopsis invicta, dependiendo de la temperatura, pueden tomar de 20 a 45 días (Oklahoma Department of Agriculture 2005). En Solenopsis molesta el tiempo mínimo que puede alcanzar los estados de huevo, larva, prepupa y pupa son 16, 21, 2 y 13 días respectivamente, para una duración total mínima de desarrollo de 52 días (Hayes 1920). De acuerdo con lo anterior, La duración de las etapas del desarrollo inmaduro de S. picea , obtenidas en condiciones de laboratorio, se encuentra dentro de los rangos observados para otras hormigas, en especial de la misma subfamilia (Myrmicinae). Además, se debe tener en cuenta que el medio artificial en que fueron mantenidas, aunque cumple requerimientos nutricionales, probablemente no replíca adecuadamente las mismas condiciones de campo. Según Wheeler y Wheeler (1976) la influencia de la temperatura, suplemento alimenticio, estación y quizás otras variables son responsables de parte de la variabilidad en las mismas especies e incluso en las mismas colonias. El número de ínstares larvales no se ha reportado de manera generalizada para hormigas. Se conoce que para algunas especies estudiadas, ocurren desde tres, cuatro y en raras ocasiones cinco instares (Wheeler y Wheeler 1976). En P. fulva midiendo la variable del ancho de la cápsula cefálica mostró cuatro valores, sugiriendo cuatro ínstares larvales, tres corresponden a producción de obreras y el cuarto para machos (Arcila et al. 2002b). En la información recopilada para la hormiga roja de fuego se presentan cuatro ínstares larvales (Oklahoma Department of Agriculture 2005). Reproducción. Para una máxima eficiencia en la colonia, la división de labor requiere acoplamiento entre la proporción de huevos puestos por las reinas y la disponibilidad de obreras para cuidar la cría (Tschinkel 1988). Para S. picea , las reinas son más productivas en presencia de altas densidades de obreras que con pocas obreras, siendo muy posible que la reina reciba mayor asistencia por parte de mayor número de obreras. La hipótesis dominante, nombrada por Tschinkel (1988) concerniente al control de la fecundidad de la reina, es que la larva tiene la prioridad sobre las reinas en la competencia por la comida y atención por parte de las obreras. Esto hace que la fecundidad de la reina esté positivamente relacionada con el número de obreras y negativamente con el número de larvas. Tschinkel (1988), al estudiar la fecundidad en colonias monoginas en S. invicta, encontró, a diferencia de otros trabajos, que las obreras por sí solas son incapaces de estimular la oviposición de las reinas y demostró que la proporción de huevos puestos en reinas está dependiendo fuertemente del número de larvas. Por lo tanto, la fecundidad de la reina en S. invicta está regulada a través de una alta especialización dependiente de la larva. Cabe mencionar que en el momento de establecer las colonias que se sometieron a prueba, se establecían más rápido cuando se adicionaban larvas, por eso todas las colonias se establecieron con muy pocas larvas (seis). En este estudio, no se evaluó la presencia de larvas, pero sí la de las obreras, las cuales influencian la postura de huevos en la reina de manera positiva, ya que mayor número de obreras proporciona más alimento para la reina y más cuidado. El control de la fecundidad de las reinas individuales se puede deber a numerosas variables, entre ellas encontramos factores externos, como la alimentación, temperatura, humedad y fotoperiodo, factores internos relacionados con la edad de los individuos (peso) y factores sociales como el número de reinas por colonia (Arcila et al. 2002a). En lo que respecta a este último factor, la fecundidad de la reina es inversamente proporcional a su número en la colonia. Ejemplo de ello lo constituye Pachycondyla villosa (Trunzer et al. 1998), donde la postura de huevos fue significativamente diferente entre una sola reina y grupos de tres reinas, al igual que entre colonias de dos reinas y de tres reinas. Paratrechina fulva (Arcila et al. 2002a), Wasmannia auropunctata (Ulloa-Chacón 2003), Solenopsis invicta (Tschinkel 1988) e Iridomyrmex humilis (Keller 1988, esta especie en la actualidad es Linepithema humile) entre otras. La disminución en la fecundidad individual de las reinas poliginas puede ser consecuencia de una cadena de hechos que finalmente afectan a la reina. En colonias poliginas de I. humilis, no existió evidencia de una dominancia en jerarquía entre reinas. En cambio, la diferencia en fecundidad de reinas, en poliginas así como en control de colonias monoginas, resultaba de las diferencias fisiológicas intrínsecas entre las reinas. Las reinas en colonias poliginas son menos atractivas a las obreras, de esta manera tienen menos contacto con obreras y en consecuencia, probablemente reciben menos comida que aquellas monoginas (Keller 1988; Keller y Passera 1989). Una feromona de inhibición entre reinas en la misma colonia reduce la producción de huevos o reduce la