










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488On-line version ISSN 2665-4385
Rev. Colomb. Entomol. vol.34 no.2 Bogotá July/Dec. 2008
Sección Agrícola
Identificación de biotipos de Spodoptera frugiperda(Lepidoptera: Noctuidae)
mediante marcadores mitocondriales y nucleares
Identification of Spodoptera frugiperda biotypes (Lepidoptera: Noctuidae) through using mitochondrial and nuclear markers
ANA MARÍA VÉLEZ-ARANGO1, RAFAEL EDUARDO ARANGO I.2, DIEGO VILLANUEVA M.3, ELIZABETH AGUILERA G.4 y CLARA INÉS SALDAMANDO B.5
1 Bióloga, Candidato M.Sc. Universidad Nacional de Colombia, sede Medellín. Corporación para Investigaciones Biológicas, CIB. Unidad de Biotecnología
Vegetal, Colombia. anamaria.velez@gmail.com.
2 Director del Laboratorio de Biotecnología Vegetal. Corporación para Investigaciones Biológicas, CIB.
3 Investigador M.Sc. Laboratorio de Biotecnología Vegetal. Corporación para Investigaciones Biológicas, CIB.
4 Investigador M.Sc. Programa de Manejo Integrado de Plagas de Corpoica, Tibaitatá.
5Profesor Asistente Ph. D. Facultad de Ciencias, Departamento de Biociencias, Universidad Nacional de Colombia, UNALMED, Calle 59A No. 63 - 20. Medellín, Colombia. Edificio 11-208. Fax 4309072. cisaldam@unal.edu.co. Autor para correspondencia
Resumen: Spodoptera frugiperda es un insecto considerado plaga primaria del cultivo de maíz y secundaria en arroz, algodón, sorgo y pastizales. Presenta dos biotipos (maíz y arroz) casi idénticos morfológicamente, pero difieren en su composición genética, resistencia hacia insecticidas y Bacillus thuringiensis, siendo el biotipo de maíz más resistente respecto al biotipo de arroz. Este trabajo es la primera identificación molecular en Colombia de estos biotipos a partir de larvas recolectadas en cultivos de maíz, arroz y algodón en el departamento del Tolima, con el uso de una PCR-RFLP a nivel del gen mitocondrial citocromo oxidasa I para identificar al biotipo de maíz, y una PCR de la región tandem FR del ADN nuclear para identificar biotipo de arroz. Los resultados diferenciaron tres poblaciones de S. frugiperda: a) biotipo de maíz, compuesto por individuos que generaron digestiones con la enzima MspI del gen COI, b) biotipo de arroz compuesto por individuos que generaron amplificaciones de peso moleculares mayores a 500 pb con el marcador FR, y finalmente c) un grupo de individuos clasificados como “híbridos” entre estos dos marcadores, producto del cruce entre hembras del biotipo de maíz y machos del biotipo de arroz y viceversa. Los individuos genotipificados como biotipo de maíz se recolectaron en cultivos de maíz y algodón exclusivamente y los individuos genotipificados como biotipo de arroz en cultivos de arroz principalmente y en bajas proporciones en cultivos de maíz y algodón.
Palabras clave: Región COI. PCR-RFLP´s. Enzima de restricción Msp1. Región FR. Híbridos.
Abstract: Spodoptera frugiperda is considered a primary pest in corn crops and a secondary pest in rice, cotton, sorghum and pastures. It exhibits two biotypes (corn and rice) that are almost identical morphologically, but differ in their genetic composition and resistance to insecticides and Bacillus thuringiensis, with the corn biotype more resistant than the rice biotype. This work is the first molecular identification in Colombia of these biotypes for larvae collected from corn, rice and cotton crops in the department of Tolima by using a PCR-RFLP of the mitochondrial gene cytochrome oxidase subunit I (COI) for the identification of corn biotype, and a PCR amplifying a FR tandem region of the nuclear DNA to identify the rice biotype. The results differentiated three populations of S. frugiperda: a) corn biotype, composed of individuals that generated digestion products with the enzyme MspI of the COI gene, b) rice biotype, composed of individuals that generated amplification products higher than 500 bp with the FR marker, and finally c) a group of individuals classified as “hybrids” between these markers, product of the cross between a corn biotype female and rice biotype male and vice versa. Individuals genotyped as the corn biotype were collected exclusively from corn and cotton crops and the individuals genotyped as the rice biotype were mainly collected from rice crops and in low proportions from corn and cotton crops.
Key words: COI region. PCR-RFLP´s. Msp1 restriction enzyme. FR region. Hybrids.
Introducción
El gusano cogollero, Spodoptera frugiperda (J. E. Smith, 1797) (Lepidoptera: Noctuidae), es un insecto tropical endémico del hemisferio occidental que se extiende desde Argentina hasta Norteamérica (López-Edwards et al. 1999; Pashley et al. 2004; Busato et al. 2004; Clark et al. 2007; Martinelli et al. 2007). Este insecto, es considerado como una de las plagas primarias de cultivos de maíz y secundaria en algodón, arroz y algunas hortalizas en Estados Unidos y Brasil (Pashley 1998; Busato et al. 2004). En 1914 apareció en Colombia asociada a cultivos de maíz en el departamento de Antioquia, posteriormente se dispersó a Caldas y Valle del Cauca, hasta establecerse en todas las áreas maiceras del país. La larva de esta polilla puede atacar alrededor de 38 cultivos y malezas al alimentarse de sus tejidos foliares y cogollos (Bustillo y Posada 1983) y por esta razón es considerada una plaga primaria en el cultivo de maíz en la gran mayoría de las zonas productoras del país, al generar pérdidas económicas de hasta un 35% (Torres y Cotes 2005) y como una plaga secundaria en los cultivos de algodón, caña de azúcar, arroz, sorgo y pastos (García et al. 2002).
En el departamento de Tolima (Colombia) esta especie ha sido el insecto que más ha interferido en los programas de manejo integrado de plagas, particularmente en los cultivos de algodón, por ello es donde más se han concentrado los trabajos de control biológico de este insecto (Álvarez y Sánchez 1983). Recientemente, en el Tolima se analizó la susceptibilidad de Heliothis virescens (F., 1777), Helicoverpa zea (Boddie, 1850), Pectinophora gossypiella (Saunders, 1843) y S. frugiperda al realizar estudios del efecto de la endotoxina Cry1Ac de cultivos transgénicos de algodón sobre estos lepidópteros (Zenner-De Polania et al. 2007). En esa investigación, se observó que la susceptibilidad de los tres primeros insectos es alta, mientras que la susceptibilidad de S. frugiperda era muy baja o casi nula (Zenner-De Polania et al. 2007) corroborando los resultados encontrados por Adamczyk et al. (1997), puesto que también demostraron que la endotoxina Cry1Ac no afectaba la supervivencia de S. frugiperda. Sin embargo, Adamczyk et al. (1997) identificaron los biotipos de S. frugiperda antes de llevar a cabo el análisis de susceptibilidad lo cual no se realizó en este último estudio en el Tolima.
En S. frugiperda se han identificado dos «biotipos» que son indistinguibles morfológicamente, pero son reconocidos como posibles nuevas especies (Dres y Mallet 2002) o razas de plantas hospederas de maíz y arroz (Pashley 1998), puesto se que encuentran con mayor frecuencia en ellos. Sin embargo, el biotipo de maíz, también se ha encontrado asociado a cultivos de sorgo y algodón, mientras que el biotipo de arroz a cultivos de arroz y pastizales (Pashley 1986; Pashley 1998; Nagoshi y Meagher 2003a, 2004; Pashley et al. 2004). Estos biotipos han sido identificados en varios países, entre ellos: Estados Unidos (Pashley 1986; Nagoshi y Meagher 2003a, b), 2004; Meagher y Gallo-Meagher 2003; Pashley et al. 2004; Nagohsi et al. 2007a, b), Puerto Rico, Guadalupe, Republica Dominicana, Jamaica, Costa Rica, México, Guyana Francesa, Ecuador (Pashley et al. 2004), Brasil (Nagoshi et al. 2007 b) y Argentina (Clark et al. 2007).
Estos biotipos han sido denominados especies verdaderas (Dres y Mallet 2002) debido a que en ellos se han observado formas de aislamiento reproductivo precigótico y postcigótico. El aislamiento precigótico es de tres tipos: 1) temporal: debido a que el biotipo de maíz tiende aparearse en los dos primeros tercios de la noche, mientras que el biotipo de arroz en el último tercio (Pashley et al. 1992), 2) comportamental parcial dado que rara vez las hembras del biotipo del maíz se aparean con los machos del biotipo del arroz, aunque el cruce recíproco si produce progenie (Pashley y Martin 1987; Pashley 1998) y 3) ecológico debido a que los biotipos se ubican principalmente en dos plantas hospederas: maíz y arroz, significando que las hembras podrían diferenciarse en su comportamiento de oviposición (Pashley et al. 2004). El aislamiento postcigótico entre los biotipos es parcial, dada la baja viabilidad de los híbridos de la F2 obtenidos a partir de cruces entre hembras del biotipo del maíz y machos del biotipo del arroz, mientras que los individuos pertenecientes al cruce recíproco producen híbridos fértiles y viables (Pashley y Martin 1987; Nagoshi y Meagher 2003a, b). Otra investigación en la que también se encontró aislamiento reproductivo en S. frugiperda fue realizada por López-Edwards et al. (1999) en México, en la que cruces de adultos entre poblaciones aisladas de cultivos de maíz, no produjeron progenie, cruces entre individuos del centro y el golfo de México con individuos de su costa Pacífica. En este ultimo estudio no se genotipificaron las poblaciones de S. frugiperda, pero si se logró concluir que el aislamiento geográfico jugaba un papel importante en el aislamiento reproductivo del insecto. Finalmente, en otro trabajo en el que se evidencia aislamiento reproductivo en Florida y Loussiana, Pashley et al. (1992) utilizaron hembras vírgenes de cada biotipo para producir trampas de feromonas, y observaron que los machos de cada biotipo de S. frugiperda preferían aquellas trampas en las que se ubicaron hembras de su mismo biotipo, comprobándose un aislamiento comportamental en la especie.
Por otro lado, en estudios realizados en Estados Unidos, se ha encontrado que en condiciones de laboratorio las larvas del biotipo del maíz muestran una mayor resistencia a componentes de insecticidas como el carbaril, diazinon, cipermetrinas, metil paration y metiomil, además de cultivos de algodón transgénico al que se le ha introducido la endotoxina Cry1AC del Bacillus thuringensis, mientras que en condiciones de campo las larvas del biotipo del arroz consumen más material vegetal que las larvas del biotipo del maíz (Adamczyk et al. 1997). Debido a que los biotipos de S. frugiperda son casi idénticos morfológicamente han sido necesarias las herramientas moleculares y bioquímicas para su diferenciación.
Los biotipos exhiben polimorfismos genéticos a nivel de cinco esterasas, siendo las enzimas B, C y D exclusivas en el biotipo de maíz y las enzimas E y F en el biotipo de arroz; los híbridos entre éstos presentan combinaciones de las esterasas de ambos biotipos, generándose individuos con esterasas B y F, C y E, entre otras (Pashley 1986; Pashley et al. 2004). A nivel del ADN mitocondrial en el gen de la citocromo oxidasa I (COI), la enzima de restricción MspI genera dos fragmentos (497 pb y 72 pb) únicamente en el biotipo del maíz y ninguna digestión en el biotipo de arroz (Levy et al. 2002; Meagher y Gallo- Meagher 2003). Estos biotipos también han sido identificados con AFLP’s (McMichael y Pashley 1999; Pashley et al. 2004; Clark et al. 2007; Martinelli et al. 2007) y con una región en tandem del ADN nuclear denominada FR (for rice) que produce amplificaciones de alto peso molecular (mayores a 500 pb) en el biotipo de arroz (Lu et al. 1994; Nagoshi y Meagher 2003a, 2004). El uso de los marcadores COI y FR ha sido de suma importancia en S. frugiperda ya que no solamente han permitido identificar los biotipos de esta especie, sino que también han servido para el reconocimiento de híbridos entre éstos. Autores como Nagoshi y Meagher (2003a, b) y Nagoshi et al. (2007a) han encontrado híbridos que presentan digestión con la enzima de restricción MspI y amplificación por encima de las 500 pb de la región FR, e híbridos sin digestión con la enzima MspI y fragmentos menores a las 500 pb en la región FR. Los híbridos entre los dos biotipos de S. frugiperda han mostrado tener una menor especificidad hacia las plantas hospederas respecto a los de los biotipos de esta especie, lo cual demuestra la relevancia que tiene la identificación molecular los biotipos y sus híbridos, para mejorar del manejo de este insecto (Nagoshi et al. 2007a).
En Colombia, los estudios con S. frugiperda han sido más enfocados en su control químico y biológico (García et al. 2002; Torres y Cotes 2005) y por ello la existencia de sus biotipos no ha sido ampliamente estudiada. Sin embargo, en un trabajo realizado por Pinzón (1992) se mencionó la posible existencia de estos biotipos en un trabajo que se basó en el desarrollo del biocontrolador Meteorus laphygmae Viereck, 1913 (Hymenoptera: Braconidae) sobre S. frugiperda en cultivos de maíz de poblaciones de El Espinal y Valle de San Juan (Tolima). Pinzón (1992) encontró que las crías provenientes de S. frugiperda de El Espinal, tuvieron un mayor parasitismo de M. laphygmae comparado con las crías del Valle de San Juan, por esta razón la autora sugiere la existencia de biotipos de S. frugiperda en el Tolima. A pesar de que éste ha sido uno de los primeros trabajos en proponer la presencia de biotipos de este insecto en Colombia, hasta el momento no se han realizado otros estudios relacionados con la identificación de biotipos lo cual es fundamental para su manejo integrado (Nagoshi y Meagher 2004); tal identificación es necesaria pues ambos biotipos podrían reflejar comportamientos diferenciales de resistencia hacia insecticidas y controladores biológicos, generando la necesidad de utilizar nuevas estrategias para su control. Además, su presencia, también es relevante desde una perspectiva ecológica y evolutiva debido a que si S. frugiperda se encuentra en un periodo de divergencia genética generada por una evolución asociada a sus plantas hospederas, este insecto podría ser utilizado como modelo para estudios de especiación ecológica (Dres y Mallet 2002; Pashley et al. 2004). Por estas razones el presente trabajo tuvo dos propósitos: a) identificar biotipos de S. frugiperda y sus respectivos híbridos con el uso de dos técnicas moleculares y b) reconocer las plantas huéspedes en las que se encontraron con mayor frecuencia cada uno de los biotipos y sus respectivos híbridos.
Materiales y Métodos
Recolecciones de campo. Los muestreos de S. frugiperda se realizaron en fincas con cultivos de rotación de algodón, maíz y arroz en el departamento del Tolima, Colombia. Estos muestreos se produjeron en tres microrregiones del departamento: norte, centro, y sur, que difieren en sus condiciones ecológicas. En el norte predominan los cultivos de café, maíz, algodón y sorgo, en el centro los cultivos de maíz, sorgo y algodón, y en el sur, cultivos de algodón, arroz y maíz, siendo esta última la zona más seca del departamento al limitar con el desierto de la Tatacoa. Estas recolecciones se llevaron a cabo durante el segundo semestre de 2006 y primer semestre de 2007 dada la rotación de los cultivos (en el primer semestre del año se producen maíz y arroz, mientras que en el segundo semestre se cultiva algodón principalmente). En cada microrregión se muestreó en cultivos de maíz y algodón, excepto en el norte donde se recolectó adicionalmente en arroz (Tabla 1).
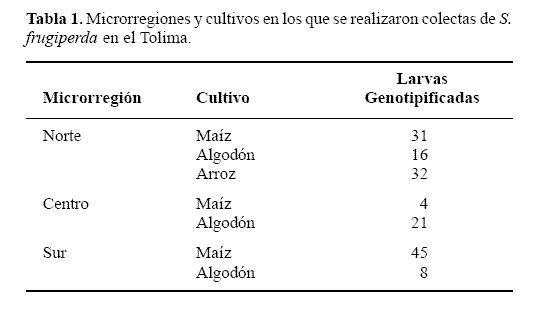
Una vez se tomaron las muestras, las larvas se ubicaron en tubos de plástico de 2,5 ml (rotulados por finca y micro-región) con etanol al 70% y posteriormente se enviaron la Unidad de Biotecnología Vegetal UNALMED-CIB (Corporación para Investigaciones Biológicas) de Medellín, donde se almacenaron en seco en una nevera a -70ºC, para su posterior genotipificación.
Caracterización molecular de S. frugiperda mediante PCRRFLP y PCR. Se genotipificaron 157 individuos mediante una PCR-RFLP de la región COI del ADN mitocondrial y una PCR de la región nuclear FR, utilizando los cebadores indicados por Nagoshi y Meagher (2003a). La extracción de ADN se realizó siguiendo el protocolo de Sambrook y Russell (2001) con algunas modificaciones. Para esto, se empleó la parte posterior del cuerpo de larvas de segundo a quinto instar, cada larva se maceró en nitrógeno líquido y se depositó en un tubo eppendorf de 1.5 µl al cual se le adicionaron 400 ml de buffer de extracción CTAB (2X) y 4 µl de ß-mercaptoetanol, cada tubo fue mezclado por medio de un vórtex y se incubó por 30 minutos a 65ºC. