










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488On-line version ISSN 2665-4385
Rev. Colomb. Entomol. vol.33 no.2 Bogotá June/Dec. 2007
Sección Morfología, Comportamiento, Ecología y Sistemática
Diferenciación morfológica y molecular de especies de crisópidos (Neuroptera: Chrysopidae)
Morphological and molecular differentiation of lacewing species (Neuroptera: Chrysopidae)
PATRICIA CADENA1, FERNANDO ÁNGEL2, LUIS A. GÓMEZ3, RANULFO GONZÁLEZ4
1 Autor por correspondencia: Cenicaña, Programa de Variedades, calle norte #3BN -110 Apartado Aéreo 9138 Cali, Colombia, tel: 2 60 66 11 ext. 5151. A.A. 9138, Cali, Colombia. pcgoyes@yahoo.com
2 Biotecnólogo. Cenicaña, Programa de Variedades. Apartado Aéreo 9138 Cali, Colombia. fangel@cenicana.org
3 Entomólogo. Cenicaña, Programa de Variedades. Apartado Aéreo 9138 Cali, Colombia. lagomez@cenicana.org
4 Entomólogo. Universidad del Valle, Departamento de Biología. Apartado Aéreo 25360 Cali, Colombia ranulfo@univalle.edu.co
Resumen: En el Valle del Cauca (Colombia), se estudió la distribución e identificación de las especies más abundantes de crisópidos en zonas donde se cultiva la caña de azúcar (Saccharum spp.) con mayor intensidad. Así, se han determinado las especies que podrían ser utilizadas para contrarrestar el efecto del pulgón amarillo Sipha flava (Hemiptera: Aphididae) sobre la producción total de la caña de azúcar. Aunque las especies y morfotipos del presente estudio son separados utilizando caracteres morfológicos, se evaluó la validez de dichos caracteres. Para esto, se realizó una diferenciación morfológica mediante genitalia interna y una diferenciación molecular, empleando la técnica PCR-RFLP sobre la región 18S del ADN ribosomal, de 11 especies de tres géneros de la familia Chrysopidae (Ceraeochrysa, Chrysoperla y Leucochrysa), algunas de las cuales podrían ser importantes como agentes de control biológico del pulgón amarillo de la caña de azúcar, S. flava. Los caracteres empleados resultaron ser útiles para distinguir las especies. En el caso de la genitalia interna, mostraron diferentes patrones de las estructuras. En el caso de los datos moleculares, las especies fueron diferenciadas utilizando solo una combinación de iniciadores NS para la amplificación del ADN y la digestión del amplificado producida con una enzima de restricción (NS5-NS6+DraI). Esta información se adiciona al conocimiento taxonómico que se tiene sobre la familia, además puede ayudar a resolver problemas sistemáticos al interior de los géneros.
Palabras clave: Genitalia. ADN ribosomal. Taxonomía. Subunidad 18S.
Abstract: In the Cauca Valley (Colombia), the distribution and identification of the most abundant lacewing species were studied in zones where sugar cane (Saccharum spp.) is cultivated. It has therefore been possible to determine the species that could be used to counter the effect of the yellow aphid Sipha flava (Hemiptera: Aphididae) on the overall production of sugar cane. Although the lacewing species and morphotypes in the current study can be separated using morphological characters, the validity of those characters was studied. To do that, we established a morphological differentiation using internal genitalia and a molecular differentiation using a PCR-RFLP analysis of the 18S rDNA region of 11 species from three genera of the family Chrysopidae (Ceraeochrysa, Chrysoperla, and Leucochrysa), some of whom could be important biological control agents of the yellow sugar cane aphid S. flava. The characters used proved to be useful for distinguishing the species. In the case of internal genitalia, there were different patterns of the structures. In the case of molecular data, the species were differentiated using only one combination of NS primers for the DNA amplification, followed by digestion of fragments with a restriction enzyme (NS5-NS6+DraI). These results add to the taxonomic understanding of the family, and may help resolve systematic problems within the genera.
Key words: Genitalia. ribosomal DNA. Taxonomy. 18S rDNA region.
Introducción
El pulgón amarillo Sipha flava (Forbes, 1884) (Hemiptera: Aphididae) es una plaga de alta incidencia en el rendimiento total de la caña de azúcar. De acuerdo con el uso de diferentes alternativas para su manejo, se han considerado depredadores de la familia Chrysopidae, algunos de los cuales se encuentran en la caña de azúcar, mientras que otros se localizan en diferentes cultivos.
