










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488On-line version ISSN 2665-4385
Rev. Colomb. Entomol. vol.34 no.2 Bogotá July/Dec. 2008
Artículo de Revisión
Diversification practices: their effect on pest regulation and production
Prácticas de diversificación: sus efectos en la regulación de plagas y en producción
KATJA POVEDA1, MARÍA ISABEL GÓMEZ2 and ELIANA MARTÍNEZ3
1 Postdoctoral researcher. Ph.D. Agroecology, Göttingen University, Waldweg 26, 37073 Göttingen, Alemania. kpoveda@gwdg.de. Autor para correspondencia.
2 Researcher. MSc. Universidad Nacional de Colombia. Facultad de Agronomia. Ciudad Universitaria. Bogotá, Colombia. migomezj@unal.edu.co.
3 Researcher. MSc. Universidad Nacional de Colombia. Departamento de Biología. Ciudad Universitaria. Bogotá, Colombia. emartinezpa@unal.edu.co.
Abstract: The interest to shift pest management strategies from the intensive use of agrochemicals to more sustainable and ecologically friendly practices has increased in recent years. One alternative to conventional farming systems is the implementation of diversification practices that increase diversity in- and around- the field to increase the incidence of natural enemies, reduce pest pressure and enhance crop production. In this review we illustrate the theoretical framework on which diversification practices are based and contrast it with the empirical evidence. The detailed review of 62 original studies published in the last ten years, shows that diversification practices (a) enhance natural enemies in 52%, (b) reduce pest pressure in 53% and (c) increase yield in only 32% of the cases where this was examined. We discuss these results on the basis of the reviewed studies providing key elements that should be taken into account to design diversification practices that can be implemented as competitive pest management strategies that cover the farmers’ needs, reducing the intensive use of agrochemicals.
Key words: Crop yield. Intercrop. Flowering-plant. Repellent-plant. Trap- plant.
Resumen: El interés por dirigir las estrategias de manejo de plagas desde el uso intensivo de agroquímicos a prácticas sostenibles y ecológicamente amigables se ha incrementado en los últimos años. Una alternativa para los sistemas de cultivo convencionales es la diversificación tanto dentro como alrededor de los cultivos buscando incrementar la incidencia de enemigos naturales, reducir la presión de las plagas e incrementar o mantener la producción del cultivo. Se presenta una revisión del marco teórico que ha sido base para el estudio de las prácticas de diversificación y se contrasta con la evidencia empírica. Los resultados reportados en 62 estudios originales publicados en los últimos diez años, muestran que las prácticas de diversificación (a) incrementan los enemigos naturales en el 52% de los casos, (b) reducen la presión de las plagas en un 53% de los estudios e (c) incrementan los rendimientos en solo el 32% de los casos. Se discuten estos resultados teniendo como base los estudios que proveen elementos claves para ser tomados en cuenta para el diseño de prácticas de diversificación que puedan ser implementadas como estrategias competitivas de manejo de plagas y que cubran las necesidades de los productores reduciendo el uso intensivo de agroquímicos.
Palabras clave: Producción. Policultivos. Plantas con flores. Plantas repelentes. Plantas trampa.
Introduction
The use of chemically synthesized fertilizers and pesticides to reduce crop pests and weeds and to increase harvest yields is common in current agricultural practices. These practices are coupled with the removal of weeds from within and around crops, large field sizes, tillage operations of varying intensity and the degradation or destruction of non-crop habitats (reviewed by Gurr et al. 2003). Although these practices have substantially increased yield, they also increased production costs, pesticide resistance and have affected ecosystem and human health (Matson et al. 1997; Krebs et al. 1999; Tilman et al. 2002). At the ecosystem level they caused serious ecological problems such as water contamination, habitat degradation and loss of biodiversity (Matson et al. 1997 ; Krebs et al. 1999; Staver et al. 2001; Tilman et al. 2002) with the concomitant loss of ecological functions such as pollination and biological control (Kruess and Tscharntke 1994; Matthies and Schmid-Hempel 1995; Didham et al. 1996; Kruess and Tscharntke 2000; Tilman et al. 2002). In response to these negative effects, the world market has increased its demand for residue free food (Thompson 1998; Magnusson and Cranfield 2005). One alternative to conventional farming practices is the increase of in-and around- crop diversity to reduce pest pressure. It has been generally assumed that this practice stimulates the presence of natural enemies and enhances pest suppression, potentially reducing the need for costly and ecologically disruptive insecticide applications (i.e. Altieri and Nicholls 1994; Gurr et al. 2004). However, in order to propose technological packages that can be implemented by the farmers, the link between diversification practices and increased crop yield must be successfully shown (Gurr and Wratten 1999). There is an extensive theoretical literature predicting that biodiversity could enhance natural enemies and increase pest suppression (see next section). Also, empirical studies have tested the relationship between species diversity and functioning of natural enemy assemblages and pest suppression (Cardinale et al. 2003; Wilby and Thomas 2002a; Wilby and Thomas 2002b; Finke and Denno 2004; Straub et al. 2008), but a convincing link between habitat diversification, pest suppression and crop production seems to be missing.
Our goal is to contrast the theoretical and empirical evidence on how diversification practices affect natural enemies, pest pressure and crop yield. We start this review by summarizing the theoretical background, then we review original literature to determine if the theoretical expectations are met in the empirical work, emphasizing studies done on crop yield. Based on the results of this review we discuss the possible causes of any discrepancy between theory and observation, and propose some guidelines for future studies to develop management practices that meet the needs of farmers and reduce the use of agrochemicals.
How does diversity increase pest control and production? Theoretical background
There have been several hypotheses to explain how vegetation diversity can directly affect crop pests. In general, vegetation diversity has been proposed to disrupt the pest’s ability to locate the host plant, to increase mortality of the pest or to repel the pest. Here we give a brief overview of the hypotheses that have been proposed until now:
a. The disruptive crop hypothesis is equivalent to Root’s (1973) resource concentration hypothesis and stipulates that herbivores in polycultures will have more difficulties finding crop plants associated with one or more taxonomically or genetically different plants than finding crop plants in monoculture (Vandermeer 1989).
b. The trap crop hypothesis suggests that pests will be attracted to associated plants and hence are less likely to leave the trap crop and wander into the principal crop (Vandermeer 1989).
c. The natural enemy hypothesis proposes that a lower number of phytophagous insects are found in complex environments because predators and parasitoids are more diverse and abundant in those environments compared to simple environments (Root 1973; Russell 1989).
d. The barrier crop hypothesis or physical obstruction hypothesis bases its effectiveness on the use of taller non-host plants to obstruct the movement of the pest insect within the cropping system (Perrin and Phillips 1978).
e. The visual camouflage hypothesis also known as the “apparency hypothesis” incorporates the visual stimuli that induce herbivores to land on plants: color and plant height. Herbivores tend to land on tall green plants, so that using noncrop plants to make the crop “less apparent” by adding more green or taller plants is a useful mechanism to camouflage the crop (reviewed by Finch and Collier 2000).
f. The associational resistance hypothesis proposes that nonhost plants confer protection to the crop by releasing “odor masking” substances into the air making the crop “invisible” to the herbivore (Tahvanainen and Root 1972).
g. The repellent chemicals hypothesis predicts that the nonhost plants emit odors that repel the herbivore (Uvah and Coaker 1984).
h. The altered profile of the host plant odor hypothesis bases its effect on changes in the physiology of the plant through certain chemicals they take up from the soil (reviewed by Finch and Collier 2000).
The above hypotheses are supported in most cases by experimental evidence (reviewed by Finch and Collier 2000). However, the application of these techniques would be useless for agriculture if pest suppression and enhanced natural enemies do not translate into increased yield. Studies showing the link between pest suppression and yield are limited (Ostman et al. 2003; Cardinale et al. 2003) but the results are promising. Ostman et al. (2003) showed that ground-living natural enemies (ground beetles, Carabidae; rove beetles, Staphylinidae and spiders) of the bird cherry-oat aphid Rhopalosiphum padi L., 1758 dramatically reduce aphid abundance. Aphid suppression led to a 23% increase in barley Hordeum vulgare L. (Poaceae) yield compared to scenarios where natural enemies of the bird cherry-oat aphid were absent. In another study performed by Cardinale et al. (2003) on the effect of three natural enemies (Harmonia axyridis Pallas, 1773, Coccinellidae; Nabis sp., Nabidae and Aphidius ervi Haliday, 1834 Braconidae) on the pea aphid Acyrthosiphon pisum Harris, 1776 (Aphididae) that feeds on alfalfa Medicago sativa L. (Fabaceae) they found an indirect effect of natural enemies on production mediated by herbivore suppression. The presence of all three enemy species reduced pea aphid density in the field. Crop yield was inversely related to pea aphid density and therefore the presence of natural enemies should increase yield. Although previous studies seem to be very promising, we have to take into account that those studies actively manipulate the presence of natural enemies in the field (Cardinale et al. 2003; Ostman et al. 2003), not reflecting what would happen in an agricultural setting. Thus the question remains open if diversification practices actually do increase the presence of natural enemies and increase pest suppression as would be predicted from the above hypotheses.
