










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488On-line version ISSN 2665-4385
Rev. Colomb. Entomol. vol.33 no.2 Bogotá June/Dec. 2007
Sección Morfología, Comportamiento, Ecología y Sistemática
Avispas no polinizadoras asociadas a Ficus andicola (Moraceae) en la Cordillera Central de Colombia
Non-pollinating fig wasps associated with Ficus andicola (Moraceae) in the Central Andes of Colombia
WILLIAM CARDONA1, PATRICIA CHACÓN DE ULLOA2, GUSTAVO KATTAN3
1 Autor para correspondencia: Biólogo, Universidad del Valle. Fundación EcoAndina/WCS - Programa Colombia. Apartado Aéreo 25527 Cali, Colombia. williamcardona@gmail.com
2 Bióloga, Ph. D. Departamento de Biología, Universidad del Valle. Apartado Aéreo 25360 Cali, Colombia. pachacon@uniweb.net.co
3 Biólogo, Ph. D. Fundación EcoAndina/WCS - Programa Colombia. Apartado Aéreo 25527 Cali, Colombia. gkattan@wcs.org
Resumen: En el interior de las inflorescencias (síconos) de los Ficus se pueden encontrar varias especies de avispas que se desarrollan. Entre estos insectos se encuentra el polinizador, que por lo general es una sola especie, mientras que las otras avispas no ofrecen ningún beneficio al árbol y por el contrario pueden producir efectos negativos sobre el número de semillas y polinizadores producidos. En este trabajo se estudió la composición de la fauna de avispas no polinizadoras asociadas a Ficus andicola en la Cordillera Central de Colombia. Se examinaron 418 síconos que fueron recolectados de 12 árboles; las avispas que emergieron de cada uno de ellos se contaron y separaron por morfoespecies. El 46,5% de las flores de F. andicola produjeron semillas y el 53,5% avispas. Se encontraron 22 morfoespecies de avispas no polinizadoras emergiendo de los síconos de F. andicola. Las ocho más frecuentes fueron identificadas a nivel de género. Las morfoespecies más abundantes, la cuales estuvieron presentes en más del 40% de los síconos, fueron en su orden, Idarnes sp.1, Heterandrium sp. e Idarnes sp.2. La mayoría de las morfoespecies fueron encontradas en menos del 10% de los síconos. Basados en curvas de acumulación de especies las 22 morfoespecies encontradas representan el 88% de las esperadas para F. andicola.
Palabras clave: Avispas de los Ficus. Agaonidae. Pegoscapus. Idarnes. Heterandrium.
Abstract: Several species of wasps may develop inside the inflorescences (figs) of Ficus spp. These species include the pollinator, usually a single species, while the other species do not provide any benefits to the tree; instead, they may affect negatively the number of seeds and pollinators produced. In this work we studied the composition of the nonpollinating fig wasp fauna associated with Ficus andicola in the Central Cordillera of the Colombian Andes. We examined 418 figs that were collected from 12 trees; the wasps that emerged from each one were counted and separated into morphospecies. A total of 46.5% of flowers produced seeds and 53.5% produced wasps. Twenty-two morphospecies were found emerging from figs of F. andicola. The eight most abundant were identified to genus. The most abundant morphospecies, which were present in more than 40% of figs, were Idarnes sp1, Heterandrium sp. and Idarnes sp2. Most wasp species were found in less than 10% of figs. Based on species accumulation curves, the 22 morphospecies found represent 88% of the expected number of species for F. andicola.
Key words: Fig wasps. Agaonidae. Pegoscapus. Idarnes. Heterandrium.
Introducción
Entre los árboles del género Ficus y las avispas de la familia Agaonidae existe una relación mutualista de tipo obligado. Esta situación se presenta porque dichas avispas son las únicas que pueden polinizar las flores de los Ficus; las avispas a su vez solo pueden desarrollarse en el interior de las flores femeninas de estos árboles (Janzen 1979; Wiebes 1979; Ramírez 1974, 1976). La inflorescencia de los Ficus es un receptáculo cerrado en forma globular denominado sícono. Solo hay una vía de acceso a su interior, a través de un pequeño túnel formado por escamas estrechamente superpuestas, que se encuentran en su parte apical formando el ostiolo (Berg 1989; Galil y Eisikowitch 1968). Los únicos insectos con adaptaciones estructurales y morfológicas que les permiten pasar a través del ostiolo son las avispas de la subfamilia Agaoninae, por lo tanto son las únicas que pueden tener contacto directo con las flores y polinizarlas (Janzen 1979; Wiebes 1979; Ramírez 1976, 1974). Anteriormente se creía que la relación entre Ficus y agaonidos era altamente específica, de tal manera que, por cada especie de Ficus había una especie de agaonido (Ramírez 1970). Sin embargo, nuevos estudios han encontrado algunas excepciones y actualmente se conocen varios casos en los que una especie de Ficus puede ser polinizada por más de una especie de avispa, y una avispa puede polinizar más de una especie de Ficus, aunque es más frecuente el primer caso (Cook y Rasplus 2003; Molbo et al. 2003; Michaloud et al. 1996; Kerdelhue et al. 1997).
La polinización del árbol y la reproducción de las avispas se da a través de un ciclo sincronizado (Galil y Eisikowitch 1968). Cuando las flores femeninas están receptivas, las avispas hembras entran cargadas de polen, pasando a través del ostiolo, proceso en el cual pierden sus alas, partes de las patas y las antenas (Ramírez 1976; Wiebes 1979). Las flores femeninas de los Ficus presentan una gradación continua en el tamaño del estilo (Bronstein 1988; Otero y Ackerman 2002), las avispas ponen sus huevos introduciendo su ovipositor a través de este, pero solo pueden poner huevos en las que tienen el estilo más corto que sus ovipositores; las flores en las que no pueden poner los huevos, por tener el estilo largo, son las que producen las semillas (Ramírez 1976; Wiebes 1979; Herre 1989; Nefdt y Compton 1996). Después de la oviposición, las hembras mueren en el interior del sícono. Las larvas inducen agallas en los ovarios, allí llevan a cabo la fase de desarrollo alimentándose del tejido de los óvulos (Galil y Eisikowitch 1968). Cuando las semillas y las avispas han culminado su fase de desarrollo, los machos emergen, fertilizan a las hembras y abren huecos en la pared del sícono por donde las hembras pueden salir. A continuación emergen las hembras, para este momento las flores masculinas están maduras, por lo tanto las hembras tienen contacto con el polen antes de abandonar el sícono en busca de otro que esté receptivo para empezar de nuevo el ciclo (Herre 1989; Galil y Eisikowitch 1968).
Dentro de los síconos se desarrollan otras avispas que nunca tienen contacto directo con las flores femeninas y por lo tanto no las polinizan (Janzen 1979). Estas avispas pertenecen a las familias Agaonidae, Eurytomidae, Orymidae, Pteromalidae y Torymidae y se les conoce como avispas no polinizadoras de los Ficus (Boucek 1993). Pueden ser desde simples comensales, hasta parásitos que afectan el número de semillas y avispas polinizadoras que se desarrollan en los síconos. El número de especies de avispas asociadas a una especie de Ficus puede variar entre 3 y 30 especies (Bronstein 1991; Compton y Nefdt 1990; Compton y Hawkins 1992; Kerdelhue y Rasplus 1996; Boucek et al. 1981). Estas avispas dependen de la presencia del polinizador para su desarrollo ya que por lo general los síconos en los que no están presentes son abortados por el árbol. Además, la mayoría de las avispas usan los huecos abiertos en la pared del sícono por el polinizador para poder abandonarlo (Bronstein 1991; Kerdelhue y Rasplus 1996; West et al.1996). Las avispas no polinizadoras de los Ficus pueden ser ubicadas en tres grupos funcionales (Cook y Rasplus 2003): 1) avispas que inducen agallas grandes y sus parasitoides, estas avispas son mucho más grandes que los polinizadores y ovipositan desde afuera, durante o antes de la polinización. Algunas inducen agallas grandes en la pared interna del sícono y las otras son parasitoides de estas. Se presentan en pocos síconos y en baja cantidad. 2) avispas que inducen agallas pequeñas, también ovipositan desde afuera durante o después de la polinización. Inducen agallas en las flores femeninas y están presentes en la mayoría de los síconos en medianas a grandes cantidades.3) parásitos internos: están presentes sólo en el viejo mundo y entran a través del ostíolo al tiempo que lo hace el polinizador. Ovipositan en las flores femeninas y son encontradas en pocos síconos, pero pueden ser muy abundantes.
Existen aproximadamente 750 especies de Ficus en el mundo, de las cuales cerca de 125 se encuentran en América; la mayoría de los Ficus de la región neotropical están asociados con bosques húmedos de tierras bajas (Berg 1989). La mayoría de los estudios han sido enfocados sobre esas especies y solo se conoce la fauna asociada a una fracción de ellas (Wiebes 1995; Herre 1996; Schiffler 2002; West y Herre 1994; West et al. 1996). En Colombia no se conoce la fauna de avispas asociadas a ninguna especie de Ficus, pues hasta el momento los trabajos realizados en el país se han enfocado en su biología floral (Martínez 1981; Ramos 1985; Largacha 1997). Ficus andicola Standl, es una especie de tamaño mediano que crece principalmente en los bordes del bosque y en zonas abiertas, la máxima altura que alcanza es de aproximadamente 15m y es una especie de Ficus muy abundante (L. Valenzuela datos sin publ.). Los síconos de F. andicola son pedunculados, con brácteas basales carnosas las cuales permanecen hasta que el fruto madura (Ramos 1985); su especie polinizadora es Pegoscapus aff. silvestre, la cual presenta un marcado dimorfismo sexual, las hembras son de color negro, con alas transparentes y cabeza en forma de pala, mientras que los machos son de color amarillo pálido transparente, carecen de alas, poseen órgano copulador retráctil, cabeza grande, mandíbulas fuertes, el primer par de patas es cavador y el segundo par casi vestigial (Ramos 1985; Jansen y Sarmiento en prensa). El objetivo de este trabajo fue determinar la composición de la fauna de avispas no polinizadoras asociadas a F. andicola como un aporte al conocimiento de este sistema para futuros estudios sobre su ecología y relaciones.
Materiales y Métodos
El trabajo se llevó a cabo entre los meses de febrero y octubre de 2004 en el Santuario de Fauna y Flora Otún Quimbaya (4°43’43”N, 75°34’42”W), localizado en el municipio de Pereira, Risaralda, en la vertiente occidental de la Cordillera Central, a elevaciones entre los 1.800 y 2.100 m (Londoño 1994). El régimen de lluvias es bimodal, con valores máximos en abril y octubre y promedio anual de 2.500 mm; la temperatura promedio anual es de 15ºC (Aguilar y Rangel 1994). El área se encuentra cubierta por un mosaico de bosque natural maduro poco intervenido, bosque secundario formado por regeneración natural y plantaciones monoespecíficas de urapán (Fraxinus chinensis) y roble (Quercus humboldtii) (Galeano y Bernal 1994).
Inicialmente se buscaron y marcaron los individuos de F. andicola en la zona de estudio y se les hizo seguimiento para observar la producción de frutos. Se colectaron entre 31 y 35 síconos de cada árbol en la fase en la que las avispas habían empezado a emerger, pero que aún no abandonaban el sícono.Los síconos colectados se llevaron al laboratorio, se abrieron longitudinalmente y posteriormente se pusieron en frascos plásticos individuales tapados con una malla de tela, que a su vez estaba sujetada al frasco con una banda elástica.
Una vez las avispas habían terminado de emerger (dos o tres días después) se colectaron y conservaron en etanol al 70%. Posteriormente las avispas se separaron y se identificaron a género las más frecuentes, utilizando la clave de Boucek (1993). Las morfoespecies fueron separadas usando las características más evidentes, como por ejemplo tamaño, longitud del ovipositor y patrones de coloración. Las semillas se separaron raspando la pared del sícono, se dejaron secar por 24 horas y posteriormente se contaron en su totalidad a través de un microscopio estereoscópico.
Se evaluó la eficiencia del muestreo mediante la obtención de curvas de acumulación de especies, usando el programa EstimateS 6.0b1 (Colwell 2000), con 100 aleatorizaciones. Aunque todos los estimadores presentaron un comportamiento similar se utilizó el no paramétrico Chao 2 por ser considerado uno de los menos sesgados (Colwell y Coddington 1994). Éste estima el número de especies esperadas considerando la relación entre el número de especies únicas y el número de especies duplicadas (Moreno 2001).
Resultados
De 96 árboles de F. andicola registrados en la zona de estudio durante 10 meses, se muestrearon 12 que produjeron síconos. En total se colectaron 418 síconos, con un promedio de 35 por árbol. De todos los síconos recolectados emergieron avispas.
El 53,5% de las flores de F. andicola desarrollaron avispas, mientras que el 46,5% restante produjeron semillas. El número promedio de avispas polinizadoras no difirió significativamente del número de semillas producidas por cada sícono (t = - 0,86, g. l. = 417, P > 0,05). Por otra parte, el número total de avispas (polinizadoras y no polinizadoras) fue significativamente mayor que el número de semillas producidas (t = - 4,16, g. l. = 417, P < 0,001) (Tabla 1).

Las avispas no polinizadoras estuvieron presentes en 11 de los 12 árboles estudiados y en el 73% de los síconos. El número promedio de avispas no polinizadoras por árbol fue altamente variable (Tabla 2). Se encontraron algunos árboles con alto número de avispas no polinizadoras (hasta 44 individuos), mientras que otros presentaron números muy bajos (un individuo).
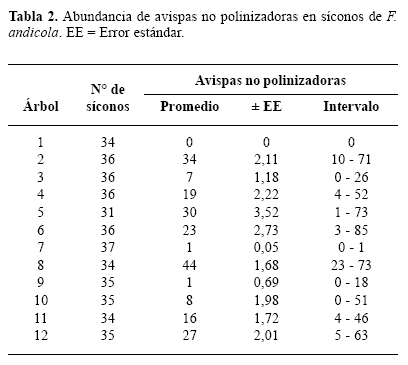
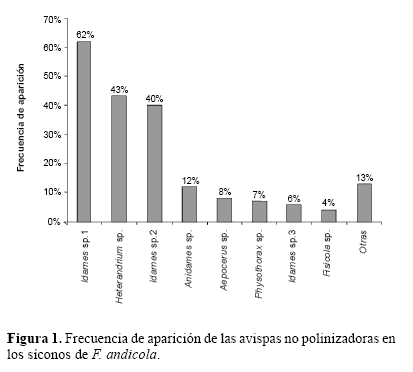
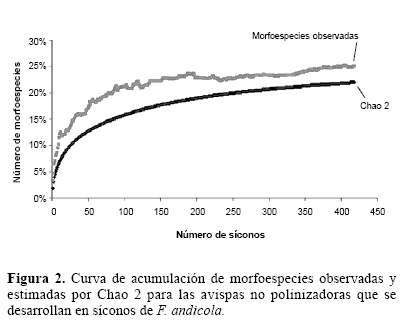
Se encontraron 22 morfoespecies de avispas en los síconos de F. andicola, pertenecientes a tres subfamilias de Agaonidae y a las familias Torymidae y Pteromalidae. Las ocho morfoespecies más frecuentes fueron identificadas hasta el nivel de género (Tabla 3), estas representan el 99% de todas las avispas no polinizadoras encontradas y en conjunto estuvieron presentes en el 73% de los síconos, es decir en todos los que produjeron avispas no polinizadoras. Sólo cuatro morfoespecies de avispas no polinizadoras estuvieron presentes en más del 10% de los síconos. De estas sólo Idarnes sp. 1, Idarnes sp. 2 y Heterandrium sp. emergieron de más del 40% de los síconos. La única morfoespecie que emergió de más del 50% de los síconos fue Idarnes sp. 1 (Fig. 1). De acuerdo al grupo funcional, el mayor número de avispas no polinizadoras fueron las que inducen agallas pequeñas, presentes en aproximadamente el 70% de los síconos. Las avispas que inducen agallas grandes estuvieron presentes en el 19% y sus parasitoides, en este caso Physothorax sp., solo en el 7%. La curva de acumulación de especies muestra que se encontró aproximadamente el 88% de las morfoespecies esperadas para F. andicola (Fig. 2).
Discusión
Se encontraron 22 morfoespecies de avispas no polinizadoras desarrollándose en una alta proporción de los síconos de F. andicola. Este número se refiere a las morfoespecies y es posible que representen un número menor de especies. De cualquier manera esta cantidad está dentro del intervalo conocido de avispas asociadas a una especie de Ficus. Compton y Hawkins (1992) encontraron que el número puede variar entre tres y 30 para 24 especies de Ficus estudiados en África. Compton y Nefdt (1990) reportan cinco especies asociadas a F. burtt-davyi en Sur África, mientras que en Costa de Marfil, Kerdelhue y Rasplus (1996) encontraron en F. sur y F. vallischoudae 10 y cuatro especies respectivamente. Boucek et al. (1981) encontraron 28 especies de avispas no polinizadoras asociadas a los síconos de F. thonningii en Zimbabwe. Bronstein (1991) registró tres especies en los síconos de F. pertusa en Costa Rica, mientras que en Brasil se encontraron tres géneros de avispas asociados a F. mexiae (Schiffler 2002). Es posible que el alto número de morfoespecies encontradas en este estudio se deba al esfuerzo de muestreo, que como lo muestra la figura 2, permitió registrar el 88% de las esperadas. La mayoría de las morfoespecies encontradas, solo estuvieron presentes en menos del 4% de los síconos. Esto podría implicar que estas solo sean oportunistas y que no dependen únicamente de F. andicola para su reproducción. Las morfoespecies más comunes, probablemente si tengan un mayor grado de especificidad y de dependencia sobre esta especie. Dentro del género Ficus existen tanto especies monoicas como ginodioicas, estas últimas desarrollan flores masculinas y femeninas en algunos árboles y sólo flores femeninas en los otros (Janzen 1979; Wiebes 1979). En el Neotrópico únicamente se encuentran especies de Ficus monoicos (Berg 1989). En general se ha encontrado que las especies de Ficus monoicas presentan un mayor número de especies de avispas no polinizadoras y una mayor tasa de parasitismo (Patel 1998; Weiblen et al. 2001).
Se encontró un alto número de avispas del género Idarnes, que ha sido reportado como el más común de las avispas no polinizadoras en los Ficus del subgénero Urostigma, al cual pertenece F. andicola (Gordh 1975; West y Herre 1994; West et al.1996). El género Heterandrium ha sido encontrado con poca frecuencia en los Ficus del subgénero Urostigma y es poco lo que se conoce de su biología y del efecto que puede causar sobre los miembros del mutualismo (West et al.1996). Otras especies menos abundantes son las formadoras de agallas grandes y sus parasitoides (Aepocerus, Physothorax y Ficicola), las cuales fueron encontradas en bajas proporciones en este estudio. El grupo funcional al cual pertenece cada morfoespecie de avispa no polinizadora, juega un importante papel en las relaciones y ecología del sistema. Por ejemplo las avispas del género Idarnes (grupo carme) son formadoras de agallas en las flores femeninas, estas por lo tanto compiten con los polinizadores por lugares de oviposición y se ha encontrado que pueden afectar negativamente su número (West y Herre 1994). Las avispas del género Heterandrium, utilizan el mismo recurso pero no suelen ser abundantes. En este caso fue la segunda morfoespecie más frecuente y por lo tanto es posible que pueda causar un efecto similar a las avispas del género Idarnes. Las formadores de agallas grandes, en este caso las del género Aepocerus, Idarnes (grupo incerta) y Anidarnes, cuando están presentes en grandes números pueden también tener efecto negativo sobre las semillas o los polinizadores. Para el caso de Aepocerus, cuando ha sido encontrado en cantidades importantes, los resultados sugieren que pueden tener efectos negativos sobre el número de semillas y polinizadores (West et al. 1996). Los parasitoides de las formadoras de agallas grandes, pueden tener importancia en la dinámica poblacional de sus hospederos pero no necesariamente tendrían un efecto directo sobre los polinizadores o las semillas. En este caso se encontró una baja proporción de avispas del género Physothorax, las cuales son parasitoides de Aepocerus spp.
Normalmente los síconos en los cuales las flores no han sido polinizadas son abortados por el árbol, pero se ha observado que algunas especies de avispas no polinizadoras pueden inducir la maduración de los frutos (Bronstein 1991; Kerdelhue y Rasplus 1996; West et al. 1996). En el presente estudio se encontraron dos síconos en los que, a pesar de que no habían ingresado avispas polinizadoras, fueron retenidos y se desarrollaron avispas no polinizadoras dentro de ellos. Estos dos síconos tenían en común a Anidarnes sp., Ficicola sp. y Aepocerus sp., lo que sugiere que una de estas tres morfoespecies puede inducir la maduración del fruto. Por lo observado en otros estudios, se esperaría que dicha morfoespecie sea Aepocerus sp., puesto que otras especies de este género inducen la maduración de los síconos no polinizados en F. popenoei, F. dugandi y F. trigonata en Panamá (West et al. 1996). Por el método utilizado no se pudo saber si estas avispas hubieran podido abandonar el sícono sin la presencia de polinizadores. Sin embargo, se pudo observar, como ya había sido sospechado por West et al. (1996), que los machos de Aepocerus sp. pueden abrir huecos en la pared del sícono. Además, se observó que estos huecos son abiertos desde el interior de la agalla en la que se desarrollaron.
Una de las preguntas centrales en el estudio del sistema Ficus - Agaonidae es cómo éste ha podido perdurar a través del tiempo a pesar de que entre los miembros del mutualismo existe un conflicto de intereses (Machado et al. 2001). Este conflicto se da porque las avispas se desarrollan en las flores femeninas a expensas de las semillas, por lo tanto, un aumento en el número de polinizadores disminuye el número de semillas producidas (Herre y West 1997). Sin embargo, este sistema que surgió hace aproximadamente 90 millones de años (Machado et al. 2001) aún perdura, lo que indica que existen mecanismos que regulan la producción de avispas, evitando que el sistema colapse (West y Herre 1994; Nefdt y Compton 1996). El patrón general observado en especies de Ficus de tierras bajas, es que la proporción de flores de las que emergen polinizadores está entre 0,45 y 0,55 (Herre 1989). Para F. andicola la proporción también fue de 0,45, es decir, que de alguna manera el árbol conserva aproximadamente la mitad de las flores para la producción de semillas. Las flores en las que se desarrollan las hembras polinizadoras pueden verse como una inversión, ya que estas son las encargadas de dispersar el polen y por lo tanto representan la función masculina del árbol. El verdadero costo está representado por la potencial pérdida de semillas o hembras polinizadoras, debida a la producción de machos polinizadores y especies de avispas que no proveen el servicio de polinización (Herre 1989).
El número de avispas no polinizadoras tiene una alta variación entre árboles, esta variación es común y se ha encontrado en otras especies de Ficus (West et al. 1996). Dicho patrón podría ser explicado por variaciones locales en la abundancia de avispas no polinizadoras y de síconos receptivos y por la fenología de fructificación de sus árboles hospederos (Patel 1998; Nefdt y Compton 1996; West et al. 1996). Para el caso de F. andicola estas variaciones locales también estarían controlando dichas diferencias. Por ejemplo, si hay muchos árboles en los que estén emergiendo las avispas (fase masculina), entonces habrá un mayor número de avispas disponibles para ovipositar en los síconos que se encuentren receptivos (fase femenina). Por otra parte, el número de síconos receptivos también tendría un efecto determinante, si hay muy pocos en un momento en el que haya un alto número de avispas, entonces habrá una mayor cantidad ovipositando en los síconos. La alta variación en el número de avispas no polinizadoras podría ser una explicación para que las avispas polinizadoras mantengan una población viable. En este caso, los árboles que no reciben avispas no polinizadoras estarían actuando como fuente de polinizadores, mientras que los árboles con alto número de no polinizadores lo harían como sumidero.
A pesar de que el sistema Ficus - Agaonidae ha sido muy estudiado en algunas localidades, todavía quedan muchos aspectos por explorar, sobre todo acerca del papel que las avispas no polinizadoras juegan. Ficus andicola representa un buen modelo para futuros estudios ya que es una especie abundante y de fácil acceso, además posee una amplia fauna de avispas de la que se podría estudiar la dinámica de sus interacciones. También ofrece la oportunidad de conocer más sobre la biología de las especies de avispas no polinizadoras, como por ejemplo la del género Heterandrium, que ha sido poco frecuente en los estudios anteriores, pero en este caso resultó ser la segunda morfoespecie más abundante.
Agradecimientos
Este trabajo fue posible gracias al apoyo financiero de US Fish and Wildlife Service y John D. and Catherine T. MacArthur Foundation. Agradecemos a la Unidad de Parques Nacionales Naturales por su apoyo logístico en el Santuario de Fauna y Flora Otún Quimbaya.
Literatura Citada
AGUILAR, M.; RANGEL, J. O. 1994. Clima del Parque Regional Natural Ucumarí y sectores aledaños, pp. 59-84. En: Rangel, J. O. (ed.). Ucumarí un caso típico de la diversidad biótica Andina. Pereira: Corporación Autónoma Regional de Risaralda (CARDER). 451 p. [ Links ]
BERG, C. C. 1989. Classification and distribution of Ficus. Experientia 45: 605-611. [ Links ]
BOUCEK, Z.; WATSHAM, A.; WIEBES, J. T. 1981. The fig wasp fauna of the receptacles of Ficus thonningii (Hymenoptera, Chalcidoidea). Tijaschrift Voor Entomologie 124: 149-234. [ Links ]
BOUCEK, Z. 1993. The genera of chalcidoid wasps from Ficus fruit in the New World. Journal of Natural History 27: 173-217. [ Links ]
BRONSTEIN, J. L. 1988. Mutualism antagonism and the fig pollinator interaction. Ecology 69: 1298-1302. [ Links ]
BRONSTEIN, J. L. 1991. The nonpollinating wasp fauna of Ficus pertusa: exploitation of a mutualism? Oikos 61: 175-186. [ Links ]
COLWELL, R. K.; CODDINGTON J. A. 1994. Estimating terrestrial biodiversity through extrapolation. Philosophical Transactions of the Royal Society of London Series B 345: 101-118. [ Links ]
COLWELL, R. K. 2000. EstimateS: Statistical estimation of species richness and shared species from samples. Version 6.0b1. http:// viceroy.eeb.uconn.edu/estimates. [ Links ]
COMPTON, S. G.; NEFDT, R. J. C. 1990. The figs and fig wasps of Ficus burtt-davyi. Mitteilungen aus den Institut für Allgemeine Botanik Hamburg 23: 441-450. [ Links ]
COMPTON, S. G.; HAWKINS, B. A. 1992. Determinants of species richness in southern African fig wasp assemblages. Oecologia 91: 68-74. [ Links ]
COOK, J. M.; RASPLUS, J.-Y. 2003. Mutualists with attitude: coevolving fig wasps and figs. Trends in Ecology and Evolution 18 (5): 241-248. [ Links ]
GALEANO, M.; BERNAL, J. 1994. Composición florística del Parque Regional Natural Ucumarí, pp. 141-187. En: Rangel, J. O. (ed.). Ucumarí un caso típico de la diversidad biótica andina. Pereira: Corporación Autónoma Regional de Risaralda (CARDER). 451 p. [ Links ]
GALIL, J.; EISIKOWITCH, D. 1968. On the pollination ecology of Ficus sycomorus in East Africa. Ecology 49 (2): 259-269. [ Links ]
GORDH, G. 1975. The comparative external morphology and systematics of the neotropical parasitic fig wasp genus Idarnes (Hymenoptera: Torymidae). The University of Kansas Science Bulletin 50 (9): 389-455. [ Links ]
HERRE, E. A. 1989. Coevolution of reproductive characteristics in 12 species of New World figs and their pollinator wasps. Experientia 45: 637-647. [ Links ]
HERRE, E. A. 1996. An overview of studies on a community of Panamian figs. Journal of Biogeography 23: 593-607. [ Links ]
HERRE, E. A.; WEST S. A. 1997. Conflict of interest in a mutualism: documenting the elusive fig wasp-seed trade-off. Proceedings of the Royal Society of London 264: 1501-1507. [ Links ]
JANSEN-G., S.; SARMIENTO C. E. (en prensa). A new species of high mountain Andean fig wasp (Hymenoptera: Agaonidae: Agaoninae) with a detailed description of its life cycle. Symbiosis 45. [ Links ]
JANZEN, D. H. 1979. How to be a fig? Annual Review of Ecology and Systematics 10: 13-51. [ Links ]
KERDELHUE, C.; RASPLUS, J.-Y. 1996. Non-pollinating Afrotropical fig wasps affect the fig pollinator mutualism in Ficus within the subgenus Sycomorus. Oikos 75: 3-14. [ Links ]
KERDELHUE, C.; HOCHBERG, M. E.; RASPLUS, J-Y. 1997. Active pollination of Ficus sur by two sympatric fig wasp species in West Africa. Biotropica 29 (1): 69-75. [ Links ]
LARGACHA, A. 1997. Biología floral de Ficus insipida Willd (Moraceae). Trabajo de grado, Universidad del Valle, Cali, Colombia. [ Links ]
LONDOÑO, E. 1994. Parque Regional Natural Ucumarí. Un vistazo histórico, pp. 13-21. En: Rangel, J. O. (ed.). Ucumarí un caso típico de la diversidad biótica andina. Pereira: Corporación Autónoma Regional de Risaralda (CARDER). 451 p. [ Links ]
MACHADO, C. A.; JOUSSELIN, E.; KJELLBERG, F.; COMPTON, S. G.; HERRE, E. A. 2001. Phylogenetic relationships, historical biogeography, and character evolution of fig pollinating wasps. Proc. R. Soc. Lond. B 268: 685-694. [ Links ]
MARTÍNEZ, L. 1981. Aspectos biológicos y químicos del sícono de Ficus gigantosyce Dugand. Trabajo de grado, Universidad Nacional, Bogotá, Colombia. [ Links ]
MICHALOUD, G.; CARRIERE, S.; KOBBI, M. 1996. Exceptions to the one: one relationship between African fig trees and their fig wasp pollinators: possible evolutionary scenarios. Journal of Biogeography 23: 513-520. [ Links ]
MOLBO, D.; MACHADO, C. A.; SEVENSTER, J. G.; KELLER, L.; HERRE, E. A. 2003. Cryptic species of fig-pollinating wasps: Implications for the evolution of the fig-wasp mutualism, sex allocation, and precision of adaptation. Evolution 100 (10): 5867- 5872. [ Links ]
MORENO, C. E. 2001. Manual de métodos para medir la diversidad. Universidad Veracruzana. Xalapa, México. 49 p. [ Links ]
NEFDT, R. J. C.; COMPTON, S. G. 1996. Regulation of seed and pollinator production in the fig - fig wasp mutualism. Journal of Animal Ecology 65: 170-182. [ Links ]
OTERO, J. T.; ACKERMAN, J. D. 2002. Flower style length and seed production in two species of Ficus (Moracea) in Puerto Rico. Caribbean Journal of Science 38 (3-4): 249-251. [ Links ]
PATEL, A. 1998. Wasp composition in inflorescences of two dioecious fig species-implications for the fig-wasp mutualism. Biotropica 30 (3): 474-480. [ Links ]
RAMÍREZ, W. 1970. Host specificity of fig wasps (Agaonidae). Evolution 24 (4): 680-691. [ Links ]
RAMÍREZ, W. 1974. Coevolution of Ficus and Agaonidae. Annals of the Missouri Botanical Garden 61: 770-780. [ Links ]
RAMÍREZ, W. 1976. Evolution of blastophagy. Brenesia 9: 1-13. [ Links ]
RAMOS, J. 1985. Biología floral de Ficus andicola Standl., Ficus schippi Standl. y Ficus velutina H. B. ex Willd (Moracea) en el Valle del Cauca. M. S. Tesis. Universidad Nacional, Bogotá, Colombia. [ Links ]
SCHIFFLER, G. 2002. Fig wasps (Hymenoptera: Agaonidae) associated to Ficus mexiae Standl (Moraceae) in Lavras, Minas Gerais, Brazil. Neotropical Entomology 31 (4): 653-655. [ Links ]
WEIBLEN, G. D.; YU, D. W.; WEST, S. A. 2001. Pollination and parasitism in functionally dioecious figs. Proceedings of the Royal Society of London 268: 651-659. [ Links ]
WEST, S. A.; HERRE, E. A. 1994. The ecology of the New World fig-parasitizing wasps Idarnes and implications for the evolution of the fig pollinator mutualism. Proceedings of the Royal Society of London 258: 67-72. [ Links ]
WEST, S. A.; HERRE, E. A.; WINDSOR, D. M.; GREEN, P. R. S. 1996. The ecology and evolution of the New World nonpollinating fig wasp communities. Journal of Biogeography 23: 447-458. [ Links ]
WIEBES, J. T. 1979. Co-evolution of figs and their insect pollinators. Annual Review of Ecology and Systematics 10: 1-12. [ Links ]
WIEBES, J. T. 1995. The New World Agaoninae (pollinators of figs). Verhandelingen Koninklijke Nederlandse Akademie Wetenschappen 94: 1-60. [ Links ]
Recibido: 4-ene-2007 - Aceptado: 18-oct-2007


{kind=link}