Chrysopidae es una de las familias más grandes del orden Neuroptera y de las más importantes en términos económicos. En los bosques tropicales húmedos, los crisópidos sobrepasan a los mirmeleóntidos en riqueza de especies, mientras que estos últimos presentan más especies en zonas áridas (Penny 2002). Brooks y Barnard (1990) citaron cerca de 1200 especies y subespecies reconocidas, clasificadas en 86 géneros y subgéneros. Este grupo de insectos se encuentran dentro de los más eficaces depredadores, debido a que la larva se alimenta de huevos, larvas y adultos de una gran diversidad de hemípteros fitófagos y de otros insectos relativamente sedentarios (Penny 2002). Algunas larvas de esta familia presentan proyecciones torácicas y setas largas, que usa para sujetar los exoesqueletos secos de su presa y desechos sobre su cuerpo, lo cual le sirve para protegerse de predadores y parásitos. Los adultos se alimentan de otros insectos, néctar y polen (Penny 2002). Se ha registrado que las larvas de Ceraeochrysa cubana (Hagen, 1861) y Chrysoperla externa (Hagen, 1861) se alimenta agresivamente de 13 y 21 plagas de cultivos importantes respectivamente. De los ocho géneros de crisópidos que se han encontrado en los trópicos, Chrysoperla y Ceraeochrysa, parecen ser los que poseen el potencial más grande para usarse en control biológico (Albuquerque et al. 2001).
La taxonomía de esta familia no es muy estable ya que muchas especies constantemente son descritas y las especies previamente descritas son reexaminadas, facilitando asignaciones genéricas y reconociendo sinonimias. En la región Neotropical, la gran mayoría de especies fueron descritas por Banks y Navás, sin utilizar genitalia; además muchas de sus especies, se describieron con base sólo en hembras, o sobre un espécimen tipo que carecía de abdomen (Adams y Penny 1987). Ramírez (2002) estableció que las especies de Chrysopidae más frecuentes y más ampliamente distribuidas en el zona de estudio en el Valle del Cauca fueron Ceraeochrysa claveri (Navás, 1911), Ceraeochrysa cubana, Chrysoperla externa; las especies nativas con mayor capacidad de depredación de individuos de pulgón amarillo fueron Ceraeochrysa cubana, Ceraeochrysa claveri, Chrysoperla externa y Leucochrysasp. 2.
Aunque existen claves de determinación taxonómica para la familia Chrysopidae, la diferenciación de las especies requiere un examen cuidadoso pues algunos géneros son muy uniformes morfológicamente, pero su eficiencia en el manejo de determinada plaga puede ser tal que se presenten diferencias significativas que determinen el éxito de un programa de control. La introducción de especies determinadas incorrectamente puede constituir un serio problema si el insecto a controlar en el programa de control biológico no está asociado con la especie usada o no se adapta a las condiciones locales del sitio de liberación (De Almeida y Stouthamer 2003).
Dado que las especies de Chrysopidae encontradas en el Valle del Cauca poseen características morfológicas externas variables, se consideraron otros métodos de diferenciación, tales como marcadores moleculares y marcadores morfológicos como la genitalia interna, con miras a corroborar la validez de los caracteres morfológicos externos, además para esclarecer la determinación de algunos morfotipos que no han sido reconocidos hasta especie, contribuyendo al conocimiento taxonómico de la familia.
Comparada con la morfología externa, la variabilidad interespecífica de la genitalia es tan grande y característica, que algunos autores no han ilustrado otras partes del cuerpo. La gran divergencia en la morfología de la genitalia entre las especies es la regla en insectos, y en la mayoría de los grupos; los taxónomos usan la genitalia (especialmente la genitalia del macho) para la determinación de las especies y su clasificación. Un artículo de la revista alemana de entomología Tijdschrisft voor Entomologie, mostró que de 26 artículos taxonómicos, la genitalia juega un papel prominente en 23 (Schilthuizen 2003).
Por otro lado, la utilización de marcadores moleculares es ampliamente usada para diferenciar especies muy relacionadas y en muchos casos sólo una región del ADN es usada, dependiendo de los propósitos de la investigación (Caldeira et al. 2003). Recientemente, diferentes regiones del ADN ribosomal han sido propuestas para ser usadas en estudios moleculares como una herramienta adicional a la caracterización morfológica. Este método incluye el análisis de la región 18S, y las regiones ITS1 (Internal Transribed Spacer) e ITS2 del ADN ribosomal a través de la reacción en cadena de la polimerasa, la cual utiliza iniciadores que amplifican estas regiones. Sin embargo, debido al alto polimorfismo intraespecífico y a los patrones complejos que se producen con este método, se puede ver restringido su uso (Vidigal et al. 2002).
Combinada con el uso del ADN ribosomal, la técnica PCRRFLP permite diferenciar especies muy cercanas y en ocasiones prácticamente indistinguibles con un análisis morfológico (Luchetti et al. 2005). Esta es una técnica simple y rápida, que puede ser usada como una herramienta auxiliar en la determinación de las especies (Vidigal et al. 2002). Estas técnicas pueden ser más útiles al momento de diferenciar entre especies de un grupo particular de cierta zona, cuando se pretende evaluar la dispersión de un parasitoide liberado y estudiar el parasitismo siguiendo las liberaciones, estudiar la composición de especies hermanas, y para diferenciar especies muy relacionadas (Silva et al. 1999).