Effect of diversification on natural enemies, herbivores and crop damage and production
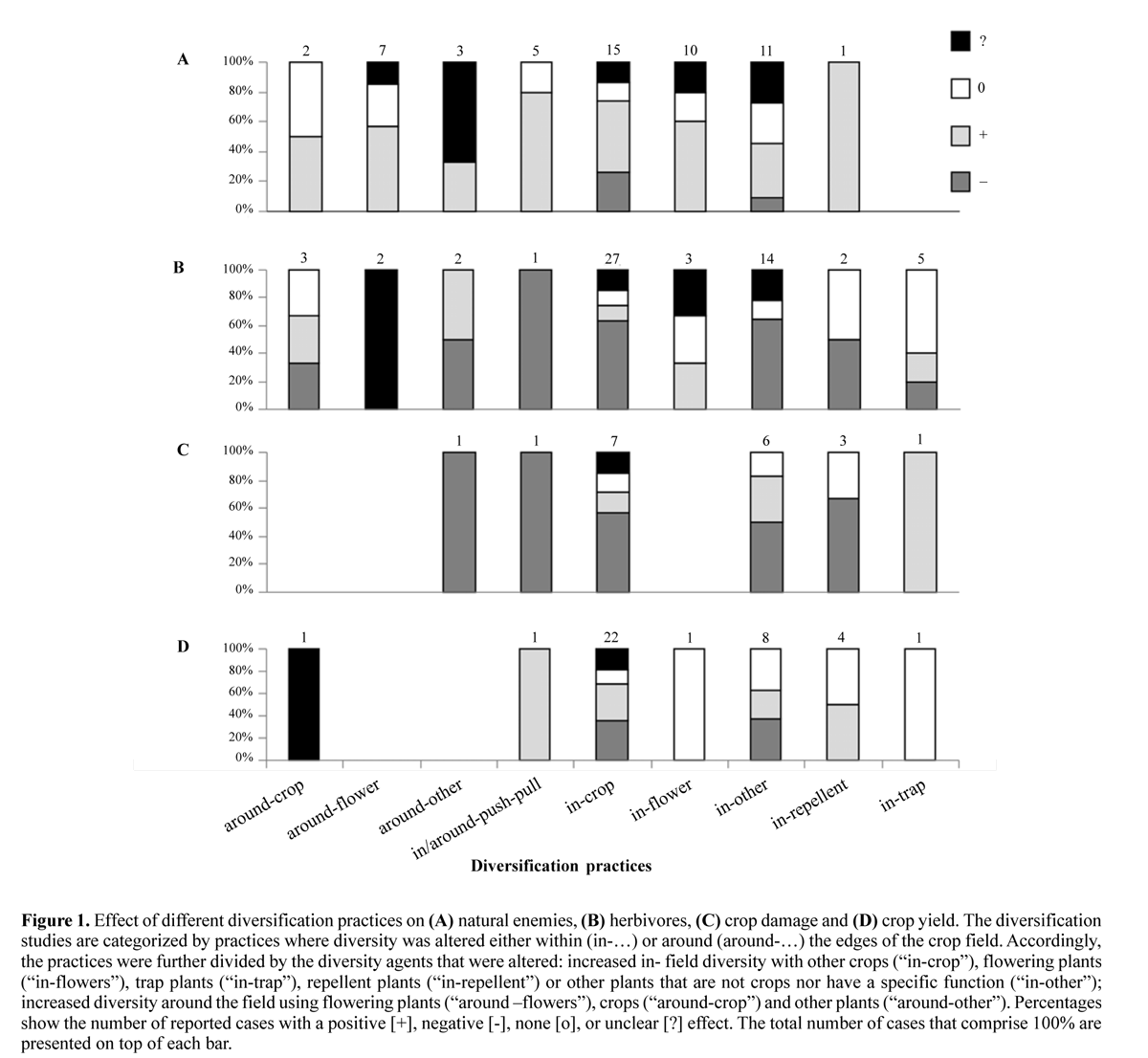
Theory predicts that diversified crops in and around the field should have a higher and more effective population of natural enemies, decreased pest pressure on the crop and consequently higher yields in comparison to monoculture. In order to test this prediction we searched for articles published in scientific journals in the last ten years that investigated the effect of diversification practices, like intercropping and local habitat manipulation, on pest suppression and biological control. To avoid biasing the articles with respect to known authors, groups or papers, we searched the literature database (ISI Web of Knowledge: http://isiknowledge.com) using the keywords: “pest* AND diversification”, “pest* AND intercrop*”, “pest* AND habitat manipulation”, “habitat manipulation AND agroecosystems”, “biological control AND agroecosystems”, “biological control AND habitat manipulation”. Out of the 279 references obtained in our search, we used the following criteria to finally select the 62 references included in our analysis (Table 1): (1) studies should be conducted at a local scale, including diversification practices in and immediately around the crop, (2) the timing of crop growth and diversification practices should be the same, excluding practices like crop rotation, (3) only studies performed in the field and on crops or their associated organisms are included, and (4) only studies that were available to us through the online libraries of the University of Göttingen (Germany) and Cornell University (USA) were included. For each study we recorded the crop, the diversification mechanism used, the effects (positive, negative and/or neutral) reported on herbivores, natural enemies, crop damage and crop production. Diversification practices were categorized into techniques performed in (52 studies) and around (seven studies) the crop and these categories were further subdivided by the type of plant that was used to increase diversity. Twenty-three studies increased within field diversity with other crops (“in-crop”), seven studies used flowering plants in the field (“in-flowers”) to attract natural enemies and four studies used flowering plants around the crop (“around-flowers”). The rest of the studies increased in-field or around-field diversity by specific functional groups like trap- (“in-trap”- five studies) or repellent- plants (“in-repellent”- four studies) to attract or repel herbivores, by crops around the field (“around-crop”- two studies) or non-specifically by using weeds, ground cover plants, or natural diversity (“in-other”- 13 studies, and “around-other”- one study) (Table 1). Only three studies used combined in- and around-field practices and push-pull (“in/around-push-pull”) strategies to simultaneously attract herbivores to trap plants around the field and repel herbivores from the center of the crop (studies 34, 39, 40 in (Table 1)). In order to quantify if diversification practices decreased, increased or had no effect on natural enemies, herbivores and production, we independently scored each of the effects reported in each study. In those cases where more than one effect was shown, as for example in ), who reported different effects on different species of natural enemies, we scored each reported effect independently. Thus, we had 62 articles that report 171 effect cases. Studies reporting contradictory effects on the same species in different locations or in different years or on different ways of measuring the same response, were quantified as an unclear response. For example, Bukovinszky et al. (2004) reported a positive effect of intercropping on the number of Plutella xylostella per broccoli plant but a negative effect of the same treatment on the abundance of P. xylostella at a plot level, so the effect on the herbivore was scored as unclear.
From the 62 studies only nine (studies 32-34, 41, 44, 47, 50, 54 & 60) actually report positive effects of diversification practices on yield coupled with enhanced presence of natural enemies and / or a reduction in pest pressure. Eight studies showed that diversification practices can cause a reduction in yield as a consequence of both positive as well as negative effects on herbivores, crop damage and / or natural enemies (studies 8, 13, 26, 31, 43, 51, 55 & 62). Most of the studies, however, just reported effects on natural enemies, pest presence or yield and there is high variation in the effect of diversification on each of these response variables. Diversification effects on natural enemy populations were recorded in 35 of the 62 studies. Natural enemies were quantified in terms of abundance or (activity) density of parasitoids (study 18), predators (studies 10, 21, 23-25, 27, 28, 36, 37, 41, 42, 44, 46, 48 & 49), or natural enemies in general (studies 11, 14, 17, 19 & 20), species richness or diversity of predators (study 37), parasitism rates (studies 3, 4, 6, 8, 12, 16, 23, 26, 34, 47 & 50) or predation rates (studies 3 & 36). In those studies where natural enemies were investigated, 52% of the cases reported a positive effect of diversification practices. The diversification practices that frequently led to an enhancement of natural enemies were increasing abundance of flowering plants in the crop (studies 3, 6, 19, 29 & 30), enhancing flowering plants around the crop (studies 15, 48), intercropping mechanisms (studies 12, 14, 21, 53, 54, 60, 62), increasing in-field plant diversity nonspecifically (25, 26, 46, 55), increasing in-field diversity with repellent plants (52), and push-pull strategies (studies 39 & 40) (Fig. 1A). In 20% of the cases there was no effect of diversification on natural enemies and only 9.5% of the cases reported a negative effect of diversification. Unclear effects were reported in 18.5% of the cases (Table 1).
In 44 out of the 62 articles the effect of diversification practices on crop herbivores was quantified. Herbivores were quantified in terms of larval infestation (study 2), number of eggs (studies 7, 12 & 59), density or abundance of immatures and adults (studies 5, 8, 9, 14, 16-20, 22, 23, 25-29, 35-37, 41- 43, 45, 47, 50-53, 55-58, 60 & 62), species richness (study 11) and oviposition preferences (studies 39 & 44). Overall, 53% of the reported cases showed a negative effect of diversification on herbivores as would be expected by theory. The diversification practices that seem to be most effective in leading to a herbivore reduction were intercropping (studies 8, 17, 21, 41, 42, 47, 50, 51, 53, 60 & 62 reporting the expected effects), non-specific in-crop diversity increase (studies 7, 16, 22, 26, 27, 44 & 55 reporting the expected effects) and pushpull strategies (studies 39 & 40) (Fig. 1B).
Diversification practices had a positive effect on herbivore presence in 11.9% of the analyzed cases, no effect in 22% of the cases and an unclear effect in 13.1% of the cases (Table 1). Effects on plant damage were reported in 18 studies and were quantified in terms of foliage consumption (studies 8, 50 & 51), deposits of frass and tunneling (study 28), tissue damage (studies 9, 16, 35, 38, 54 & 62), stem boring (studies 12, 23, 33, 34, 44 & 62), root knotting (study 31) or root necrosis (study 38). In 57.9% of the reported cases, plant damage was reduced with diversification practices. This expected effect was achieved when implementing diversification practices like intercropping (studies 8, 13, 51, 54 & 62), in-field use of repellent plants (studies 32 & 33), push-pull practices (study 34) and the use of non-specific plants in (studies 26, 28 & 44) and around the crop (study 23) (Fig. 1C). Diversification practices increased crop damage in 21.1% of the reported cases. No effect of diversification practices on crop damage occurred in 15.8% of the cases, while only 5.2% reported an unclear effect (Table 1).