atracción de obreras a las reinas bajo condiciones poliginas, las cuales causan decrecimiento en la alimentación y por ende a la reducción en la tasa de oviposición en P. fulva (Arcila et al. 2002a). En W. auropunctata la menor oviposición por reina se produce como consecuencia de una menor coordinación y eficiencia entre obreras de sociedades poliginas (Ulloa-Chacón 2003). Es posible considerar que una reina pueda resultar más atractiva que otras y por ende recibir mayor atención por parte de las obreras. De esta manera, reinas en colonias poliginas estarían compitiendo, no solo por una jerarquía reproductiva, sino también por alimento y otros cuidados. Muy similar ocurre en sociedades monoginas donde la competencia por la atención de la obrera es entre reinas y larvas. Una o varias reinas que se encuentren menos atendidas por las obreras recibirán menos alimento y cuidado que reinas más atractivas. Por consiguiente el número de huevos puestos será menor. Observaciones que puedan apoyar este hecho se encontraron en una colonia poligina de dos reinas en particular, en la cual la postura de una de ellas (25 huevos) fue muy por encima de su compañera (un huevo). Estos datos contribuyeron de manera contundente a la variación; y son consistentes con la explicación propuesta. Durante el presente estudio, cuando se intentó constituir más colonias de cuatro o dos reinas, se observaron dos situaciones particulares: por un lado una o varias reinas sin atención, desplazadas del nido artificial de yeso hasta llegar a morir, y por el otro agresividad hacia la reina por parte de las obreras hasta encontrar una de las reinas descuartizada. En la literatura se reporta que en la mayoría de casos estudiados con varias especies de hormigas, la fundación de la colonia con múltiples reinas se reducen a una sola reina ponedora, por lo menos dentro las áreas locales del nido, poco después de la primera eclosión de las obreras (Hölldobler y Wilson 1990). Estas obreras eliminan el exceso de reinas, o las reinas se agreden y eliminan entre sí. Cuando la colonia tolera otras reinas inseminadas cerca de la asociación con la primera ponedora, el desarrollo de sus ovarios son usualmente suprimidos (Hölldobler y Wilson 1990). Teniendo en cuenta la fecundidad de las reinas de S. picea , basado en lo observado, el tipo de colonia en que la reina es más productiva es en sociedades con altas densidades de obreras con una única reina, incluso hasta grupos de dos reinas. Esto significa que, aunque en campo se encuentran colonias con entre dos y cinco reinas en condiciones naturales, es muy posible que estas colonias puedan sufrir una reducción del número de sus reinas para épocas que deben ser más productivas en términos de número de huevos. El intervalo de postura en general, bajo condiciones de laboratorio, de una reina era de 0-35 huevos por día en monoginas y de 0-25 en poliginas. Esto significa que existe un potencial reproductivo muy alto para la especie. En W. auropunctata el intervalo varía de 16 a 41 huevos reina por día en monogina y en poligina entre cinco a 25 huevos reina por día (Ulloa-Chacón 2003). La diferencia se logra apreciar en el promedio. Mientras en S. picea en colonias monoginas el promedio fue de 6,5 huevos reina por día, en monoginas de W. auropunctata fue de 27,4 huevos reina por día y en poliginas de 12,3 huevos reina por día (Ulloa-Chacón 2003). En P. fulva el promedio en monoginas fue de 21 huevos y en hexaginas (seis reinas por colonia) fue de 8,2 huevos reina por día. De todo lo anterior se puede deducir que S. picea se comporta reproductivamente en forma similar a otras especies de hormigas tropicales estudiadas en condiciones homologables. Aunque la especie es un poco más susceptible a la manipulación, es de fácil recuperación una vez se deja de manipular, y así lo evidencian dos colonias en laboratorio establecidas con anterioridad (dos años) a este estudio. S. picea no presenta una alta poliginia y se encuentra naturalmente con más frecuencia en colonias monoginas. Por lo observado en este estudio y combinando los datos recopilados en campo y laboratorio, es probable que se presente en esta especie, una fundación de la sociedad acompañada con múltiples reinas (pleometrosis) pero sobreviviendo al final una reina (monogina secundaria). Con miras al control biológico, ya se conoce el tipo de colonias que son más productivas en términos de número de obreras. Si se asume que el consumo de broca es directamente proporcional a la cantidad de obreras que tenga la colonia, va a ser más ventajoso mantener en el cafetal sociedades monoginas (o diginas) con altas densidades de obreras. El ciclo de vida no es muy largo y se podría esperar que estas sociedades alcancen grandes tamaños en periodos relativamente cortos. Faltaría conocer las condiciones que estimulan la aparición de reproductores de S. picea , ya que en el tiempo de estudio no se observó el nacimiento de ningún reproductor en cautiverio.
