










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488On-line version ISSN 2665-4385
Rev. Colomb. Entomol. vol.33 no.2 Bogotá June/Dec. 2007
Sección Agrícola
Inhibidores de proteasas en papas resistentes y susceptibles a la mosca minadora Liriomyza huidobrensis (Diptera: Agromyzidae)
Protease inhibitors in potatoes resistant and susceptible to leaf the miner fly Liriomyza huidobrensis (Diptera: Agromyzidae)
JOSÉ OLIVERA1, MARÍA PALACIOS2, JORGE TENORIO3, ÁLVARO MARCELO4, ALÍ GOLMIRZAIE5
1 Biólogo, M. Sc. Centro de Investigación de Bioquímica y Nutrición (CIBN), Apto 1546 Lima-Perú. E-mail: joliverag@unmsm.edu.pe
2 Bióloga, M. Sc. Consultora Asociada, Agroconsult Internacional. E. Mail: mariapalacioslazo@viabcp.com
3 Biólogo, M. Sc. Centro Internacional de la Papa (CIP), Apto. 1558 Lima-Perú. E-mail: j.tenorio@cgiar.org
4 Ph. D. Facultad de Ciencias Biológicas, Universidad Nacional Mayor de San Marcos. Lima-Perú. E-mail: amarcelor@unmsm.edu.pe
5 Ph. D. Dept. of horticulture Fayetteville Arkansas, 72701 USA.E-mail: ali_golmirzaie@mtnsystems.com
Resumen: Se estudiaron las diferencias bioquímicas entre cuatro clones de papa resistentes (220LM87B, 662LM86B, 136LM86B y cultivar María Tambeña) y uno susceptible (cultivar Revolución) a la mosca minadora Liriomyza huidobrensis mediante comparación del grado de inhibición de la actividad proteolítica de la tripsina sintética y de los extractos de larvas de la mosca por acción de las proteínas totales extraídas de hojas dañadas de los clones. La actividad proteolítica, en unidades relativas, por efecto de los clones 662LM86B, 136LM86B, 220LM87B, María Tambeña, y Revolución sobre la actividad de la tripsina fue 1,7; 2,5; 1,8; 1,3 y 2,4 y sobre la actividad de los extractos de larvas fue 0,24; 0,4; 0.6; 0,1 y 0,5 respectivamente. Por otro lado, la inhibición de la actividad proteolítica de la tripsina por las proteínas de los cultivares María Tambeña y Revolución fue del 93% y 77% respectivamente. Finalmente, usando la técnica de zimografía se identificó una banda de inhibidores de proteasas de 105 KDa y otra de 63 KDa; el grado de intensidad de esas bandas fue mayor en los clones resistentes.
La banda de 105 kDa mostró una intensidad de hasta 4,2 veces en el cultivar Maria Tambeña con respecto a Revolución. Los resultados demuestran diferencias en la inhibición de la actividad proteolítica y en la presencia de la banda de 105 kDa entre los clones resistentes y susceptibles a la mosca minadora lo cual sugiere a los inhibidores de proteasas como factores importantes en la resistencia de estos clones a la mosca minadora.
Palabras clave: Solanum tuberosum. Defensa vegetal. Inhibidores de proteasas. Actividad proteolítica.
Abstract: Abstract Biochemical differences were studied among five potato clones: four resistant (220LM87B, 662LM86B, 136LM86B and cultivar María Tambeña) and one susceptible (cultivar Revolución) to attack of the leafminer fly Liriomyza huidobrensis (Diptera, Agromyzidae) through a comparison of the degree of inhibition of the proteolytic activity of synthetic trypsin and extracts of leafminer larvae by action of total proteins extracted from the leaves of the different potato clones before damage. The proteolytic activity, in relative units, of the clones 662LM86B, 136LM86B, 220LM87B, María Tambeña, and Revolution on trypsin activity were 1.7, 2.5, 1.8, 1.3, and 2.4, and on larval extracts were 0.24, 0.4, 0.6, 0.1 and 0.5, respectively. In contrast, when inhibition of the proteolytic activity of trypsin from proteins of the cultivars María Tambeña and Revolución was quantified, there was an inhibition of 93% and 77%, respectively. Finally, using zymography, two bands of protease inhibitors were identified, one 105 kDa and the other 63 kDa; the degree of intensity of these bands was greater in the resistant clones than the susceptible. The 105 kDa band showed an intensity up to 4.2 times higher in the cultivar María Tambeña than the cultivar Revolución.
The results demonstrate differences in the proteolytic activity and the presence of the 105 kDa band between the clones resistant and susceptible to the leafminer fly, which suggests that protease inhibitors are important factors in the resistance of these clones to the leafminer fly.
Key words: Solanum tuberosum. Plant defense. Protease inhibitor. Proteolytic activity.
Introducción
En la costa central de Perú, la mosca minadora Liriomyza huidobrensis Blanchard, 1926 es plaga clave del cultivo de la papa (Herrera 1963; Chávez y Raman 1987; Lizarraga 1990; Palacios y Raman 1993; Cisneros 1995). Al igual que en otras partes del mundo, como Argentina, Brasil, Chile, Costa Rica, Guatemala, México, Panamá, Israel, Indonesia y Egipto donde está reportada (CIP 1998), la mosca minadora se ha constituido en problema central como consecuencia del uso intensivo de plaguicidas para controlar otras plagas en este cultivo, por lo que ha desarrollado niveles de resistencia significativos a los insecticidas más comúnmente usados (CIP 1998). La plaga daña exclusivamente el follaje de las plantas: las hembras adultas pican las hojas para alimentarse y para depositar sus huevos, las larvas durante su desarrollo realizan minas serpenteantes en los foliolos, afectando de esta manera la capacidad fotosintética y el rendimiento (Palacios y Raman 1993). El daño en plantas jóvenes puede ocasionar mortalidad temprana (Morales et al. 1994). En el Valle de Cañete, la principal zona de producción de papa en la Costa Central del Perú, se han registrado pérdidas del rendimiento entre el 40 y 50 % cuando no se utilizaron métodos de control (Ewell et al. 1990; Palacios y Raman 1993).
Entre los factores que favorecen el incremento de las poblaciones de la mosca minadora en el Perú destacan el uso de variedades susceptibles y el excesivo uso de insecticidas (Palacios y Raman 1993). En la búsqueda de alternativas para el manejo de plagas en papa, el Centro Internacional de la Papa (CIP) desarrolló un programa de mejoramiento para obtener materiales resistentes a las principales plagas del cultivo: polilla de la papa (Phthorimaea operculella Zeller, 1873), mosca minadora (Liriomyza huidobrensis Blanchard, 1926) y gorgojo de los Andes (Premnotrypes spp) (Raman et al. 1994). Para la selección de plantas resistentes a mosca minadora se desarrolló un método de tamizado en campo, el cual, permitió identificar doce clones con altos niveles de resistencia a esta plaga y buenas características agronómicas (Tenorio et al. 1993). Posteriormente estos clones se evaluaron bajo condiciones de infestación controlada en laboratorio e invernadero, concluyéndose que el principal mecanismo de resistencia era el de antibiosis (Tenorio et al. 1993). Los clones 386768.10 (282LM86B) (actualmente es el cultivar “María Tambeña”), 220LM87B, 386766.43 (136LM86B) y 386767.21 (662LM86B) destacaron por su mayor nivel de resistencia (Tenorio et al. 1995).
Para defenderse de múltiples agresores, las plantas presentan complejas estrategias, como el uso generalizado de sustancias químicas (Fraenkel 1959). Estas sustancias pueden encontrarse en forma constitutiva en los tejidos, como por ejemplo, los glucósidos de cianógeno, terpenoides, alcaloides y sustancias fenólicas. Otras solo aparecen como respuesta a un estímulo primario por lo que son conocidas como sustancias inducidas, como por ejemplo las fitoalexinas y proteínas de defensa (Chrispeels y Sadava 1994). Entre las proteínas de defensa se mencionan a los inhibidores de proteasas, proteínas relacionadas con la patogénesis y lectinas (Bowles 1990; Dixon y Harrison 1994; Smirnoff 1998). El factor biótico más común en la aparición de sustancias inducidas es el estrés por herida, este es el caso del daño producido por las larvas y los adultos de la mosca minadora sobre las plantas de papa al causar en las hojas innumerables heridas externas e internas. Se arguye que el estrés por herida desencadena en la planta la activación coordinada de sus genes, no solo para la defensa de la planta sino también para la regeneración del tejido dañado (Sánchez- Serrano et al. 1993). En plantas de papa y de tomate, se ha observado que el daño en el follaje por herida induce en unas horas la acumulación de una serie de proteínas colectivamente denominadas Proteínas de Respuesta Sistémica a la Herida (PRSHs) (Bergey et al. 1996). Trabajos con plantas previamente heridas y utilizadas después como alimento para insectos han determinado alteraciones del comportamiento en estos insectos (Hodge et al. 1999). Cabe señalar que desde hace mucho tiempo los inhibidores de proteasas, dado que afectan la viabilidad de los insectos, son considerados factores importantes de resistencia (Broadway et al. 1986; Ryan 1990; Dymock et al. 1992; Barker et al. 1995). Además, se han aislado genes de diferentes fuentes biológicas con la intención de introducirlos en plantas para conferirles resistencia (Wolfson y Murdock 1995).