Effects on production were quantified in 30 of the 62 studies. Production was quantified in terms of yield (studies 1, 9, 13, 19, 22, 24, 26, 32-35, 38, 41, 43-45, 47, 49, 50-56, 60 & 62), size of the product (studies 8 & 27) and development time (study 31). There was a positive effect of diversification practices in 32% of the cases. Out of all the diversification mechanisms, the push-pull strategy reported a consistently positive effect on production, however, this strategy was represented by only one study evaluating effects on production (study 34) (Fig. 1D). Diversification practices had a negative effect on production in 28.9% of the cases, no effects in 26.1% of the cases and an unclear effect in 13% of the cases.
Discussion
We did not find that diversification practices consistently enhance natural enemies, decrease herbivores, or increase production. Rather, for natural enemies and herbivores, only about half of the cases report the expected effects. Of even more concern, for practicing farmers, only one third of the cases report an increase in production. Given the somewhat discouraging results we discuss the possible causes that lead to these unexpected effects. Using studies that show positive results as examples, we explore how particular approaches could help future diversification studies achieve the expected goals that will result in farmer adoption of these kinds of technologies.
The importance of the “right kind” of diversity. Although diversification practices base their effectiveness on the fact that high diversity should lead to pest suppression, it is also known that high plant diversity in agroecosystems does not automatically reduce pest pressure and enhance the activity of natural enemies (Landis et al. 2000; Heemsbergen et al. 2004). Several authors have noted that to selectively enhance natural enemies, the functionally important elements of diversity should be identified and provided, rather than encouraging diversity per se (Landis et al. 2000). Heemsbergen et al. (2004) suggest that it is not the species number but the degree of functional differences between species that enhance overall ecological functions. The species-specific contribution to the range of functional groups in a community might be an important mechanism by which biodiversity generates positive interactions that enhance ecological services like pest suppression. Therefore, the screening of key plants is of crucial importance to shape agricultural systems to specifically reduce pest pressure and enhance production.
Increasing diversity with other crops and plants. The use of other crops to reduce pest pressure and increase yield of the main crop, known as intercropping, is a long established practice (Vandermeer 1989; Altieri and Nicholls 1994). The effectiveness of this practice is exemplified in one of the reviewed studies where cowpea Vigna unguiculata (L.) Walp. (Fabaceae) and okra Abelmoschus esculentus (L.) Moench (Malvaceae) were intercropped with tomato Solanum lycopersicum L. (Solanaceae) (Pitan and Olatunde 2006). Intercropping had a negative effect on the herbivores and a positive effect on yield in both crops, though the exact mechanism remains unclear.
However, increasing diversity can increase pest problems (reviewed by Landis et al. 2000). This undesired effect can be avoided with knowledge of pest natural history. This was certainly shown in the study by Ngeve (2003) where intercropping cassava Manihot esculenta Crantz (Euphorbiaceae) with maize Zea mays L. (Poaceae) and groundnuts Arachis villosulicarpa Hoehne (Fabaceae) actually increased the severity of root mealybug Stictococcus vayssierei Richard (Stictococcidae) infestation. This increased pest pressure was a consequence of using other mealybug host plants as the intercropping species (Ngeve 2003). It becomes obvious from this example that knowledge of the alternative hosts of the pest is crucial, in order not to add additional food resources to a pest that is meant to be controlled. This factor is also important when choosing flowering plants to attract natural enemies, and will be discussed in the next section.
Regardless of the previously published work on how pest suppression leads to an increased yield (Cardinale et al. 2003; Ostman et al. 2003), our literature review demonstrates that diversification practices that reduce pest pressure do not necessarily achieve an increased production (i.e. Showler and Greenberg 2003; Sastawa et al. 2004; Schulthess et al. 2004). Mechanisms like competition and allelopathic effects between plants could be responsible for these effects. Sastawa et al. (2004) compared intercropping systems varying in their complexity: simple intercrops of millet Pennisetum glaucum (L.) R. Br. (Poaceae) and soybean Glycine max (L.) Merr. (Fabaceae), and more complex intercrops of millet, soybean, groundnut and cowpea. They found that the more complex systems actually led to a reduction in the number of the pod sucking bug Nezara viridula Linnaeus, 1758 (Pentatomidae) and a reduction in the defoliation caused by two carabids (Egadroma discriminatum Basi and Siderodactylus sagitarius Meigen) to soybean. However, soybean yield also decreased in the more complex diversification systems. The authors suggest that competition and shading by the intercropped plants were the possible causes for the reduced production (Sastawa et al. 2004). Very similar results are reported by Schulthess et al. (2004) and Showler and Greenberg (2003) where diversification practices suppress the pest but simultaneously reduce yield, probably as a consequence of competition. Moreover, empirical evidence shows that competition not only decreases yield, but could also be the cause of reduced pest pressure. Bukovinszky et al. (2004) assessed the effect of intercropping Brussels sprouts Brassica oleracea var. gemmifera D. C. (Brassicaceae) with malting barley (H. vulgare) on the populations of P. xylostella and Brevicoryne brassicae L., 1758 (Aphididae). They reported a lower incidence of both herbivores on the intercropped Brussels sprout in comparison to monocrops, but the effect seemed to be caused by the effect of competition between both plants. Competition caused drought stress on Brussels sprout plants, leading to reduced size and delayed phenology, which made those plants less apparent and less attractive to the herbivore (Bukovinszky et al. 2004). Effects of plant-plant interactions like competition and allelopathy (Kamunya et al. 2008) can negatively affect production and override positive effects on pest suppression. The previous examples make clear that effects on pest pressure cannot be simply extrapolated to crop yield and that great caution has to be taken when choosing the plant to intercrop.
The goal of diversifying crops is often to increase the availability of appropriate microhabitats for the natural enemies of the pests (Sunderland and Samu 2000; Gurr et al. 2003). Examples from our literature review show that broccoli (Brassica oleracea var. botrytis L. - Brassicaceae) stands intercropped with different kinds of clover (Trifolium fragiferum L., Trifolium repens L., Melilotus officinalis (L.) Lam.) have increased spider density and increased yield in comparison to broccoli monocrops (Hooks and Johnson 2004). Also intercropping maize with groundnut, soybean and Phaseolus beans increases nesting of predatory ants in the field, reducing termite attack and increasing yield (Sekamatte et al. 2003). In both cases the enhanced predator presence is explained by the provision of extra food resources and refuges as proposed by Root (1973), making these desirable characteristics in the plants used to intercrop. However, there is a confounding effect in the last two studies when reporting a yield increase given by the use of legumes as intercrop. Legumes are known for their nitrogen fixing capacity that should increase the nitrogen available to the main crop through organic residues and the residual effect of the biologically fixed nitrogen (Lal et al. 1978). Although in the previous examples it is not clear if the increased yield was accomplished by pest suppression or by the presence of legumes, the desired effect of increased yield was reached. The previous examples show that legumes are excellent candidates for intercropping giving their characteristics of enhancing the presence of natural enemies and at the same time increasing yield. However, factors like competition for resources can also be playing a role when intercropping legumes. In one study Rao and Mathuva (2000) report two different outcomes of intercropping legumes. They showed that intercropping maize with pigeonpea Cajanus cajan (L.) Millsp. (Fabaceae) increased yield by 24% in comparison to monocultured maize, while intercropping maize with the perennial legume Gliricidia sepium (Jacq. Kunth ex Walp.) did not affect maize yield. The difference in the response was attributed to the type of legume. The competition for water between the superficial roots of Gliricidia and maize seem to be the reason that there was no yield increase (Govindarajan et al. 1996; Rao and Mathuva 2000). Negative yield effects as a result of intercropping with a legume are reported by Harvey and Eubanks (2004), who intercropped white clover (T. repens) in broccoli to control P. xylostella with fire ants. Competition lead to smaller, fewer and deformed broccoli leaves and finally to a reduced yield. These latter studies show that although legumes can have the added advantage of increasing yield through their nitrogen fixing capacities, this effect cannot be generalized for all legumes in all crops. Competition between the chosen legume and the crop has to be tested before implementing them in a diversification practice.
Like plants from other groups, legumes can also have an effect on pest oviposition. Bjorkman et al. (2007) showed that the turnip root fly Delia radicum L., 1758 (Anthomyiidae) reduced oviposition by approximately 50% when intercropping cabbage Brassica oleraceae L. (Brassicaceae) with red clover T. pratense. A similar result was reported by Chabi-Olaye et al. (2005a) who showed that intercropping maize with legumes could reduce the percentage of plants with stem borer eggs also by approximately 50%. The incidence of Thrips tabaci Lindeman, 1889 (Thripidae) is also reduced when intercropping leek Allium porrum L. (Liliaceae) with the legume T. fragiferum (den Belder et al. 2000). In neither study was the effect on production reported, thus it remains unclear if the negative effect on herbivore oviposition translates into a positive effect on production. Although none of the studies emphasized the mechanism underlying the herbivore response, the disruption of host finding could be a feasible explanation (Chabi-Olaye et al. 2005a; Bjorkman et al. 2007), and changes in plant quality through intercropping seem also to be playing a role (den Belder et al. 2000).