Agradecimientos Los autores agradecen a Patricia Chacón, Angela Arcila y Leonardo Rivera del grupo de investigación en biología, ecología y manejo de hormigas de la Universidad del Valle, por compartir su experiencia y conocimiento, a Carlos Cultid, Patricia Cadena, Paola Gonzáles y Jennifer Girón por la colaboración en campo. A José Pacheco de la Universidad de Texas, por identificar S. cf. picea, Miguel Márquez y María Cristina Gallego que establecieron las primeras colonias dos años antes, a la Universidad del Valle y a la Sociedad Colombiana de Entomología por la financiación de este proyecto mediante las becas a estudiantes de pregrado.
Literatura Citada AGOSTI, D.; MAJER, J. D.; ALONSO, L. E.; SHULTS, T. R. (eds.). 2000. Ants: Standard methods for measuring & monitoring biodiversity. Smithsonian Institution. Washington, D. C. 280 p. [ Links ] ARCILA, A. M.; ULLOA-CHACÓN, P.; GÓMEZ, L. A. 2002a. Factors that influence individual fecundity of queens and queen production in crazy ant Paratrechina fulva (Hymenoptera: Formicidae). Sociobiology 39 (2): 1-12. [ Links ] ARCILA, A. M.; GÓMEZ, L. A.; ULLOA-CHACÓN, P. 2002b. Immature development and colony growth of crazy ant Paratrechina fulva under laboratory conditions (Hymenoptera: Formicidae). Sociobiology 39 (2): 307-321. [ Links ] ARMBRECHT, I. 2003a. Habitat changes in Colombian coffee farms under increasing management intensification. Endangered Species Update 20 (4-5): 129-208. [ Links ] ARMBRECHT, I. 2003b. Diversity and function of leaf litter ants in Colombian coffee agroecosystems. Tesis de Doctorado. Ann Arbor, Michigan. Universidad de Michigan. Escuela de Recursos Naturales y Ambiente. 242 p. [ Links ] ARMBRECHT, I.; PERFECTO, I. 2003. Litter-twig dwelling ant species richness and predation potential within a forest fragment and neighboring coffee plantations of contrasting habitat quality in Mexico. Agriculture, Ecosystems and Environment 97: 107-115. [ Links ] ARMBRECHT, I.; GALLEGO, M. C. (en prensa). Testing ant predation on the coffee berry borer in shaded and sun coffee plantations in Colombia. Entomologia Experimentalis et Applicata. [ Links ] ARMBRECHT, I.; RIVERA, L.; PERFECTO, I. 2005. Reduced diversity and complexity in the leaf litter ant assemblage of Colombian coffee plantations. Conservation Biology 19 (3): 897-907. [ Links ] GALLEGO, M. A. 2004. La función depredadora de hormigas asociadas a cafetales cultivados bajo dos niveles de sombrío (Apía, Risaralda). Tesis de Maestría. Cali-Colombia, Universidad del Valle, Facultad de Ciencias, Departamento de Biología. 71 p. [ Links ] GALLEGO, M. C.; ARMBRECHT, I. 2005. Depredación por hormigas sobre la broca del café en cafetales cultivados bajo dos niveles de sombra en Colombia. Revista Manejo Integrado de Plagas, Costa Rica 76: 1-9. [ Links ] HAMMER, Ø.; HARPER, D.; RYAN, P. 2001. PAST: Paleontological statistics software package for education and data analysis. Palaeontología Electrónica 4: 1-9. [ Links ] HAYES, W. P. 1920. Solenopsis molesta Say (Hym.): A Biological Study. Kansas State Agricultural College Manhattan, Kansas. 55 p. [ Links ] HÖLLDOBLER, B.; WILSON, E. 1990. The Ants. Springer- Verlag: Estados Unidos. 