Se planteó la necesidad de realizar una diferenciación molecular dada su utilidad como un medio rápido y eficaz para encontrar marcadores específicos (Arnold et al. 1991; Wilkerson 1995), así como una diferenciación morfológica a partir de genitalia interna, con miras a establecer marcadores diagnósticos de las especies, para luego evaluar que tanto se correlacionan estos datos con los obtenidos a partir de la morfología externa; esta aproximación podría arrojar resultados más confiables sobre el estatus de las especies de estudio. El conocimiento que se obtenga, contribuye a la información que sobre el manejo de plagas se está generando y, en este caso, sobre el manejo del pulgón amarillo, buscando un tratamiento integral que conlleve una reducción de su incidencia en la producción comercial de la caña de azúcar.
Materiales y Métodos
Material de estudio. Se trabajó tanto con especies criadas en el laboratorio de entomología del Centro de Investigación de la Caña de Azúcar de Colombia CENICAÑA, como especies recolectadas en el campo. Se realizaron recolecciones manuales de crisópidos (Neuroptera: Chrysopidae) en lotes de caña de azúcar, yuca y cítricos en CENICAÑA (3°21'51”N; 76°18'1”W) y en lotes de maíz en sitios aledaños, en lotes de caña en el ingenio MAYAGÜEZ (3°23'40”N; 76°19'54”W), y en lotes de maíz y yuca ubicados en el Centro Internacional de Agricultura Tropical CIAT (3°30'27”N; 76°21'26”W). Las recolecciones se hicieron entre julio de 2004 y julio de 2005. El estudio se realizó con 11 especies: Ceraochrysa sp. 1, Ceraeochrysa sp. 2, Ceraeochrysa sp. 3, Ceraeochrysa cubana, Ceraeochrysa claveri, Chrysoperla sp. 1, Chrysoperla carnea (exótica), Chrysoperla rufilabris (exótica), Leucochyrsa sp. 1, Leucochyrsa sp. 2, y Leucochyrsa sp. 3 y se analizaron ejemplares tanto de campo como de laboratorio. Se usaron diez ejemplares por especie (cinco machos y cinco hembras), para un total de 110. Debido a que la abundancia relativa de algunas especies no era suficiente para obtener todos los individuos que se necesitaban tanto para la estandarización como para el análisis final, fue necesario criar algunas especies.
Determinación de las especies a partir de caracteres morfológicos. Se diferenciaron las especies a partir de caracteres de la morfología externa, usando la clave de Ramírez (2002). Antes de realizar las pruebas moleculares, también se verificó la diferenciación de las especies y morfotipos, usando los dibujos de las estructuras de genitalia interna de machos disponibles, tanto los de la literatura como los realizados durante el estudio.
Análisis molecular
Extracción de ADN. Para la extracción de ADN se utilizó el tórax y parte del abdomen (aproximadamente siete segmentos) de cada uno de los ejemplares. Se siguió el método de extracción de Cheung et al. (1993) y se modificó de acuerdo con la cantidad de tejido a utilizar, que en este caso dependía del tamaño del insecto. El tejido se maceró en 300μl de tampón de extracción frío, y se adicionaron 75 μl de solución Sarcosyl 5% para precipitar polisacáridos (Sharma et al. 2002). La suspensión se incubó por una hora en un baño maría a una temperatura de 65°C, invirtiéndose cada 10 minutos. Las muestras se centrifugaron a 14.000 rpm por seis minutos a 20°C. Se pasó el sobrenadante a otro tubo y se le adicionó 168,75 μl de acetato de amonio 10M y 300 μl de isopropanol frío, invirtiéndose varias veces y dejándolo a -20°C por dos horas, para precipitar el ADN. Posteriormente las muestras se centrifugaron a 14000 rpm por 10 minutos a 20°C, se descartó el sobrenadante dejando el botón (ADN) y se lavó con 300 μl de etanol 70% (-20°C), invirtiendo el tubo varias veces, teniendo cuidado de no desprender el precipitado. Se dejó secar por 20 minutos aproximadamente a temperatura ambiente hasta que el etanol se evaporara y se resuspendió en 100 μl de tampón TE. Finalmente se agregó a cada muestra 1,5 μl de ribonucleasa A (10 mg/ml) y se incubó a 37°C por 20 minutos. El ADN se conservó a -20°C hasta su utilización.
Condiciones de la PCR. La PCR (reacción en cadena de la polimerasa) se llevó a cabo en un termociclador PTC-100 (Programmable Termal Controller) de MJ Research, utilizando un volumen total de 12,5 μl, 17,5 ng de ADN, Tampón 1X, dNTPs 0,2 mM, Iniciador A 0,5 μM, Iniciador B 0,5 μM, BSA 0,1 μg, Taq polimerasa 1U, ajustado a 12,5 μl con H2O. Las condiciones para realizar la amplificación del ADN ribosomal fueron: un paso de denaturación inicial a 94°C de 2 minutos, 40 ciclos de denaturación a 94°C por 30 segundos, hibridación a 57°C por 1 minuto, 30 segundos, extensión a 72°C por 30 segundos y un paso de extensión final a 72°C por 10 minutos, después del último ciclo. En este caso se realizó un ajuste en el perfil térmico de la PCR en el paso de hibridación.