Flowering plants to enhance natural enemies. Potential mechanisms of positive diversity effects include improving the availability of alternative foods such as nectar, pollen and honeydew for the natural enemies of pests (Patt et al. 1997; Landis et al. 2000; Tylianakis et al. 2004). However, the mere presence of flowering plants in an agroecosystem is not always sufficient to guarantee nectar supply for parasitoids (Baggen and Gurr 1998; Wäckers 2004) and identification of the key flowering plants for certain parasitoids is required to guarantee the enhancement of natural enemies. The first important factor is to determine plant identity. Colley and Luna (2000) studied the effect of 11 different flowering plants on the presence of aphidophagous hoverflies (Syrphidae) giving an example of how a screening process for a flowering plant takes place. However, it is important to take into account that resources that are available for natural enemies could also be a food source for herbivorous pests (Lavandero et al. 2006). For example, Jones and Gillett (2005) intercropped polycultures with sunflowers Helianthus annuus L. (Asteraceae), which increased the presence of arthropod natural enemies (Jones and Gillett 2005) and insectivorous birds (Jones and Sieving 2006), but at the same time herbivorous pests (Jones and Gillett 2005). For this reason screening for suitable flowering plants should also include the screening of the suitability for pest herbivores as was done by Begum et al. (2006). They screened five flowering plants to detect their effect on natural enemies and herbivores.
After greenhouse and field experiments they determined that Lobularia maritima (L.) Desv. (Brassicaceae) provided benefits to the egg parasitoid Trichogramma carverae Oatman and Pinto (Trichogrammatidae) when mass released in vineyards, but not on the leafroller pest Epiphyas postvittana (Walker) (Tortricidae). Another important factor is that field conditions and the type of management can alter the outcome of diversification practices. Although the results from Begum et al. (2006) seem very promising, the applicability to different conditions seems to be inconsistent. Bell et al. (2006) used the same species (L. maritima) in vineyards to control the same type of pest (E. postvittana) but they did not find the same results; plots intercropped with the flowering species did not have increased parasitism rates. In this case biotic factors like proximity to an orchard, which seems to be the source for parasitoids, had a higher effect on parasitism than the increased availability of local resources like L. maritima. This emphasizes screening for the right flowering plant is not sufficient to achieve the expected results, but that results from laboratory settings or given field conditions may not yield the same effects under different conditions.
Using flowering plants around the crop could have the disadvantage that the population of predators and parasitoids stays within the flowering strips around the crop and does not migrate to the field when resources in that strip are more abundant (Rand et al. 2006). This was exemplified by the study of Frere et al. (2007) where rose Rosa rugosa Thunb. (Rosaceae) bushes were used to increase diversity around wheat Triticum aestivum L. (Poaceae) fields. However, the presence of rose bushes did not influence the aphid population within the field. One likely explanation is the relatively higher availability of resources such as pollen, nectar, aphid hosts for predators and parasitoids in the rose borders.
Although reviews and original studies (Baggen and Gurr 1998; Gurr and Wratten 1999; Landis et al. 2000; Wäckers 2004; Lavandero et al. 2006) have already highlighted the importance of selecting the appropriate flowering plant, our literature review reveals that the link between enhancing natural enemies through flowering plants and increasing crop yield is still missing. Only one of the eleven studies in which diversity was increased with flowering plants reported an effect on production. Fitzgerald and Solomon (2004) found no effect on apple Malus domestica Borkh. (Rosaceae) yield when the trees were undersown with flowering plants. However, there is evidence from other studies that flowering plants can reduce yield, probably as a result of competition (Brown and Glenn 1999).
Repellent plants for herbivores. An alternative method to reduce pest pressure is to identify key plants that repel herbivores (Vanhuis 1991; Finch et al. 2003; Lapointe et al. 2003; Morley et al. 2005). In this review only four studies that used repellent plants against herbivores also studied their effects on production. Two out of the four studies successfully achieved the goal of reducing pests, increase yield and even suppress weeds (Khan et al. 2006a; Khan et al. 2006b). One of the studies (Khan et al. 2006b) exemplifies the importance of continuing screening for appropriate plants, to cover the different needs and the heterogeneity found in different regions. Knowing that Desmodium uncinatum (Jacq.) DC. (Fabaceae) had the potential to control the stemborers Chilo partellus (Swinhoe) (Crambidae) and Busseola fusca (Füller) (Noctuidae) on maize and suppress the witchweed Striga hermonthica (Del.) Benth. (Scrophulariaceae), Khan et al. (2006b) continued searching for the effectiveness of four other species of Desmodium to be used under different agroecological conditions. All Desmodium species tested achieved the same results on stemborer suppression, witchweed control, and maize yield increase as D. uncinatum. This result is the basis for a technological tool that does not depend on a single species, increasing the range of sites where the technology can be implemented. Once a promising plant is identified as having a repellent effect, its properties to control herbivores in different crops should be investigated. This was performed by Khan et al. (2006a), who studied the effectiveness of D. uncinatum in sorghum Sorghum bicolor (L.) Moench (Poaceae) fields after demonstrating their effectiveness in maize. They found that with the same repellent plant (D. uncinatum) they could achieve pest reduction, weed control, and increased yield not only in maize but also in sorghum (Khan et al. 2006a), increasing the applicability of a given technology to more than one crop.
Schader et al. (2005) reported that intercropping cotton Gossypium barbadense L.(Malvaceae) with basil Ocimum basilicum L. (Lamiaceae) as a repellent plant reduced pest infestation and increased the abundance of the epigeic fauna. However, no correlation between pest infestation and cotton yield was detected; there was no decreased cotton yield even though there was a 33% decrease in the amount of cotton cultivated due to the intercropping. It is assumed that both a basil-induced repellence against pest insects and a stimulation of beneficial epigeic fauna might be responsible for the lower pest infestation observed in intercropped plots.
The previous results emphasize that the identification of appropriate plants is a long-lasting process that is based on the screening of hundreds of species (as will be discussed in the section of push-pull strategies) or a longer history of research on each plant. Moreover, it is very important to study the chemical properties of plants such as repellent plants, to better understand their interaction with the crop and pest, and to permit future manipulation of the desired effects. For example the reduced infestation by stemborers in maize–D. uncinatum intercrops has been shown to be mediated by specific volatiles released by the repellent plant (Khan et al. 2000). Knowing the chemical properties of repellence not only permits a better understanding of the mechanisms, but it also gives the possibility to produce synthetic volatiles, to simulate those of the plant and have the potential to repel the herbivore or recruit natural enemies (Pickett et al. 1997; Khan et al. 2008a). Using molecular tools it may also be possible to modify the secondary metabolism of the plant to release a higher concentration of the repellent volatiles at all or only some stages of development (Khan et al. 2008a).
Not all pests react in the same way to repellent plants; what can be very effective for one pest is not necessarily effective on another pest. This was exemplified by the study of McIntyre et al. (2001), who intercropped banana with three leguminous crops, that had previously been reported as having repellent or insecticidal properties on different pest species of different crops. They failed to detect any negative effects of legumes on the banana weevil Cosmopolites sordidus (Germar) (Curculionidae) population and the presence of the nematodes Radopholus similis (Cobb) and Helicotylenchus spp, demonstrating that the repellence of several different organisms does not mean that a plant will be effective on other pests.
Trap plants to attract herbivores. Trap crops can be plants of a preferred growth stage, cultivar, variety, or species that are more attractive to the pest than the main crop. Thus trap crops reduce herbivore pressure and concentrate the pest population to a limited area, where it can be easily controlled by traditional methods (Hokkanen 1991; Asman 2002; Shelton and Nault 2004; Shelton and Badenes-Perez 2006). In this literature review five studies used trap plants as intercrops to control pests (Bender et al. 1999; Smith et al. 2000; Smith and McSorley 2000; Badenes-Perez et al. 2005; Bullas-Appleton et al. 2005). As for the repellent plants, the effective identification and use of trap plants will depend on an exhaustive screening of the potential trap crop (Khan et al. 2000), its effectiveness when using different crops or different pests and the importance of local differences in abiotic and biotic factors (Khan et al. 2008b). For example, Bender et al. (1999) used Indian mustard Brassica juncea (L.) Czern. intercropped in cabbage to study its effectiveness on controlling lepidopterous larvae, mainly of the diamondback moth P. xylostella. In the introduction of their study they already report contradictory results of the effectiveness of this potential trap species on the diamondback moth in cabbage in regions as different as Taiwan, India, and Hawaii. They tested the effectiveness of this trap species in Texas and concluded that there was no effect of intercropping cabbage with Indian mustard on any lepidopterous larvae. The actual causes of the differences achieved using the same trap plant in the same crop on the same pest remains inconclusive. However, it is clear that regional differences in biotic or abiotic factors could determine the effectiveness of such a practice. A similar case is reported in the paper by Smith and McSorley (2000) who studied the effect of intercropping eggplant Solanum melongena L. (Solanaceae) as a trap crop for management of whiteflies Bemisia argentifolii Bellows & Perring (Aleyrodidae) on bean Phaseolus vulgaris L. (Fabaceae). They report no effect of the eggplant intercropping system on the density of eggs and nymphs. This experiment exemplifies that the trap plant used was not effective under their growing conditions and they report that air currents determine the migration of adult whiteflies into plots, showing again that abiotic factors can be playing a crucial role.
The importance of determining if a reduced pest pressure translates into an increased productivity is a concern in the studies with trap crops. Only one study showed the effect of an attractive plant on pest suppression and production. Bullas- Appleton et al. (2005) investigated the effect of inter-planting the highly susceptible cultivar Berna Dutch brown bean as a trap crop in the moderately susceptible cultivar Stingray white bean P. vulgaris on pest pressure and yield. Although they reported that at the beginning of the season intercropping reduced damage on the plants by potato leafhoppers Empoasca fabae (Harris) (Cicadellidae), this effect disappeared at the end of the season, and there was no effect of intercropping with trap plants on yield.