732 p. [ Links ] KASPARI, M. 2000. A primer on Ant Ecology, pp. 9-24. En: Agosti, D.; Majer, J. D.; Alonso, L. E.; Shultz, T. R. (eds.). Ants: Standard methods for measuring & monitoring biodiversity. Smithsonian Institution. Washington and London. [ Links ] KELLER, L. 1988. Evolutionary implications of polygyny in the Argentine ant, Iridomyrmex humilis (Mayr) (Hymenoptera: Formicidae): an experimental study. Animal Behaviour 36: 159-165. [ Links ] KELLER, L.; PASSERA, L. 1989. Influence of the number of queens on nestmate recognition and attractiveness of queens to workers in the Argentine ant, Iridomyrmex humilis (Mayr). Animal Behaviour 37: 733-740. [ Links ] KELLER, L.; CHERIX, D.; ULLOA-CHACÓN, P. 1989. Description of a new artificial diet for rearing ant colonies as Iridomyrmex humilis, Monomorium pharaonis and Wasmannia auropunctata (Hymenoptera; Formicidae). Insectes Sociaux 36 (4): 348-352. [ Links ] MOGUEL, P.; TOLEDO, V. M. 1999. Biodiversity conservation in traditional coffee systems of Mexico. Conservation Biology 13 (1): 11-21. [ Links ] OKLAHOMA DEPARTMENT OF AGRICULTURE. 2005. The biology of red imported fire ants (Brochure). Web:http://www.ento.okstate.edu/fireants/rifabiology2.pdf. Fecha última consulta: [21 noviembre 2005]. [ Links ] RISCH, S. J.; CARROLL, R. 1982. Effect of a keystone predaceous ant, Solenopsis geminata, on arthropods in a tropical agroecosystem. Ecology 63 (6): 1979-1983. [ Links ] RIVERA, L. F. 2003. Diversidad de tres gremios de hormigas en cafetales bajo diferentes manejos de sombra. Tesis de pregrado. Cali-Colombia. Universidad del Valle. Facultad de Ciencias. 56 p. [ Links ] SWIFT, M. J.; VANDERMEER, J.; RAMAKRISHNAN, P. S.; ADERSON, J. M.; ONG, C. K.; HAWKINS, B. A. 1996. Biodiversity and Agroecosystem Function, pp. 261-298. En: Mooney, H. A.; Cushman, J. H.; Medina, E.; Sala, O. E.; Schulze, E. D. (eds.). Functional Roles of Biodiversity: A Global Perspective. UNEP. [ Links ] TRUNZER, B.; HEINZE, J.; HÖLLDOBLER, B. 1998. Cooperative colony founding and experimental primary polygyny in the ponerine ant Pachycondyla villosa. Insectes Sociaux. 45: 267-276. [ Links ] TSCHINKEL, W. R. 1988. Social control of egg-laying rate in queens of the fire ant, Solenopsis invicta. Physiological Entomology 13: 327-350. [ Links ] ULLOA-CHACÓN, P. 2003. Biología Reproductiva de Wasmannia auropunctata (R.) (Hymenoptera: Formicidae). Revista Academia Colombiana de Ciencias Exactas Físicas y Naturales. 27 (104): 441-447. [ Links ] VARÓN, E. H. 2002. Distribución espacio-temporal de hormigas con potencial como depredadoras de Hypothenemus hampei e Hypsipyla grandella, en sistemas agroforestales de café, en Costa Rica. Tesis de postgrado. Turrialba-Costa Rica, Centro Agronómico de Tropical de Investigación y Enseñanza (CATIE), Programa de Educación para el Desarrollo y la Conservación. 97 p. [ Links ] VÉLEZ, M.; BUSTILLO, A. E.; POSADA, F. J. 2001. Hormigas de la zona central cafetera y perspectivas de su uso en el control de Hypothenemus hampei (Ferrari) (Coleoptera: Scolytidae) En: XXVIII Congreso Sociedad Colombiana de Entomología. Libro de Resúmenes. Pereira, Colombia. p. 51. [ Links ] VÉLEZ, M.; BUSTILLO, A. E.; MONTOYA, E. C. 2003. Depredación de Hypothenemus hampei (Ferrari) por Solenopsis geminata y Gnamptogenys sp. (Hymenoptera: Formicidae) en: XXX Congreso Sociedad Colombiana de Entomología. Libro de Resúmenes. Cali, Colombia. p. 75. [ Links ] WAY, M. J.; KHOO, K. C. 1992. Role of Ants in Pest Management. Annual Review of Entomology. 37: 479-503. [ Links ] WHEELER, G. C.; WHEELER, J. 1976. Ant Larvae: Review and Synthesis. 7. The Entomological Society of Washington. 108 p. [ Links ] Recibido: 05-may-2006 - Aceptado: 26-abr-2007