Selección de combinación de iniciadores NS. Para amplificar diferentes regiones de la subunidad 18S del ADN ribosomal, se evaluaron 52 combinaciones de iniciadores NS (Nuclear Small rDNA) y digestiones usando varias enzimas de restricción, en 11 muestras de ADN de los crisópidos (una por especie). De las 52 combinaciones, finalmente se seleccionaron seis para evaluar la diferenciación entre todas las especies, utilizando 10 muestras de ADN por especie (Tabla 1). Los amplificados de ADN ribosomal se separaron por electroforesis en geles de agarosa al 1,2% y las digestiones posteriores fueron separadas en geles de agarosa al 1,7% utilizando un voltaje de 100V en tampón TAE 1X; los geles se prepararon en tampón TAE 1X (Tris-ácido glacial ácetico 0.04M; EDTA 0.001M), usando Bromuro de Etidio (0,4 ìg/ml para la tinción) (Sambrook et al. 1989). La longitud de los productos de amplificación se estimó por comparación con un marcador Lambda (ë) digerido con la enzima PstI. Las electroforesis se realizaron en una cámara horizontal Marca Gibco BRL, modelo Horizon 20-25.

Análisis morfológico. Se siguió el método propuesto por Henry et al. (1993), pero utilizando el mismo tratamiento para machos y hembras. La porción de tejido se colocó en KOH (10%), y se dejó en ebullición de cinco a 10 minutos. Después se lavó con bastante agua destilada y se tiñó con dos gotas de chlorazol black E (Sigma) (5%) dejándolo de dos a tres minutos. Finalmente se lavó con agua destilada y se disectó. Las genitalias se conservaron en viales con glicerina.
Identificación y esquematización de las estructuras de la genitalia interna. Se emplearon estructuras de la genitalia interna, tanto de machos como de hembras, fáciles de observar y reconocer, es decir que permitieran una determinación rápida de las especies; gonarcus y gonapsis/tignum en el caso de los machos, y espermateca para las hembras. Se realizaron ilustraciones de todas las estructuras empleando un microscopio SWIFT con un aumento 100X y un micrómetro ocular. La identificación de las estructuras y su posterior esquematización se hicieron empleando como guía diferentes artículos (Adams 1977; Henry 1983; Nuñez 1988; Penny 2002; De Freitas 2003; Brooks y Barnard 1990; Henry et al. 1993; Tauber et al. 2000; Kang-Zhen et al. 2004) que se refieren a la genitalia interna de algunos géneros de la familia Chrysopidae.
Resultados y Discusión
Análisis molecular
Extracción de ADN. El método de extracción usado permitió obtener la cantidad necesaria de ADN, tanto para la estandarización como para el análisis final con una cantidad total promedio de 28,9 μg. Este método, también ha sido empleado exitosamente para la extracción de ADN de otros artrópodos como avispas del género Trichogramma (Hymenoptera: Trichogrammatidae) (Landry et al. 1993) y varias especies de arañas de diferentes familias (Linyphiidae, Theridiidae y Clubionidae) (A´Hara et al. 1998).
Amplificación de ADN ribosomal y selección de iniciadores NS. Las diferentes combinaciones de iniciadores NS permitieron la amplificación de regiones específicas de la subunidad 18S del ADN ribosomal de las especies de crisópidos. Aunque las combinaciones de iniciadores mostraban bandas intensas y reproducibles, casi todas las combinaciones mostraron un patrón monomórfico, una o dos bandas de igual tamaño para todas las especies utilizando una combinación de iniciadores. Las regiones codificantes del ADN ribosomal se encuentran altamente conservadas, por lo cual se recomiendan cuando se pretende realizar estudios taxonómicos o sistemáticos en un nivel superior de clasificación (Hoy 1994; Navajas et al. 1992).
Con una de las combinaciones y el amplificado digerido con enzimas de restricción (NS5-NS6+DraI), se logró discriminar todas las especies utilizando sólo los fragmentos específicos, por lo cual, este ensayo es útil para arrojar un diagnóstico molecular en la zona de muestreo. Dado lo anterior, se construyó una clave molecular para la determinación de las especies del estudio. El ensayo NS5-NS6+DraI mostró en total 10 bandas para todas las especies (tres bandas en promedio por especie). De las 10 bandas, sólo una se presentó en todas las especies (500 pb), cinco fueron únicas por especie y las cuatro restantes fueron variablemente compartidas por varias especies. Estos resultados muestran el poder discriminante de este ensayo y la utilidad para reconocer las especies estudiadas.