The integrated use of repellent and attractive plant stimuli: the push-pull strategy. From the previous section we could infer that repellent stimuli seem to be very effective to reduce pest pressure and increase yield, while trap plants seem not to be as effective and their effects on production remain unclear. One possible reason for the mixed results when using trap plants is that the local attraction sought in trap crops also causes a regional attraction that increases the presence of the pest in the field since they are more attracted from outside the field by the trap plants (Vandermeer 1989). This negative effect could be compensated for by the integrated use of behaviormodifying stimuli to manipulate the distribution and abundance of pests, which has been named a “push-pull” strategy. This strategy is based on selectively increasing plant diversity to decrease pest pressure by identifying key plants that repel herbivores to make the protected culture unattractive for the pests (push) (Vanhuis 1991; Lapointe et al. 2003), while at the same time using trap plants that lure the pest toward them (pull) (Hokkanen 1991). A review on the principles of this strategy and the current knowledge is presented by Cook et al. (2007).
Only three studies in our literature review evaluated the effect of push-pull strategies as pest management systems (Midega et al. 2006; Khan et al. 2008b; Midega et al. 2008). All three studies were performed by the same group of investigators and are based on the same system. They developed a push-pull strategy to control the corn stemborers C. partellus and B. fusca in maize fields from Kenya. This strategy is based on the use of herbaceous plants of economic importance. The push stimulus is an intercrop of the forage legume D. uncinatum, and border rows of Napier grass Pennisetum purpureum Schumach. (Poaceae) exert the pull effect. This practice enhanced the abundance of natural enemies like spiders (Midega et al. 2008), increased predation rates of C. partellus (Midega et al. 2006) and reduced oviposition of C. partellus (Midega et al. 2006). Khan et al. (2008b) evaluated the effectiveness of this attractive-repellent practice under farmers’ conditions, comparing the push-pull technology against maize monocrops in 280 farms. Field surveys agree with the farmers’ perception that the push-pull strategy reduced stemborers and increased yield. Besides controlling the stem borers and increasing yield, witchweed (which decreases maize yield) is also controlled (Khan et al. 2008b). Although the push-pull technology seems to be achieving more than the expected results of a diversification practice on pest suppression and yield increase, we are aware that these results were only obtained as a consequence of many years of studying the system and its effectiveness (as can be inferred from the following studies: Khan et al. 1997; Khan et al. 2000; Khan and Pickett 2004; Khan et al. 2006b; Cook et al. 2007). The starting point to develop this technology involved a screening process of several hundred plant species, mainly of the family Poaceae, but also Cyperaceae, Thyphaceae and some Fabaceae (Khan et al. 2000). The attack rate by the different species of stem borers was examined and the colonization rate was taken to choose potential trap plants (as being those species with the highest colonization rates) and potential repellent plants (as being the least attractive plants). The two most attractive crop plants were Napier grass and Sudan grass, Sorghum sudanensis Stapf (Poaceae), while the most repellent plants were molasses grass, Melinis minutiflora Beauv. (Poaceae) and two legume species, silverleaf, D. uncinatum, and greenleaf, D. intortum (Mill.) Urb. (Fabaceae) (Khan et al. 2000). The legumes had the added advantage of suppressing development of the problematic weed S. hermonthica. With these potentially effective trap and repellent plants, experiments where performed in 1996. Napier grass was highly effective as a trap plant since it attracted most of the oviposition but at the same time reduced larval survival on the plant to 20% (in comparison with 80% on maize) (Khan et al. 2000). The effect was caused by the production of sticky sap by the Napier grass that trapped and killed the larvae (Khan and Pickett 2004). This effect was confirmed in further years of experiments that showed a yield improvement of more than 1 t/ha (Khan et al. 2000). The effectiveness of intercropping with the repellent plants was also confirmed in the field showing that the use of M. minutiflora and Desmodium significantly reduced the presence of the stemborers. The rate at which the repellent plants had to be intercropped in the fields was also assessed in further studies determining that M. minutiflora would be ideally planted at a density of 1:3 although it could be planted in densities of 1:10 while still achieving the expected results (Khan et al. 2000). After choosing the plants responsible for pest control, the mechanisms behind the effect were analyzed to increase the robustness and reliability of this pest control method. Plants use indirect defenses such as volatile organic compounds (VOCs) to attract or repel herbivores and their natural enemies (Karban and Baldwin 1997). Khan et al. (2008a) reported that for stem borer control, the plant chemistry responsible involves release of attractant VOCs (hexanal, (E)-2-hexenal, (Z)-3-hexen-1-ol, (Z)-3-hexen-yl acetate) from the trap plants and repellent VOCs ((E)-ocimene, ß- terpinolene, ß-caryophyllene, humulene, (E)-4,8-dimethyl- 1,3,7-nonatriene, ß-cedrene)) from the intercrops. If the selected plant can have additional properties that meet other farmer needs like increased nitrogen input in the soil or weed suppression, these qualities should be promoted to achieve multiple goals with only one plant. Such is the case of the repellent plant Desmodium uncinatum, which has a series of very astonishing properties. For example, the weed suppressing property is achieved by a blend of secondary metabolites in the root exudates that include seed germination stimulants and at the same time post-germination inhibitors resulting in “suicidal germination” (Tsanuo et al. 2003). Not only its weed suppressive properties but also the fact that Desmodium is a legume that increases nitrogen availability in the soil that improves land productivity, and increases gross cash returns (e.g. Khan et al. 2001) makes it highly attractive. At the same time, farmers can use this species as a nutritious and perennial fodder for cattle improving the productivity of meat and milk. Moreover the seeds of D. uncinatum represent a valuable commodity that has a local high demand among different groups of farmers (Khan et al. 2000). Screening for multiple properties can therefore increase the advantages of diversifying a crop, by supplying natural fertilizers, herbicides, pesticides and also providing fodder for cattle.
But development of the push-pull strategy does not end here. Khan et al. (2008a) also studied the adoption of this practice as a technological package by farmers, showing that by 2007 it was already adopted by thousands of farmers in eastern Africa and the program is still expanding (Khan et al. 2008b). The implementation of this push-pull technology has been shown to increase maize yields by 30%, providing the best evidence that diversification practices are useful in managing pests, increasing yield and moreover giving farmers the possibility of additional income, without an intensive use of pesticides.
Summary
Our literature review revealed contradictory effects of increased diversity on natural enemies, herbivores and production, and the expected results of reduced pest damage were only achieved in 50% of the cases. However, some examples demonstrate that diversification practices can translate into a successful management technology that is adopted by thousands of farmers. The current available data suggest a series of steps that should be taken to design successful and competitive diversification practices that can be adopted by the farmers:
- Gather precise information on the natural history of the pest and their natural enemies to selectively provide resources and shelter for the natural enemies, but not for the pest.
- Take into account the farmer’s needs to choose the “right” plant(s).
- Be open in the search for the appropriate functional plant and screen as many plants as possible.
- Favor plants that fulfill more than one function at the same time.
- Evaluate the effect of the chosen plant(s) on pests, natural enemies, crop damage, crop development and yield.
- Study the effectiveness of different arrangement patterns.
- Perform comparative field experiments at different locations and in different years to define the limitations of the proposed practice.
- Perform an economic study comparing the conventional methods with the proposed practice.
- Evaluate the labor intensity of the practice and the willingness of the farmer to implement it.
- Reach a mechanistic understanding of how the selected plant achieves the expected results to reinforce those characteristics on the selected plants or search for them in other plants.
- Test if the combination of several different functional plants leads to a synergistic effect on pest suppression and crop yield.
- Distribute the knowledge among farmers, including onfarm experiments where farmers evaluate and quantify the effectiveness of the practice.
Acknowledgements
The authors want to thank Andre Kessler for discussion and Elizabeth Wheat and Jason Tylianakis for helpful comments on the manuscript. K. Poveda was funded by the German Science Foundation (DFG)
Cited literature
ADENIYI, O. R. 2001. An economic evaluation of intercropping with tomato and okra in a rain forest zone of Nigeria. Journal of Horticultural Science & Biotechnology 76 (3): 347-349. [ Links ]
ALTIERI, M. A.; NICHOLLS, C. 1994. Biodiversity and Pest Management in Agroecosystems, New York, USA, Haworth Press. 214 p. [ Links ]
ASMAN, K. 2002. Trap cropping effect on oviposition behaviour of the leek moth Acrolepiopsis assectella and the diamondback moth Plutella xylostella. Entomologia Experimentalis et Applicata 105 (2): 153-164. [ Links ]
BADENES-PEREZ, F. R.; SHELTON, A. M.; NAULT, B. A. 2005. Using yellow rocket as a trap crop for diamondback moth (Lepidoptera: Plutellidae). Journal of Economic Entomology 98 (3): 884-890. [ Links ]
BAGGEN, L. R.; GURR, G. M. 1998. The influence of food on Copidosoma koehleri (Hymenoptera Encyrtidae), and the use of flowering plants as a habitat management tool to enhance biological control of potato moth, Phthorimaea operculella (Lepidoptera: Gelechiidae). Biological Control 11: 9-17. [ Links ]
BEGUM, M.; GURR, G. M.; WRATTEN, S. D.; HEDBERG, P. R.; NICOL, H. I. 2006. Using selective food plants to maximize biological control of vineyard pests. Journal of Applied Ecology 43 (3): 547-554. [ Links ]
BELL, V. A.; BRIGHTWELL, R. J.; LESTER, P. J. 2006. Increasing vineyard floral resources may not enhance localised biological control of the leafroller Epiphyas postvittana (Lepidoptera: Tortricidae) by Dolichogenidea spp. (Hymenoptera: Braconidae) parasitoids. Biocontrol Science and Technology 16 (10): 1031- 1042. [ Links ]
BENDER, D. A.; MORRISON, W. P.; FRISBIE, R. E. 1999. Intercropping cabbage and Indian mustard for potential control of lepidopterous and other insects. Hortscience 34 (2): 275-279. [ Links ]
BERNDT, L. A.; WRATTEN, S. D.; SCARRATT, S. L. 2006. The influence of floral resource subsidies on parasitism rates of leafrollers (Lepidoptera: Tortricidae) in New Zealand vineyards. Biological Control 37 (1): 50-55. [ Links ]
BJORKMAN, M.; HAMBACK, P. A.; RAMERT, B. 2007. Neighbouring monocultures enhance the effect of intercropping on the turnip root fly (Delia floralis). Entomologia Experimentalis et Applicata 124 (3): 319-326. [ Links ]
BROWN, M. W.; GLENN, D. M. 1999. Ground cover plants and selective insecticides as pest management tools in apple orchards. Journal of Economic Entomology 92 (4): 899-905. [ Links ]
BUKOVINSZKY, T.; TREFAS, H.; VAN LENTEREN, J. C.; VET, L. E. M.; FREMONT, J. 2004. Plant competition in pestsuppressive intercropping systems complicates evaluation of herbivore responses. Agriculture Ecosystems & Environment 102 (2): 185-196. [ Links ]
BULLAS-APPLETON, E. S.; GILLARD, C.; SCHAAFSMA, A. W. 2005. Aggregation of potato leaf hoppers, Empoasca fabae (Harris) (Homoptera: Cicadellidae), on a trap crop in an edible bean, Phaseolus vulgaris L., intercropping system. Canadian Journal of Plant Science 85 (1): 237-242. [ Links ]
BUTTS, R. A.; FLOATE, K. D.; DAVID, M.; BLACKSHAW, R. E.; BURNETT, P. A. 2003. Influence of intercropping canola or pea with barley on assemblages of ground beetles (Coleoptera: Carabidae). Environmental Entomology 32 (3): 535-541. [ Links ]
CAI, H. J.; LI, Z. S.; YOU, M. S. 2007. Impact of habitat diversification on arthropod communities: a study in the fields of Chinese cabbage, Brassica chinensis. Insect Science 14 (3): 241-249. [ Links ]
CARDINALE, B. J.; HARVEY, C. T.; GROSS, K.; IVES, A. R. 2003. Biodiversity and biocontrol: emergent impacts of a multi-enemy assemblage on pest suppression and crop yield in an agroecosystem. Ecology Letters 6 (9): 857-865. [ Links ]
CHABI-OLAYE, A.; NOLTE, C.; SCHULTHESS, F.; BORGEMEISTER, C. 2005a. Abundance, dispersion and parasitism of the stem borer Busseola fusca (Lepidoptera: Noctuidae) in maize in the humid forest zone of southern Cameroon. Bulletin of Entomological Research 95 (2): 169-177. [ Links ]
CHABI-OLAYE, A.; NOLTE, C.; SCHULTHESS, F.; BORGEMEISTER, C. 2005b. Relationships of intercropped maize, stem borer damage to maize yield and land-use efficiency in the humid forest of Cameroon. Bulletin of Entomological Research 95 (5): 417-427. [ Links ]
CIVIDANES, F. J.; BARBOSA, J. C. 2001. Effects of no-tillage and of soybean-corn intercropping on natural enemies and pests. Pesquisa Agropecuaria Brasileira 36 (2): 235-241. [ Links ]
COLLEY, M. R.; LUNA, J. M. 2000. Relative attractiveness of potential beneficial insectary plants to aphidophagous hoverflies (Diptera: Syrphidae). Environmental Entomology 29 (5): 1054- 1059. COOK, S. M.; KHAN, Z. R.; PICKETT, J. A. 2007. The use of pushpull strategies in integrated pest management. Annual Review of Entomology 52: 375-400. [ Links ] [ Links ]
DEN BELDER, E.; ELDERSON, J.; VEREIJKEN, P. F. G. 2000. Effects of undersown clover on host-plant selection by Thrips tabaci adults in leek. Entomologia Experimentalis et Applicata 94 (2): 173-182. [ Links ]
DIDHAM, R. K.; GHAZOUL, J.; STORK, N. E.; DAVIS, A. J. 1996. Insects in fragmented forests: a functional approach. Trends in Ecology & Evolution 11 (6): 255-260. [ Links ]
DUALE, A. H.; NWANZE, K. F. 1999. Incidence and distribution in sorghum of the spotted stem borer Chilo partellus and associated natural enemies in farmers’ fields in Andhra Pradesh and Maharashtra states. International Journal of Pest Management 45 (1): 3-7. [ Links ]
ENGLISH-LOEB, G.; RHAINDS, M.; MARTINSON, T.; UGINE, T. 2003. Influence of flowering cover crops on Anagrus parasitoids (Hymenoptera: Mymaridae) and Erythroneura leafhoppers (Homoptera: Cicadellidae) in New York vineyards. Agricultural and Forest Entomology 5 (2): 173-181. [ Links ]
FINCH, S.; BILLIALD, H.; COLLIER, R. H. 2003. Companion planting - do aromatic plants disrupt host-plant finding by the cabbage root fly and the onion fly more effectively than nonaromatic plants? Entomologia Experimentalis et Applicata 109 (3): 183-195. [ Links ]
FINCH, S.; COLLIER, R. H. 2000. Host-plant selection by insects - a theory based on ‘appropriate/inappropriate landings’ by pest insects of cruciferous plants. Entomologia Experimentalis et Applicata 96 (2): 91-102. [ Links ]
FINKE, D. L.; DENNO, R. F. 2004. Predator diversity dampens trophic cascades. Nature 429: 407-410. [ Links ]
FITZGERALD, J. D.; SOLOMON, M. G. 2004. Can flowering plants enhance numbers of beneficial arthropods in UK apple and pear orchards? Biocontrol Science and Technology 14 (3): 291-300. [ Links ]
FRERE, I.; FABRY, J.; HANCE, T. 2007. Apparent competition or apparent mutualism? An analysis of the influence of rose bush strip management on aphid population in wheat field. Journal of Applied Entomology 131 (4): 275-283. [ Links ]
GAO, F.; GE, F.; LIU, X.; SONG, Y. 2008. Impact of insecticides on the structure and productivity of insect pest and natural enemy communities associated with intercropping in cotton agroecosystems. International Journal of Pest Management 54 (2): 103-114. [ Links ]
GIANOLI, E.; RAMOS, I.; ALFARO-TAPIA, A.; VALDEZ, Y.; ECHEGARAY, E. R.; YABAR, E. 2006. Benefits of a maize-bean-weeds mixed cropping system in Urubamba Valley, Peruvian Andes. International Journal of Pest Management 52 (4): 283- 289. [ Links ]
GIRMA, H.; RAO, M. R.; SITHANANTHAM, S. 2000. Insect pests and beneficial arthropod populations under different hedgerow intercropping systems in semiarid Kenya. Agroforestry Systems 50 (3): 279-292. [ Links ]
GOVINDARAJAN, M.; RAO, M. R.; MATHUVA, M. N.; NAIR, P. K. R. 1996. Soil-water and root dynamics under hedgerow intercropping in semiarid Kenya. Agronomy Journal 88 (4): 513- 520. [ Links ]
GURR, G. M.; WRATTEN, S.; ALTIERI, M. A. 2004. Ecological Engineering for Pest Management: advances in habitat manipulation for arthropods. CABI. Wallington. CABI. Wallingford. UK. 252 p. [ Links ]
GURR, G. M.; WRATTEN, S. D. 1999. ‘Integrated biological control’: a proposal for enhancing success in biological control. International Journal of Pest Management 45 (2): 81-84. [ Links ]
GURR, G. M.; WRATTEN, S. D.; LUNA, J. M. 2003. Multi-function agricultural biodiversity: pest management and other benefits. Basic and Applied Ecology 4 (2): 107-116. [ Links ]
GUVENC, I. N.; YILDIRIM, E. 2006. Increasing productivity with intercropping systems in cabbage production. Journal of Sustainable Agriculture 28 (4): 29-44. [ Links ]