La inducción de las proteínas inhibidoras de proteasas por efecto de la herida ha sido estudiada más profundamente (Ryan 1990) y se ha propuesto que, en la ruta de señalización para producir estos componentes, la herida activa una fosfolipólisis específica en la membrana plasmática, liberando al citoplasma los ácidos linoleico y linolénico, poniendo en marcha la ruta del ácido jasmónico que ha de conducir a la activación y síntesis de dichos inhibidores (Farmer y Ryan 1992). En estudios donde se han medido los niveles de inhibidores de proteasas en el tejido de plantas resistentes a insectos, se ha encontrado una correlación entre la resistencia, la muerte o disuasión de los insectos y la presencia de los inhibidores de proteasas (Broadway et al. 1986; Ryan 1990). Se considera que el mecanismo de acción de estas proteínas inhibidoras de proteasas sobre los insectos se inicia en el intestino, con la activación de un sistema de retroalimentación que provoca la hipersecreción de enzimas digestivas y la disminución consecuente del apetito, produciéndose en el tiempo una pérdida de aminoácidos esenciales que ocasionan finalmente una deficiencia proteica (Broadway et al. 1986). Se ha observado que el nivel y la rapidez con que pueden presentarse los productos inducidos en las plantas, parecen marcar una diferencia característica en la resistencia de las mismas (Bowles 1990). Es decir, cuando se comparan cultivares resistentes y susceptibles, se observa en los resistentes la rápida acumulación de diferentes moléculas relacionadas con la defensa (Bowles 1990).
La finalidad entonces del presente estudio fue identificar diferencias en el nivel de inhibición de la actividad proteolítica e inhibidores de proteasas entre los clones de papa 220LM87B, 662LM86B, 136LM86B, la variedad María Tambeña y la variedad Revolución.
Materiales y Métodos
Material vegetal. Los clones de papa resistentes a mosca minadora utilizados fueron 220LM87B, 136LM86B, 662LM86B, y 282LM87B (actualmente el cultivar María Tambeña). El clon susceptible a la plaga fue el cultivar Revolución. Todos ellos se desarrollaron en condiciones de invernadero entre los meses de abril y julio, con un intervalo de temperatura de 16-22°C. Los tubérculos se sembraron en macetas de plástico de 21 cm de diámetro con una mezcla de 1:1:2 de arena, tierra y musgo.
Extracción de proteínas solubles
De las hojas de papa. Antes de la extracción, plantas de 20 días de edad se dañaron con un estilete hiriendo de tres a cuatro hojas de la parte inferior media y superior del follaje de la planta (daño mecánico). 24 horas después, se colectaron, se fijaron inmediatamente con nitrógeno líquido y se guardaron a –20°C en bolsas de plástico. En el laboratorio, estas hojas se pesaron y luego se molieron con nitrógeno líquido en un mortero, obteniéndose así un polvo el cual fue homogenizado con búffer fosfato (25 mM fosfato de sodio, pH 5,0, conteniendo 5% glicerol, 10 mM metabisulfito de sodio, y 2 mM ß-mercaptoetanol) en una proporción 1:7 (p/v). El homogenizado se centrifugó a 16,000 r.p.m. por 20 minutos y el sobrenadante se trató con acetona fría (1:1 v/v) para precipitar las proteínas. Todo el tratamiento se realizó en baño de hielo. El precipitado de proteínas se separó por centrifugación y se secó bajo presión reducida (Speed Vac).
De las larvas de mosca minadora. Se utilizaron larvas de mosca minadora del tercer estadio que miden en promedio 2,5 mm obtenidas de hojas de lechuga infestadas en campo. Las larvas se extrajeron cuidadosamente con un estilete y se almacenaron a – 20°C en tubos de microcentrifuga de 1,5 ml. Para la extracción de proteínas solubles se maceraron grupos de 20 larvas con 250 µl de búffer Tris (50 mM, pH 7 a 4°C) en baño de hielo, luego el homogenizado se centrifugó a 4,000 r.p.m. por 2 min y el sobrenadante se usó inmediatamente en las pruebas enzimáticas. La concentración de las proteínas se midió utilizando albúmina bovino (BSA) como proteína estándar siguiendo el método de Bradford (1976).
Ensayos Enzimáticos
Actividad proteolítica. Para determinar la actividad proteolítica de la tripsina sintética y de los extractos de larva se siguió el método enzimático de Fry et al. (1994) que utiliza gelatina como substrato de proteasas. Para esto se mezclaron 0,25 ml de gelatina al 1% (diluido en 50 mM de Tris-HCl, pH 8) y 0,25 ml de una solución enzimática: tripsina (20 mg/ml) o extracto de larvas (20 larvas con 250 ml de 50 mM Tris-Cl, pH 7,0 – 4 a 5 ug/ul). Posteriormente la mezcla se incubó a 30°C en tres tiempos: tres, cuatro y cinco horas. Al cabo del tiempo la reacción se detuvo con 0,75 ml de ácido tricloroacético (TCA) al 20%. Se dejó reposar por 15 min a temperatura ambiente y se centrifugó a 10.000 r.p.m. por 5 min. El sobrenadante se centrifugó nuevamente a la misma velocidad. La lectura en el espectrofotómetro se realizó a 280 nm y se utilizó como blanco la solución de gelatina combinado con tripsina, o con extracto de larva, respectivamente, pero incubados por separado y mezclados justo antes de la fijación con TCA. Una unidad de actividad proteolítica (U) se definió como el incremento de la absorbancia en 0,01 a 280 nm (Fry et al. 1994). Estos valores fueron considerados como el 100% de actividad proteolítica para los ensayos de inhibición.
Inhibición de la actividad proteolítica. Para los ensayos de inhibición de la actividad proteolítica de tripsina y de los extractos de larva, por efecto de las proteínas solubles de hojas de papa de los diferentes clones, se utilizó el método de Fry et al. (1994) descrito anteriormente. Previo al ensayo las soluciones enzimáticas se preincubaron con los extractos de proteínas de las hojas de papa (1 mg/ml diluidos en 25 mM de fosfato de sodio a pH 5) por 15 min a temperatura ambiente. En el caso de la mezcla de tripsina (20 ug/ml) con las proteínas de hojas de papa, ésta tuvo una proporción de 1:5 (v/v) y en el caso de los extractos de larva y proteínas de hoja, la proporción fue de 1:1 (v/v). Una unidad de actividad relativa (UR) se definió como el incremento de la absorbancia en 0,01 a 280 nm de la mezcla de enzima y proteínas de hoja sobre el incremento de la absorbancia en 0,01 a 280 nm de enzima sola.
Finalmente para comparar la inhibición proteolítica de la tripsina entre los cultivares M. Tambeña y Revolución se realizó un ensayo utilizando el substrato sintético TAME (Na-p-tosil- L-arginina-metil-ester) para la determinación de proteínas semejantes a tripsina según un método descrito en el Manual Whortington (1988) utilizando una concentración final de 4 ug/ml de tripsina y 10 mM de TAME. Para el cálculo de las unidades de actividad de tripsina se usó la fórmula Umg-1 = (5,55)(rA247/min)(mg-1 tripsina) (Wortington 1988).
Geles de poliacrilamida (Zimografía). Para detectar inhibidores de proteasas se prepararon geles de poliacrilamida (PAGE) al 8% copolimerizado con gelatina (1 mg/ml) (Hanspal et al. 1983). Se utilizó una unidad de electroforesis vertical (MINIPROTEAN II, Bio-rad) con espaciadores de 0,75 mm. Se pesaron 6 mg de proteínas de hojas por muestra las cuales se diluyeron en búffer muestra de electroforesis 2X (0,125 M tris- Cl, pH 6,8, conteniendo 20% glicerol, 4% de SDS (Lauryl sulfato), 1% de ß-mercaptoetanol y 0,002% de azul de bromofenol). Para el revelado los geles se tiñeron con una solución de azul de Coomassie R-250 (0,01%) disuelto en ácido acético:metanol:TCA:agua (24:24:2:10), y se destiñeron con metanol:ácido acético:agua (2:10:2). Simultáneamente se corrieron estándares de proteínas SDS-PAGE (Fosforilasa b, 97.400; Albúmina bovino, 66.000; Albúmina de huevo, 45.000; Anhidrasa Carbónica, 29.000, alfa-Lactoalbumina, 14.200), en otro gel con las mismas condiciones.
Análisis de geles y cálculos. Los geles se fijaron en membranas de celofán utilizando el búffer de decoloración de electroforesis como solución de fijación. Luego, se colocaron bajo un densitómetro (Hoefer) que estuvo conectado a una computadora PC IBM compatible desde el cual se registraron las bandas, y se analizaron mediante el programa GS365W.EXE (Hoefer Software). Las imágenes con bandas claras en fondo oscuro se invirtieron. Por otro lado, los datos obtenidos por los ensayos enzimáticos, a partir de tres repeticiones cada uno, se linearizaron mediante rectas de regresión con el programa Excel (Microsoft Inc.).
Resultados y Discusión
En este trabajo se asumió que las plantas responden de forma similar al ataque del insecto y al daño mecánico produciendo diferentes vías para iniciar la defensa (Farmer y Ryan 1992). Por lo tanto, se presume que algunos genes inducidos por herida, producen sustancias como los inhibidores de proteasas, que actúan en la defensa del hospedero contra el ataque del insecto.
Actividad proteolítica de la tripsina y de los extractos de larvas. La actividad proteolítica de la tripsina sobre gelatina fue de 1,8 unidades (U) y de los extractos de larva fue de 0,5 U. Cuando la tripsina se incubó con los diferentes extractos de papa, la actividad proteolítica aumentó en todos los casos produciéndose el mayor aumento con el extracto de 136LM86B (4,5 U) y menor con M. Tambeña (2,35 U) (Tabla 1). Este aumento de la actividad enzimática se interpretó como un sinergismo proteolítico, es decir, los valores son superiores al valor de la actividad proteolítica de la tripsina actuando sola (1,8 U). La mezcla de proteínas obtenidas de hojas indudablemente contiene muchas enzimas con actividad endopeptidasas sobre la gelatina, que es un sustrato muy utilizado para aislar proteasas de diferentes fuentes vegetales (Santaurius y Ryan 1977; Santarius y Belitz 1978) dado que los sustratos sintéticos son menos sensibles. Se puede notar que los clones resistentes presentan mayor actividad proteolítica. Giri et al. (1998) por ejemplo, encuentra diferencias entre dos cultivares de garbanzo resistentes y susceptibles a Fusarium oxysporum Schltdl. midiendo las unidades de actividad proteasa. En la interacción con los patógenos es conocida la respuesta hipersensible en la cual se inducen tempranamente y con rapidez varias enzimas hidrolíticas (Dixon y Harrison 1994). Dado que las larvas de la mosca minadora producen túneles dentro de las hojas, y por tanto las heridas que ocasiona pueden llegar a ser severas, la mayor síntesis de endopeptidasas podría estar dirigida a curar rápidamente las heridas producidas (Bowles 1990) o producir proteínas tóxicas como los inhibidores de proteínas (Bergey et al. 1996; Reymond et al. 2000).
Inhibición de la actividad proteolítica de tripsina y extractos de larvas. Al calcular la inhibición de la actividad enzimática de tripsina expresados en unidades relativas, en el cual los valores más cercanos a cero son considerados como de menor actividad proteolítica o de mayor inhibición proteolítica, se observó que María Tambeña y el clon 662LM86B tienen mayor inhibición con respecto al cultivar Revolución (1,3 UR y 1,7 UR respectivamente), mientras que el clon 136LM86B fue similar con el cultivar Revolución (Tabla 1). Dado que el sinergismo proteolítico de las hojas y la tripsina producen niveles elevados de absorbencia en forma diferente con los resultados de los extractos de larvas, se prefirió utilizar las unidades relativas para hacer una comparación del porcentaje de inhibición o reducción de la actividad proteolítica (Tabla 1). De esta forma es M. Tambeña la que presenta la mayor inhibición. Los inhibidores de proteasas inducidos por herida y por ataque de insectos que más se han estudiado son de la familia serina proteinasa (Ryan 1990; Bowles 1990; Sanchez-Serrano 1993). En este trabajo las hojas de papa fueron previamente dañadas con estilete y luego fijados con nitrógeno líquido para extraer las proteínas. Se conoce que por este efecto se sintetizan una serie de proteínas denominadas colectivamente proteínas de respuesta sistémica a la herida (Bergey et al. 1996) entre las cuales están los inhibidores de serina proteinasas. Actualmente se sabe que esta inducción puede ser modulada sistémicamente por un péptido de 18 aminoácidos llamado sistemina (Ryan 1998; Shaller 1999). Al parecer, M Tambeña contiene muchos de esos inhibidores que contrarrestan la actividad de tripsina (una serina preoteinasa).
La inhibición proteolitica de María Tambeña y Revolución se corroboró en el ensayo realizado con TAME. El ensayo de tripsina y TAME presenta una actividad de 363,88 unidades por miligramo, pero cuando la tripsina se combina con las proteínas de hojas de M. Tambeña y Revolución esta actividad disminuyó en 93% y 77% respectivamente (Tabla 2). En términos generales los clones resistentes presentan mayor inhibición de la actividad proteolítica de los extractos de larvas de mosca minadora y de tripsina en comparación con el cultivar susceptible Revolución. Siendo M. Tambeña la que produce las mayores inhibiciones.

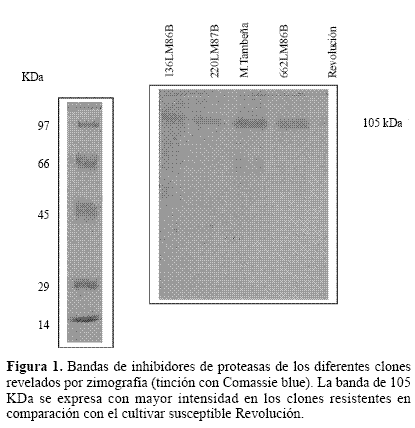
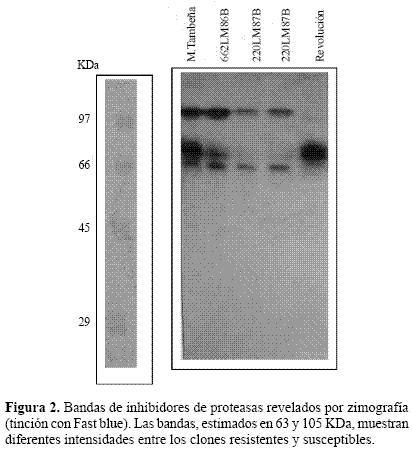
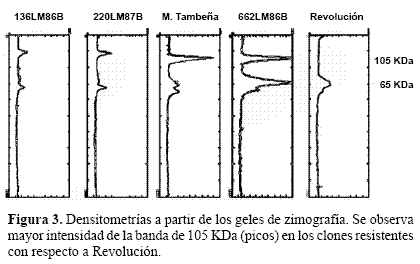
Inhibidores de proteasas mediante zimografía. Los geles de electroforesis para ver actividad de proteínas con base en su función (zimografía) son técnicas ampliamente utilizadas para estudiar extractos crudos de diverso origen, como los inhibidores de proteasas (Hansal 1983; Michaud 1993). Estos métodos se basan en la capacidad de la proteína de perder su actividad cuando se expone a alta concentración de SDS pero lo recupera una vez que el detergente es removido de la solución. En nuestro caso usamos el método de Hanspal (1983) donde se encontró una banda de aproximadamente 105 KDa que se expresa en diferentes intensidades entre los clones resistentes, siendo de menor intensidad en el material susceptible Revolución (Fig. 1). Otra banda menos definida, de aproximadamente 63 KDa se muestra también en estos geles (Fig. 2). El grado de intensidad de la banda de 105 KDa que es la de mayor tinción, se midió por densitometría registrándose en cada uno de los genotipos picos mayores que en Revolución. En M Tambeña el nivel de expresión es de 4.2 a uno en comparación con Revolución (Fig. 3). Es interesante que la intensidad de la banda de 105 kDa este correlacionada con los clones de papa resistentes a la mosca minadora. A pesar que también Revolución presenta la banda de este inhibidor, no alcanza la mayor producción presentada en los clones resistentes. Estos resultados nos inducen a considerar que la identificación de la naturaleza molecular de la banda de 105 KDa para el desarrollo de marcadores bioquímicos de resistencia podría utilizarse para facilitar la identificación de materiales que expresen esta cualidad.
Se asume que las plantas perciben la depredación el insecto en forma similar a como perciben la herida, y por tanto utilizan las mismas señales de traducción en cascada, o vías similares para iniciar la respuesta de la defensa (Farmer y Ryan 1992; Hodge et al. 1999), por tanto, se presume que algunos genesinducibles por herida, como los que codifican inhibidores de serina proteinasas, probablemente funcionan en la defensa vegetal contra la depredación del insecto. Este resultado junto con la observación del incremento de mayores proteasas detectadas en los materiales resistentes, en especial M. tambeña, podrían revelarnos el origen de la resistencia. Sin embargo, no hay antecedentes que los insectos puedan inducir inhibidores de proteasas debido a la depredación (Ryan 1990). Pero si hay reportes de enzimas como bromelaina (Boller 1986), leucina aminopeptidasa (Pautot et al. 1993), proteinasas semejantes a subtilisina (Jorda et al. 1999), carboxipeptidasa (Schaller y Ryan 1996) y cisteina proteinasa (Linthorst et al. 1993) que se acumulan en respuesta al ataque de un insecto, del mismo modo que produce una herida. La inducción de inhibidores de proteasas y el conjunto de proteasas si es reportado como una respuesta al daño mecánico en plantas (Farmer 1992; Reymond 2000).
Conclusiones
Tomando en conjunto los resultados se puede postular que los clones de papa resistentes a la mosca minadora, en especial el cultivar Maria Tambeña y el clon 662LM86B sintetizan o activan en corto tiempo los niveles de sus proteínas frente al daño provocado por el ataque de la mosca minadora en sus hojas, entre los cuales están los inhibidores de proteasas. Esta inducción diferencia en términos generales los clones resistentes del susceptible al ataque de mosca minadora y podría ser el factor principal de la resistencia.
Literatura Citada
BARKER, A. M.; WRATTEN, S.D.; EDWARDS, P. J. 1995. Woundinduced changes in tomato leaves and their effects on the feeding patterns of larval Lepidoptera. Oecologia 101: 251-257. [ Links ]
BERGEY, D. R.; HOWE, G. A.; RYAN, C. A. 1996. Polypeptide signaling for plant defensive genes exhibits analogies to defense signaling in animals. Proceedings National Academy of Sciences (USA). 93: 12053-12058. [ Links ]
BOLLER, T. 1986. Roles of proteolytic enzymes in interactions of plants with other organism. Vol, 2, M.J. Dalling, ed (Boca Raton, FL; CRC Press), pp 67-95. En: Plant Proteolytic Enzymes. [ Links ]
BOWLES, D. J. 1990. Defense-Related Proteins in Higher Plants. Annual Review of Biochemistry 59: 873-907. [ Links ]
BRADFORD, M. M. 1976. A rapid and sensitive method for the quantification of microgram quantities of protein using the principle of protein-dye binding. Analytical Biochemistry 72: 248- 254. [ Links ]
BROADWAY, R. M.; DUFFEY, S. S.; PEARCE, G.; RYAN, C. A. 1986. Plant Proteinase Inhibitors: A defense against herbivorous insects?. Entomologia Experimentalis et Applicata 41: 33-38. [ Links ]
CIP. 1998. Biotechnology-Assisted breeding to reduce pesticide sse in potatoes. 1993-1997 (GLO-91-016). Final Report. International Potato Center and United Nations Development Program Division of Global and Interregional Projects. Lima, Perú. 48 p. [ Links ]
CISNEROS, F. H. 1995. Control de Plagas Agrícolas. AGLIS Electronics. Lima, Perú. 313 p. [ Links ]
CHAVEZ, G. L.; RAMAN, K. V. 1987. Evaluation of trapping and trap types to reduce damage to potatoes by the leafminer, Liriomyza huidobrensis (Diptera, Agromyzidae). Insect Science and its Application 6: 265-270. [ Links ]
CHRISPEELS, M. J.; SADAVA, D.E. 1994. Plants, genes and agriculture. Jones and Bartlett Publishers Inc. Boston, USA. 562 p. [ Links ]
DIXON, R. A.; HARRISON, M. J. 1994. Early Events in the Activation of Plant Defense Responses. Annual Review of Phytopathology 32: 479-501. [ Links ]
DYMOCK, J. J.; LAING, W. A.; SHAW, B. D.; GATEHOUSE, A. M. R.; CHRISTELLER, J. T. 1992. Behavioral and physiological responses of grass grub larvae (Costelytra zealandica) feeding on protease inhibitors. New Zealand Journal of Zoology 19: 123- 131. [ Links ]
EWELL, P. T.; FANO, H.; RAMAN, K. V.; ALCAZAR, J.; PALACIOS, M.; CARHUAMACA, J. 1990. Farmer Management of Potato Insect Pest in Peru. Food Systems Research Series No. 6. International Potato Center (CIP). Lima, Perú. 77 p. [ Links ]
FARMER, E. E.; RYAN, C. A. 1992. Octadecanoid precursors of jasmonic acid active the synthesis of wound-inducible proteinase inhibitors. The Plant Cell 4: 129-134. [ Links ]
FRAENKEL, G. 1959. The raison d'être of secondary plant substances. Science 129: 1466-1470. [ Links ]
FRY, S. M.; HUANG, J. S.; MILHOLLAND, R. D. 1994. Isolation and preliminary characterization of extracellular protease produced by strains of Xylella fastidiosa from grapevines. Phytopathology 84: 357-363. [ Links ]
HANSPAL, J. S.; BUSHELL, G. R.; GHOSH, P. 1983. Detection of Protease Inhibitors Using Substrate-Containing Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis. Analytical Biochemistry 132: 288-293. [ Links ]
HERRERA, A. J. 1963. Problemas insectiles del cultivo de la papa en el Valle de Cañete. Revista Peruana de Entomología 6 (1): 1- 9. [ Links ]
HODGE, S.; WRATTEN, S.; SMIYH, J.; THOMAS, M.; FRAMPTON, C. 1999. The role of leaf wounding and epigeal predator on caterpillar damage to tomato plants. Annals of Applied Biology 134: 137-141. [ Links ]
JORDA, L.; COEGO, A.; CONEJERO, V.; VERA, P. 1999. A genomic cluster containing four differentially regulated subtilisinlike processing protease genes is in tomato plants. Journal of Biological Chemistry 274: 2360-2365. [ Links ]
LINTHORST, H. J. M., VAN DER DOES, C. BREDERODE, F. T.; BOL, J. F. 1993. Circadian expression and induction by wounding of tobacco genes for cysteine proteinase. Plant Molecular Biology 21: 685-694. [ Links ]
LIZARRAGA, A. D. 1990. Biología de la Mosca Minadora Liriomyza huidobrensis, Blanchard (Diptera, Agromyzidae). Revista Latinoamericana de la Papa 3: 30-40. [ Links ]
METHA, R. A.; WARMBART, R. D. MATTO, A. K. 1996. Tomato fruit carboxypeptidase: Properties, induction upon wounding, and immunocytochemical localization. Plant Physiology 110: 883- 892. [ Links ]
MICHAUD, D.; FAYE, L.; YELLE, S. 1993. Electrophoretic analysis of plant cysteine and serine proteinases using gelatin-containing polyacrylamide gels and class-specific proteinase inhibitors. Electrophoresis 14: 94-98. [ Links ]
MORALES, R. A.; ATENCIO, F. A.; LARA, J. A.; MUÑOZ, J. A. 1994. La mosquita minadora (Liriomyza spp. ) en Panamá. Volcan, Chiriqui (Panamá). Programa Regional Cooperativo de Papa (PRECODEPA) Instituto de Investigación Agropecuaria de Panamá (IDIAP). Monografía No. 1. Panamá. 21 p. [ Links ]
PALACIOS, M.; RAMAN, K. V. 1993. Control integrado de la mosca minadora Liriomyza huidobrensis Blanchard. Memorias. Taller de manejo Integrado de Plagas. p. 48-55. Balcarce. [ Links ]
PAUTOT, V.; HOLZER, F. M. REISCH, B., WAILLING, L. L. 1993. Leucine aminopeptidase: An inducible component of the defense response in Lycopersicon esculetum (tomato). Proceedings of the National Academy of Sciences USA 90: 9906-9910. [ Links ]
RAMAN, K. V.; GOLMIRZAIE, A. M.; PALACIOS, M.; TENORIO, J. 1994. Inheritance of Resistance to Insects and Mites. pp. 447- 463. En: Brashaw, J. E. and Mackay G. R. (eds.). Potato Genetics. CAB International. Wallingford, UK. 552 p. [ Links ]
REYMOND, P.; WEBER, H., DAMOND, M.; FARMER, E. 2000. Differential gene expression in response to mechanical wounding and insect feeding in Arabidopsis. The plant Cell 12: 707-719. [ Links ]
RYAN, C. A. 1990. Protease inhibitors in plants: genes for improving defenses against insects and pathogens. Annual Review of Phytopathology 28: 425-449. [ Links ]
SANCHEZ-SERRANO, J. J.; ANATI, S.; DAMMANN, C.; ENETH, M.; HERBERS, K.; HILDMAN, T.; LORORTH, R.; PRATT, S.; WILLNITZRE, L. 1993. Proteinase Inhibitors in the potato response to wounding, pp. 157-173. En: Chet, I. (ed.). Biotechnology in plant disease control. Wiley-Liss. New York, USA. [ Links ]
SANTARIUS K.; RYAN, C. 1977. Radial diffusion as a sensitive method for screening endopeptidase activity in plant extracts. Analytical Biochemistry 77: 1-9. [ Links ]
SANTARIUS K.: BELITZ, H. D. 1978. Proteinase activity in potato plants. Planta 141: 145-153. [ Links ]
SCHALLER, A. RYAN, C. A. 1996. Molecular cloning of a tomato leaf cDNA encoding an aspartate protease, a systemic wound response protein. Plant Molecular Biology 31: 1073-1077. [ Links ]
SMIRNOFF, N. 1998. Plant resistance to environmental stress. Current Opinions in Biotechnology 9: 214-219 [ Links ]
TENORIO, J.; PALACIOS, M.; GOLMIRZAIE, A. M.; CISNEROS, F.; ORTIZ, O. 1993. Evaluación preliminar de clones resistentes a Liriomyza huidobrensis en el Valle del Tambo, Arequipa. Resúmenes. XXXV Convención Nacional de Entomología, p.18. Arequipa, Perú. [ Links ]
TENORIO, J.; PALACIOS, M.; CAÑEDO, V.; GOLMIRZAIE, A.M.; VELARDE, H. 1995. "María Tambeña-INIA" una variedad resistente a mosca minadora Liriomyza huidobrensis. Resúmenes. XXXVIII Convención Nacional de Entomología, p.15. Chincha, Perú. [ Links ]
WOLFSON, J. L.; MURDOCK, L. L. 1995. Potential use of protease inhibitor for host plant resistance: A test case. Environmental Entomology 24: 52-57. [ Links ]
WORTHINGTON, C. C. 1988. Manual Worthington. Worthington Biochemical Corporation. New Jersey, USA. 346 p. [ Links ]
Recibido: 13-jun-2006 - Aceptado: 9-ago-2007


{kind=link}