Clave molecular para la determinación de 11 especies de crisópidos (Neuroptera: Chrysopidae), presentes en la zona de muestreo. Información a partir de fragmentos digeridos con la enzima DraI procedentes del ADN ribosomal amplificado utilizando iniciadores NS5-NS6.

Se obtuvieron en total 57 bandas, en un rango de tamaños entre 100 y 800 pb. De éstas, 47 fueron polimórficas y 25 fueron únicas. Ninguna especie presentó gran cantidad de bandas para todos los ensayos, lo cual demuestra que el uso de los seis ensayos provee un alto polimorfismo para diferenciar las especies de crisópidos evaluadas. En general, se obtuvo un alto poder discriminante, a excepción de uno (NS3-NS4+HinfI) con el que se logró que sólo el 83,6 % de las combinaciones posibles de especies se diferenciaran (el porcentaje se calculó de acuerdo con el número de combinaciones de especies posibles [(n-1) (n)/2=55]). Para probar que el patrón fuera reproducible, se varió la cantidad de amplificado digerido (se redujo a la mitad) utilizando la misma concentración de enzima; los resultados coincidieron con los obtenidos inicialmente.
Análisis morfológico
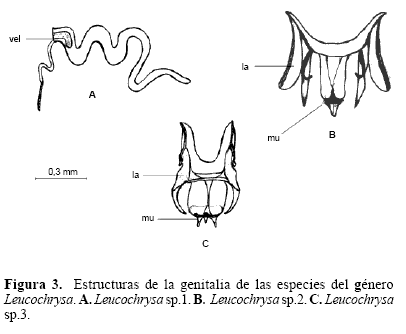
Extracción e ilustración de la genitalia interna de machos y hembras. El método de extracción permitió diferenciar las estructuras que hacen parte de la genitalia interna de las especies. Para el género Ceraeochrysa se observaron estructuras de la genitalia interna como el gonarcus (gc) y la gonapsis (gps). Para el género Chrysoperla se observaron estructuras como el gonarcus (gc), y el tignum (tg), y para el género Leuchochrysa la estructura presente es el gonarcus (gc). La forma de estas estructuras permite una caracterización y diferenciación de cada una de las especies.
La genitalia de machos ofrece una mayor diferenciación de especies que la de hembras. Los géneros del estudio pueden ser fácilmente separados de acuerdo con la presencia de la gonapsis, que distingue al género Ceraeochrysa y de acuerdo a la presencia del tignum, que distingue al género Chrysoperla. En el género Leucochrysa no se observó ni gonapsis ni tignum. En el género Ceraeochrysa, el gonarcus es bastante variable y permite distinguir cada una de las especies. De las estructuras presentes en el gonarcus, dos son las más útiles para separar las especies, la gonocornua y el entoprocessus, de acuerdo con su forma, la primera permite distinguir a Ceraeochrysa sp. 1 de Ceraeochrysa cubana, y el segundo permite distinguir a Ceraeochrysa sp. 2, Ceraeochrysa sp. 3 y C. claveri.
En el caso del tignum, aunque presenta un patrón muy parecido entre las especies del género Chrysoperla, con un análisis cuidadoso de la estructura, es útil para diferenciar las especies. Al caracterizar el tignum es fácil separar a Chrysoperla sp. 1 de C. carnea y C. rufilabris, y estas dos especies pueden ser distinguidas al caracterizar el gonarcus, haciendo especial énfasis en los brazos laterales y el entoprocessus. Con la información generada a partir de las estructuras de la genitalia se construyó una clave dicotómica de determinación específica dentro de los tres géneros.
Clave morfológica para la determinación de 11 especies de crisópidos (Neuroptera: Chrysopidae), en la zona de muestreo

Al observar las estructuras de los géneros Ceraeochrysa y Leucochrysa, es evidente la variación, en comparación con la variación de los caracteres de morfología externa usados normalmente, para determinar especies. Aunque en ocasiones se registró variación intraespecífica en las estructuras, debido seguramente a diferentes factores tales como la edad o el tipo de alimentación, en general se conservó un patrón básico que permitió determinar un espécimen dentro de un grupo dado. De los tres morfotipos de Ceraeochrysa, según la morfología de genitalia, Ceraeochrysa sp. 1 muestra el mismo patrón que Ceraeochrysa cincta (Schneider, 1851) y al revisar la morfología externa se presenta la misma descripción, con la variación del número de manchas en el escapo. Para el caso de Ceraeochrysa sp. 3, la morfología de genitalia interna es bastante similar a la descrita para Ceraeochrysa caligata (Banks, 1946), pero no se realizó una revisión de caracteres de morfología externa que corroboren la determinación de la especie.
Diferenciación morfológica y molecular. Se observó una relación más estrecha entre los géneros Chrysoperla y Leucochrysa debido a que las estructuras como entoprocessus y mediuncus son más frecuentes en los primeros géneros. Es importante exponer que en la literatura existe cierta confusión en cuanto a la identificación o a la designación del nombre de una estructura, específicamente refiriéndose a estas dos. Brooks y Barnard (1990) no consideraron la existencia mediuncus en el género Chrysoperla, mientras Henry et al. (1993) lo identificaron y describieron para tres especies de este género. De Freitas (2003) describió el arcessus para otras tres especies del género, pero no se refirió al mediuncus. Adams (1977) se refiere a estas dos estructuras como sinónimas (mediuncus = arcessus). Brooks y Barnard (1990) no describieron la existencia de mediuncus en el género Leucochrysa, mientras que Tauber (2004) reporta el mediuncus para este género, mas nó el arcessus. De acuerdo con esta situación, para el análisis se decidió dejar la descripción hecha por Henry et al. (1993) y trabajar con el mediuncus para el género Chrysoperla, dada la descripción que hacen de la estructura; en el caso de Leucochrysa se decidió dejar la designación expuesta por Tauber (2004) de mediuncus para este género.(Fig. 1)(Fig. 2)(Fig. 3)
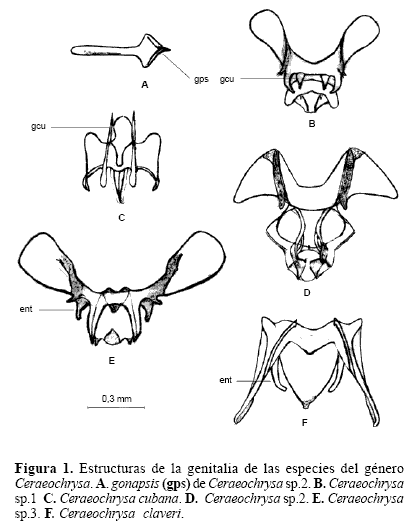
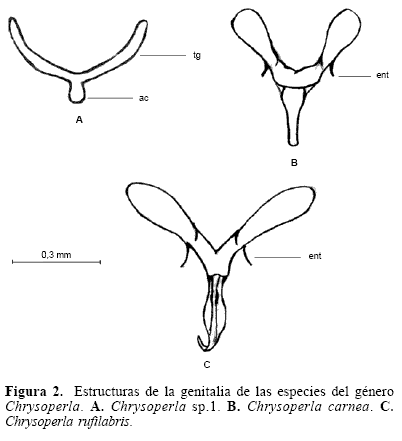
Las estructuras de genitalia para las especies del género Ceraeochrysa muestran mayor diferenciación interespecífica, que la que se observó en las especies de los otros géneros. Esto se explica ya que en el género Ceraeochrysa existe una mayor variación de formas, dimensiones y presencia o visualización de estructuras. Para el género Chrysoperla, las especies Chrysoperla sp. 1 y Chrysoperla rufilabris mostraron estructuras de genitalia bastante similares, mientras que la genitalia de C. carnea está más diferenciada. En general este género presentó poca variación interespecífica, en contraste con las especies de los otros dos géneros. Es necesario revisar con detenimiento los caracteres de genitalia interna al momento de examinar especimenes, a partir de los cuales se puede elucidar más fácilmente la determinación de una especie.
De los tres géneros evaluados, Leucochrysa presentó la menor variación intraespecífica en el análisis molecular, conservándose las bandas específicas de especie y mostrando menos ocurrencia de bandas no específicas (Fig. 4). Este resultado puede ser de gran utilidad, ya que el género ha sido reportado con problemas significativos en su sistemática. Tantos los machos como las hembras exhiben una variación geográfica substancial tanto en el color como en la morfología (Adams 1977). Además, muchos especímenes están esparcidos en las colecciones de la mayoría de los museos y muchas especies recientemente nombradas fueron descritas a partir especímenes únicos. De acuerdo con esto, el género presenta numerosas sinonimias e identificaciones erróneas, que confunden su sistemática notablemente (Tauber 2004).

El género Chrysoperla, al igual que Leucochrysa, presentó baja variación intraespecífica en relación con las bandas no específicas, que no son informativas en el análisis. Esto pudo haber sido influenciado por el hecho de que todos los individuos de dos especies proceden de la cría de laboratorio, encontrándose una fuerte endogamia que puede mostrar un bajo polimorfismo entre las muestras. Sin embargo, para la Chrysoperla sp. 1, aunque se realizó cría, los especímenes estudiados se obtuvieron de la descendencia de varios padres recolectados en el campo, estando poco tiempo sometida a los efectos de endogamia de una cría artificial. La diferenciación morfológica y molecular sirve de diagnóstico en este grupo, en donde se ratifica la baja variación de caracteres morfológicos externos, pero se encuentran nuevos elementos de discriminación entre las especies.
La diferenciación molecular puede ser bastante útil, teniendo en cuenta, que aunque estas especies presentan una baja diferenciación morfológica, se logran separar por técnicas moleculares, que se pueden aplicar sobre un mayor número de especies de este género, y utilizar una muestra que represente poblaciones naturales. Además, teniendo en cuenta que se seleccionaron fragmentos específicos, bien definidos, que se presentaron en todos los individuos de cada especie, no se registró la variación intraespecífica entre las poblaciones.
Todos los resultados en el presente estudio son de suma importancia debido a la poca información que se tiene sobre especies de la familia Chrysopidae para la zona. De 11 especies del estudio, sólo cuatro han sido determinadas hasta especie, y las siete restantes, algunas de las cuales han sido evaluadas por especialistas, permanecen sin determinar.
Se probó la validez de los caracteres de morfología externa usados para separar las especies. Sin embargo, los caracteres de morfología externa pueden no ser siempre ventajosos. Casos como el ya citado de Ceraeochrysa sp. 1, o el de Chrysoperla sp. 1 en la cual no se pudo determinar con seguridad la especie (posiblemente Chrysoperla externa) dadas las variaciones en la coloración del adulto, muestran que en los casos en los que se presente confusión, la información de morfología externa debe ser apoyada por otro tipo de información en este caso la de genitalia interna y la molecular.
En forma general, se puede decir que los caracteres utilizados para distinguir las especies del estudio son útiles, teniendo en cuenta que los datos de morfología de genitalia interna y los datos moleculares mostraron la diferenciación de los once grupos, en el caso de la genitalia interna, con diferentes patrones de las estructuras y en el caso de los datos moleculares, con la utilización de solo una combinación de iniciadores NS y la digestión del amplificado producido con una enzima de restricción (NS5-NS6+DraI). Con la información generada se contribuye a enriquecer el conocimiento taxonómico que sobre la familia se tiene, además de ayudar a resolver los problemas sistemáticos dentro de los géneros.
Agradecimientos
Los autores agradecen a la Doctora Marta Martínez Wells de la Universidad de Connecticut y al Doctor John Ewer de la Universidad de Cornell por toda la documentación suministrada, y al Centro de Investigación de la Caña de Azúcar de Colombia CENICAÑA por la financiación de este trabajo.
Literatura Citada
A'HARA, S.; HARLING, R.; MCKINLAY, R.; TOPPING, C. 1998. RAPD profiling of spider (Aranae) DNA. The Journal of Arachnology 26: 397-400. [ Links ]
ADAMS, P. A. 1977. Taxonomy of United States Leucochyrsa (Neuroptera: Chrysopidae). Psyche 84: 92-102. [ Links ]
ADAMS, P. A.; PENNY, N. D. 1987. Neuroptera of the Amazon basin. Parte 11a. Introduction and Chrysopini. Acta Amazonica 15 (1985): 413-479. [ Links ]
ALBUQUERQUE, G. S.; TAUBER, C. A.; TAUBER, M. J. 2001. Chrysoperla exterma and Ceraeochrysa spp.: potential for biological control in the New World tropics and subtropics. Cap. 21, pp. 408-423. En: McEwen, P.; New, T.; Whittington, A. (eds.). Lacewings in the crop environment. Cambridge University Press, USA. 564 p. [ Links ]
ARNOLD, M. L.; BUCKNER, C. M.; ROBINSON, J. L. 1991. Pollen-mediated introgression and hybrid separation in Luisiana irises. Proceedings of the National Academy of Science of America 88: 1398-1402. [ Links ]
BROOKS, S. J.; BARNARD, P. C. 1990. The green lacewings of the world: a generic review (Neuroptera: Chrysopidae). Bulletin British Museum National History (Entomology) 59: 117-286. [ Links ]
CALDEIRA, R.; CARVALHO, O.; MENDONCA, C.; GRAEFFTEIXEIRA, C.; SILVA, M.; BEN, R.; MAURER, R.; LIMA, W.; LENZI, H. 2003. Molecular differentiation of Angiostrongylus costaricencis, A. cantonensis, and A. vasorum by Polymerase Chain Reaction- Restriction Fragment Length Polymorphism. Memórias do Instituto Oswaldo Cruz 98 (8): 1039-1043. [ Links ]
CHEUNG, W.Y.; HUBERT, N.; LANDRY, B. S. 1993. A simple and rapid DNA extraction method for plant, animal, and insects suitable for RAPD and other PCR analysis. PCR Methods and Applications 3: 69-70. [ Links ]
DE ALMEIDA, R. P.; STOUTHAMER, R. 2003. Molecular identification of Trichogramma cacoeciae Marchal (Hymenoptera: Trichogrammatidae): A new record for Peru. Neotropical Entomology 32: 269-272. [ Links ]
DE FREITAS, S. 2003. Chrysoperla Steinmann, 1964 (Neuroptera: Chrysopidae): Descrição de uma nova espécie do Brasil. Revista Brasileira de Entomologia 47 (3): 385-387. [ Links ]
HENRY, C. S. 1983. Acoustic recognition of sibling species within the holartic lacewing Chrysoperla carnea (Neuroptera: Chrysopidae). Systematic Entomology 8: 293-301. [ Links ]
HENRY, C. S.; WELLS, M. M.; PUPEDIS, R. J. 1993. Hidden taxonomic diversity within Chrysoperla plorabunda (Neuroptera: Chrysopidae): two new species based on courtship songs. Annals of the Entomological Society of America 86 (1): 1-13. [ Links ]
HOY, M. A. 1994. Insect molecular genetics. An introduction to principles and applications. Academic Press, Inc. San Diego, California. 546 p. [ Links ]
KANG-ZHEN, D.; WEN-ZHU, L.; JUN-ZHI, C.; XING-KE, Y. 2004. Three new species of Dichochrysa (Insecta: Neuroptera: Chrysopidae) from China with a checklist of Chinese Dichochrysa. The Raffles Bulletin of Zoology 52 (1): 67-74. [ Links ]
LANDRY, B. S.; DEXTRAZE, L.; BOIVIN, G. 1993. Ramdom amplified polymorphic DNA markers for DNA fingerprinting and genetic variability assessment of minute parasitic wasp species (Hymenoptera: Mymaridae and Trichogrammatidae) used in biological control programs of phytophagous insects. Genome 36: 580-587. [ Links ]
LUCHETTI, A.; MANTOVANI, B.; TRENTINI, M. 2005. Rapid identification of non-neosomic Tunga penetrans and Tunga trimamillata (Insecta: Siphonaptera) specimens through PCRRFLP method. Bulletin of Insectology 58 (1): 15-18. [ Links ]
NAVAJAS, M.; COTTON, D.; KREITER, S.; GUTIÉRREZ, J. 1992. Molecular approach in spider mites (Acari: Tetranychidae): preliminary data on ribosomal DNA sequences. Experimental and Applied Acarology 15: 211-218. [ Links ]
NUÑEZ, E. 1988. Chrysopidae (Neuroptera) del Perú y sus especies más comunes. Revista Peruana de Entomología 31: 69-75. [ Links ]
PENNY, N. D. 2002. A guide to the lacewings (Neuroptera) of Costa Rica. Proceedings of the California Academy of Sciences 53 (12): 161-457. [ Links ]
RAMÍREZ, D. 2002. Reconocimiento y evaluación del uso de especies de la familia Chrysopidae para el manejo del pulgón amarillo Sipha flava (Homoptera: Aphididae) en caña de azúcar. Tesis de pregrado. Cali-Colombia. Universidad Nacional. Palmira, Facultad de Ciencias Agropecuarias. 137 p. [ Links ]
SAMBROOK, J.; FRITSCH, C. R.; MANIATIS, T. 1989. Preparation and examination of agarose gels, pp. 6.9-6.19. En: Cold Spring Harbor Laboratory Press, N.Y. 2da Ed. Molecular cloning: A laboratory manual. [ Links ]
SCHILTHUIZEN, M. 2003. Shape matters: the evolution of insect Genitalia. Proceedings of the Section Applied and Experimental Entomology of the Netherlands Entomological Society 14: 9-15. SHARMA, A. D.; GILL, P. K.; SINGH, P. 2002. DNA isolation from dry and fresh samples of polysaccharide-rich plants. Plant Molecular Biology Reporter 20: 415a-415f. [ Links ] [ Links ]
SILVA, I.; HONDA, J.; VAN KAN, F.; HU, J.; NETO, L.; PINTUREAU, B.; STOUTHAMER, R. 1999. Molecular differentiation of five Trichogramma species occurring in Portugal. Biological Control 16: 177-184. [ Links ]
TAUBER, C. A. 2004. A systematic review of the genus Leucochrysa (Neuroptera: Chrysopidae) in the United States. Annals of the Entomological Society of America 97 (6): 1129-1158. [ Links ]
TAUBER, C. A.; DE LEÓN, T.; PENNY, N. D.; TAUBER, M. J. 2000. The genus Ceraeochrysa (Neuroptera: Chrysopidae) of America North of Mexico: larvae, adults, and comparative biology. Annals of the Entomological Society of America 93 (6): 1195-1221. [ Links ]
VIDIGAL, T.; MONTRESOR, L.; SIMPSON, A.; CARVALHO, O. 2002. Polymerase Chain Reaction and Restriction Fragment Length Polymorphism of Cytocrome Oxidase Subunit I Used for differentiation of Brazilian Biomphalaria species intermediate host of Schistosoma mansoni. Memórias do Instituto Oswaldo Cruz 97 (1): 47-52. [ Links ]
WILKERSON, R. C.; GAFFIGAN, T. V.; LIMA, J. 1995. Diagnosis by random amplified polymorphic DNA polymerase chain reaction of four cryptic species related to Anopheles (Nyssorhynchus) albitarsis (Diptera: Culicidae) from Paraguay, Argentina and Brazil. Memórias do Instituto Oswaldo Cruz 90 (6): 1-12. [ Links ]
Recibido: 18-ene-2007 - Aceptado: 3-nov-2007