HANNA, R.; ZALOM, F. G.; ROLTSCH, W. J. 2003. Relative impact of spider predation and cover crop on population dynamics of Erythroneura variabilis in a raisin grape vineyard. Entomologia Experimentalis et Applicata 107 (3): 177-191. [ Links ]
HARVEY, C. T.; EUBANKS, M. D. 2004. Effect of habitat complexity on biological control by the red imported fire ant (Hymenoptera: Formicidae) in collards. Biological Control 29 (3): 348-358. [ Links ]
HEEMSBERGEN, D. A.; BERG, M. P.; LOREAU, M.; VAN HAJ, J. R.; FABER, J. H.; VERHOEF, H. A. 2004. Biodiversity effects on soil processes explained by interspecific functional dissimilarity. Science 306 (5698): 1019-1020. [ Links ]
HOKKANEN, H. M. T. 1991. Trap cropping in pest management. Annual Review of Entomology 36: 119-138. [ Links ]
HOOKS, C. R. R.; JOHNSON, M. W. 2004. Using undersown clovers as living mulches: effects on yields, lepidopterous pest infestations, and spider densities in a Hawaiian broccoli agroecosystem. International Journal of Pest Management 50 (2): 115-120. [ Links ]
HOOKS, C. R. R.; JOHNSON, M. W. 2006. Population densities of herbivorous lepidopterans in diverse cruciferous cropping habitats: effects of mixed cropping and using a living mulch. Biocontrol 51 (4): 485-506. [ Links ]
JONES, G. A.; GILLETT, J. L. 2005. Intercropping with sunflowers to attract beneficial insects in organic agriculture. Florida Entomologist 88 (1): 91-96. [ Links ]
JONES, G. A.; SIEVING, K. E. 2006. Intercropping sunflower in organic vegetables to augment bird predators of arthropods. Agriculture Ecosystems & Environment 117 (2-3): 171-177. [ Links ]
KAMUNYA, S. M.; WACHIRA, F. N.; LANG’AT, J.; OTIENO, W.; SUDOI, V. 2008. Integrated management of root knot nematode (Meloidogyne spp.) in tea (Camellia sinensis) in Kenya. International Journal of Pest Management 54 (2): 129-136. [ Links ]
KARBAN, R.; BALDWIN, I. T. (1997) Induced responses to herbivory, Chicago and London, The University of Chicago Press. [ Links ]
KHAN, Z.; PICKETT, J. A. 2004. The ‘push-pull’ strategy for stemborer management: a case study in exploiting biodiversity and chemical ecology, p. 255-164 In: Gurr, G. M., Wratten, S. & Altieri, M. A. (Eds.) Ecological Engineering for Pest management: Advances in Habitat Manipulation for Arthropods. CABI, Wallingford, UK, CABI Publishing.“ [ Links ]
KHAN, Z. R.; AMPONGNYARKO, K.; CHILISWA, P.; HASSANALI, A.; KIMANI, S.; LWANDE, W.; OVERHOLT, W. A.; PICKETT, J. A.; SMART, L. E.; WADHAMS, L. J.; WOODCOCK, C. M. 1997. Intercropping increases parasitism of pests. Nature 388 (6643): 631-632. [ Links ]
KHAN, Z. R.; JAMES, D. G.; MIDEGA, C. A. O.; PICKETT, J. A. 2008a. Chemical ecology and conservation biological control. Biological Control 45 (2): 210-224. [ Links ]
KHAN, Z. R.; MIDEGA, C. A. O.; AMUDAVI, D. M.; HASSANALI, A.; PICKETT, J. A. 2008b. On-farm evaluation of the ‘pushpull’ technology for the control of stemborers and striga weed on maize in western Kenya. Field Crops Research 106 (3): 224- 233. [ Links ]
KHAN, Z. R.; MIDEGA, C. A. O.; HASSANALI, A.; PICKETT, J. A.; WADHAMS, L. J.; WANJOYA, A. 2006a. Management of witchweed, Striga hermonthica, and stemborers in sorghum, Sorghum bicolor, through intercropping with greenleaf desmodium, Desmodium intortum. International Journal of Pest Management 52 (4): 297-302. [ Links ]
KHAN, Z. R.; PICKETT, J. A.; VAN DEN BERG, J.; WADHAMS, L. J.; WOODCOCK, C. M. 2000. Exploiting chemical ecology and species diversity: stem borer and striga control for maize and sorghum in Africa. Pest Management Science 56 (11): 957-962. [ Links ]
KHAN, Z. R.; PICKETT, J. A.; WADHAMS, L. J.; HASSANALI, A.; MIDEGA, C. A. O. 2006b. Combined control of Striga hermonthica and stemborers by maize-Desmodium spp. intercrops. Crop Protection 25 (9): 989-995. [ Links ]
KREBS, J. R.; WILSON, J. D.; BRADBURY, R. B.; SIRIWARDENA, G. M. 1999. The second silent spring? Nature 400 (6745): 611-612. [ Links ]
KRUESS, A.; TSCHARNTKE, T. 1994. Habitat fragmentation, species loss, and biological control. Science 264 (5165): 1581- 1584. [ Links ]
KRUESS, A.; TSCHARNTKE, T. 2000. Species richness and parasitism in a fragmented landscape: experiments and field studies with insects on Vicia sepium. Oecologia 122: 129-137. [ Links ]
LAL, R. B.; DE, R.; SINGH, R. K. 1978. Legume contribution to fertilizer economy in legume-cereal rotations. Indian Journal of Agricultural Sciences 48 (7): 419-424. [ Links ]
LALE, N. E. S.; SASTAWA, B. M. 2000. Evaluation of host plant resistance, sowing date modification and intercropping as methods for the control of Mylabris and Coryna species (Coleoptera: Meloidae) infesting pearl millet in the Nigerian Sudan savanna. Journal of Arid Environments 46 (3): 263-280. [ Links ]
LAMONDIA, J. A.; ELMER, W. H.; MERVOSH, T. L.; COWLES, R. S. 2002. Integrated management of strawberry pests by rotation and intercropping. Crop Protection 21 (9): 837-846. [ Links ]
LANDIS, D. A.; WRATTEN, S. D.; GURR, G. M. 2000. Habitat management to conserve natural enemies of arthropod pests in agriculture. Annual Review of Entomology 45: 175-201. [ Links ]
LAPOINTE, S. L.; MCKENZIE, C. L.; HUNTER, W. B. 2003. Toxicity and repellency of Tephrosia candida to larval and adult diaprepes root weevil (Coleoptera: Curculionidae). Journal of Economic Entomology 96 (3): 811-816. [ Links ]
LAVANDERO, B.; WRATTEN, S. D.; DIDHAM, R. K.; GURR, G. 2006. Increasing floral diversity for selective enhancement of biological control agents: A double-edged sward? Basic and Applied Ecology 7 (3): 236-243. [ Links ]
LEE, J. C.; HEIMPEL, G. E. 2005. Impact of flowering buckwheat on lepidopteran cabbage pests and their parasitoids at two spatial scales. Biological Control 34 (3): 290-301. [ Links ]
MAGNUSSON, E.; CRANFIELD, J. A. L. 2005. Consumer demand for pesticide free food products in Canada: a probit analysis. Canadian Journal of Agricultural Economics-Revue Canadienne D Agroeconomie 53 (1): 67-81. [ Links ]
MATSON, P. A.; PARTON, W. J.; POWER, A. G.; SWIFT, M. J. 1997. Agricultural intensification and ecosystem properties. Science 277 (5325): 504-509. [ Links ]
MATTHIES, D. B.; SCHMID-HEMPEL, P. 1995. The importance of population processes for the maintenance of biological diversity. GAIA 4 (4): 199-209. [ Links ]
MCINTYRE, B. D.; GOLD, C. S.; KASHAIJA, I. N.; SSALI, H.; NIGHT, G.; BWAMIKI, D. P. 2001. Effects of legume intercrops on soil-borne pests, biomass, nutrients and soil water in banana. Biology and Fertility of Soils 34 (5): 342-348. [ Links ]
MIDEGA, C. A. O.; KHAN, Z. R.; VAN DEN BERG, J.; OGOL, C.; DIPPENAAR-SCHOEMAN, A. S.; PICKETT, J. A.; WADHAMS, L. J. 2008. Response of ground-dwelling arthropods to a ‘push-pull’ habitat management system: spiders as an indicator group. Journal of Applied Entomology 132 (3): 248- 254. [ Links ]
MIDEGA, C. A. O.; KHAN, Z. R.; VAN DEN BERG, J.; OGOL, C.; PICKETT, J. A.; WADHAMS, L. J. 2006. Maize stemborer predator activity under ‘push-pull’ system and Bt-maize: A potential component in managing Bt resistance. International Journal of Pest Management 52 (1): 1-10. [ Links ]
MORLEY, K.; FINCH, S.; COLLIER, R. H. 2005. Companion planting-behaviour of the cabbage root fly on host plants and non-host plants. Entomologia Experimentalis et Applicata 117 (1): 15-25. [ Links ]
NABIRYE, J.; NAMPALA, P.; OGENGA-LATIGO, M. W.; KYAMANYWA, S.; WILSON, H.; ODEKE, V.; ICEDUNA, C.; ADIPALA, E. 2003. Farmer-participatory evaluation of cowpea integrated pest management (IPM) technologies in Eastern Uganda. Crop Protection 22 (1): 31-38. [ Links ]
NDEMAH, R.; SCHULTHESS, F.; KORIE, S.; BORGEMEISTER, C.; POEHLING, H. M.; CARDWELL, K. 2003. Factors affecting infestations of the stalk borer Busseola fusca (Lepidoptera: Noctuidae) on maize in the forest zone of Cameroon with special reference to scelionid egg parasitoids. Environmental Entomology 32 (1): 51-60. [ Links ]
NGEVE, J. M. 2003. The cassava root mealybug (Stictococcus vayssierei Richard) (Homoptera: Stictococcidae): a threat to cassava production and utilization in Cameroon. International Journal of Pest Management 49 (4): 327-333. [ Links ]
OGOL, C.; SPENCE, J. R.; KEDDIE, A. 1999. Maize stem borer colonization, establishment and crop damage levels in a maizeleucaena agroforestry system in Kenya. Agriculture Ecosystems & Environment 76 (1): 1-15. [ Links ]
OSTMAN, O.; EKBOM, B.; BENGTSSON, J. 2003. Yield increase attributable to aphid predation by ground-living polyphagous natural enemies in spring barley in Sweden. Ecological Economics 45 (1): 149-158. [ Links ]
PARAJULEE, M. N.; SLOSSER, J. E. 1999. Evaluation of potential relay strip crops for predator enhancement in Texas cotton. International Journal of Pest Management 45 (4): 275-286. [ Links ]
PATT, J. M.; HAMILTON, G. C.; LASHOMB, J. H. 1997. Foraging success of parasitoid wasps on flowers: interplay of insect morphology, floral architecture and searching behavior. Entomologia Experimentalis et Applicata 83 (1): 21-30. [ Links ]
PERFECTO, I.; VANDERMEER, J. H.; BAUTISTA, G. L.; NUNEZ, G. I.; GREENBERG, R.; BICHIER, P.; LANGRIDGE, S. 2004. Greater predation in shaded coffee farms: the role of resident neotropical birds. Ecology 85 (10): 2677-2681. [ Links ]
PERRIN, R. M.; PHILLIPS, M. L. 1978. Some effects of mixed cropping on the population dynamics of insect pests. Entomologia Experimentalis et Applicata 24 (3): 585-593. [ Links ]
PICKETT, J. A.; WADHAMS, L. J.; WOODCOCK, C. M. 1997. Developing sustainable pest control from chemical ecology. Agriculture Ecosystems & environment 64 (2): 149-156. [ Links ]
PITAN, O. O. R.; OLATUNDE, G. O. 2006. Effects of intercropping tomato (Lycopersicon esculentum) at different times with cowpea (Vigna unguiculata) or okra (Abelmoschus esculentus) on crop damage by major insect pests. Journal of Agricultural Science 144: 361-368. [ Links ]
PONTIN, D. R.; WADE, M. R.; KEHRLI, P.; WRATTEN, S. D. 2006. Attractiveness of single and multiple species flower patches to beneficial insects in agroecosystems. Annals of Applied Biology 148 (1): 39-47. [ Links ]
RAND, T. A.; TYLIANAKIS, J. M.; TSCHARNTKE, T. 2006. Spillover edge effects: the dispersal of agriculturally subsidized insect natural enemies into adjacent natural habitats. Ecology Letters 9 (5): 603-614. [ Links ]
RAO, M. R.; MATHUVA, M. N. 2000. Legumes for improving maize yields and income in semi-arid Kenya. Agriculture Ecosystems & Environment 78 (2): 123-137. [ Links ]
ROOT, R. B. 1973. Organization of a plant-arthropod associations in simple and diverse habitats: the fauna of collards (Brassica oleracea). Ecological Monographs 43: 95-118. [ Links ]
RUKAZAMBUGA, N.; GOLD, C. S.; GOWEN, S. R.; RAGAMA, P. 2002. The influence of crop management on banana weevil, Cosmopolites sordidus (Coleoptera: Curculionidae) populations and yield of highland cooking banana (cv. Atwalira) in Uganda. Bulletin of Entomological Research 92 (5): 413-421. [ Links ]
RUSSELL, E. 1989. Enemies hypothesis: a review of the effect of vegetational diversity on predatory Insects and Parasitoids. Annals of the Entomology Society of America 18 (4): 590-599. [ Links ]
SASTAWA, B. M.; LAWAN, M.; MAINA, Y. T. 2004. Management of insect pests of soybean: effects of sowing date and intercropping on damage and grain yield in the Nigerian Sudan savanna. Crop Protection 23 (2): 155-161. [ Links ]
SCHADER, C.; ZALLER, J. G.; KOPKE, U. 2005. Cotton-basil intercropping: effects on pests, yields and economical parameters in an organic field in Fayoum, Egypt. Biological Agriculture & Horticulture 23 (1): 59-72. [ Links ]
SCHULTHESS, F.; CHABI-OLAYE, A.; GOUNOU, S. 2004. Multitrophic level interactions in a cassava-maize mixed cropping system in the humid tropics of West Africa. Bulletin of Entomological Research 94 (3): 261-272. [ Links ]
SEKAMATTE, B. M.; OGENGA-LATIGO, M.; RUSSELL-SMITH, A. 2003. Effects of maize-legume intercrops on termite damage to maize, activity of predatory ants and maize yields in Uganda. Crop Protection 22 (1): 87-93. [ Links ]
SHELTON, A. M.; BADENES-PEREZ, E. 2006. Concepts and applications of trap cropping in pest management. Annual Review of Entomology 51: 285-308. [ Links ]
SHELTON, A. M.; NAULT, B. A. 2004. Dead-end trap cropping: a technique to improve management of the diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae). Crop Protection 23 (6): 497-503. [ Links ]
SHOWLER, A. T.; GREENBERG, S. M. 2003. Effects of weeds on selected arthropod herbivore and natural enemy populations, and on cotton growth and yield. Environmental Entomology 32 (1): 39-50. [ Links ]
SKELTON, L. E.; BARRETT, G. W. 2005. A comparison of conventional and alternative agroecosystems using alfalfa (Medicago sativa) and winter wheat (Triticum aestivum). Renewable Agriculture and Food Systems 20 (1): 38-47. [ Links ]
SMITH, H. A.; KOENIG, R. L.; MCAUSLANE, H. J.; MCSORLEY, R. 2000. Effect of silver reflective mulch and a summer squash trap crop on densities of immature Bemisia argentifolii (Homoptera: Aleyrodidae) on organic bean. Journal of Economic Entomology 93 (3): 726-731. [ Links ]
SMITH, H. A.; MCSORLEY, R. 2000. Potential of field corn as a barrier crop and eggplant as a trap crop for management of Bemisia argentifolii (Homoptera: Aleyrodidae) on common bean in North Florida. Florida Entomologist 83 (2): 145-158. [ Links ]
SMITH, H. A.; MCSORLEY, R.; IZAGUIRRE, J. A. S. 2001. Effect of intercropping common bean with poor hosts and nonhosts on numbers of immature whiteflies (Homoptera: Aleyrodidae) in the Salama Valley, Guatemala. Environmental Entomology 30 (1): 89-100. [ Links ]
SONGA, J. M.; JIANG, N.; SCHULTHESS, F.; OMWEGA, C. 2007. The role of intercropping different cereal species in controlling lepidopteran stemborers on maize in Kenya. Journal of Applied Entomology 131 (1): 40-49. [ Links ]
SPERBER, C. F.; NAKAYAMA, K.; VALVERDE, M. J.; NEVES, F. D. 2004. Tree species richness and density affect parasitoid diversity in cacao agroforestry. Basic and Applied Ecology 5 (3): 241-251. [ Links ]
STAVER, C.; GUHARAY, F.; MONTERROSO, D.; MUSCHLER, R. G. 2001. Designing pest-suppressive multistrata perennial crop systems: shade-grown coffee in Central America. Agroforestry Systems 53 (2): 151-170. [ Links ]
STRAUB, C. S.; FINKE, D. L.; SNYDER, W. E. 2008. Are the conservation of natural enemy biodiversity and biological control compatible goals? Biological Control 45 (2): 225-237. [ Links ]
SUNDERLAND, K.; SAMU, F. 2000. Effects of agricultural diversification on the abundance, distribution, and pest control potential of spiders: a review. Entomologia Experimentalis et Applicata 95 (1): 1-13. [ Links ]
TAHVANAINEN, J. O.; ROOT, R. B. 1972. The influence of vegetational diversity on population ecology of a specialized herbivore, Phyllotreta cruciferae (Coleoptera: Chrysomelidae). Oecologia 10 (4): 321-346 [ Links ]
THOMPSON, G. D. 1998. Consumer demand for organic foods: what we know and what we need to know. American Journal of Agricultural Economics 80 (5): 1113-1118. [ Links ]
TILMAN, D.; CASSMAN, K. G.; MATSON, P. A.; NAYLOR, R.; POLASKY, S. 2002. Agricultural sustainability and intensive production practices. Nature 418 (6898): 671-677. [ Links ]
TSANUO, M. K.; HASSANALI, A.; HOOPER, A. M.; KHAN, Z.; KABERIA, F.; PICKETT, J. A.; WADHAMS, L. J. 2003. Isoflavanones from the allelopathic aqueous root exudate of Desmodium uncinatum. Phytochemistry 64 (1): 265-273. [ Links ]
TYLIANAKIS, J. M.; DIDHAM, R. K.; WRATTEN, S. D. 2004. Improved fitness of aphid parasitoids receiving resource subsidies. Ecology 85 (3): 658-666. [ Links ]
UVAH, III; COAKER, T. H. 1984. Effect of mixed cropping on some insect pests of carrots and onions. Entomologia Experimentalis et Applicata 36 (2): 159-167. [ Links ]
VANDERMEER, J. 1989. The ecology of intercropping, Cambridge, UK, Cambridge University Press. 237 p. [ Links ]
VANHUIS, A. 1991. Biological methods of bruchid control in the Tropics - a review. Insect Science and its Application 12 (1-3): 87-102. [ Links ]
WÄCKERS, F. L. 2004. Assessing the suitability of flowering herbs as parasitoid food sources: flower attractiveness and nectar accessibility. Biological Control 29: 307-314. [ Links ]
WALE, M.; SCHULTHESS, F.; KAIRU, E. W.; OMWEGA, C. O. 2007. Effect of cropping systems on cereal stemborers in the coolwet and semi-arid ecozones of the Amhara region of Ethiopia. Agricultural and Forest Entomology 9 (2): 73-84. [ Links ]
WILBY, A.; THOMAS, M. B. 2002a. Are the ecological concepts of assembly and function of biodiversity useful frameworks for understanding natural pest control? Agricultural and Forest Entomology 4 (4): 237-243. [ Links ]
WILBY, A.; THOMAS, M. B. 2002b. Natural enemy diversity and pest control: patterns of pest emergence with agricultural intensification. Ecology Letters 5 (3): 353-360. [ Links ]
Recibido: 1-sep-2008 - Aceptado: 24-sep-2008


{kind=link}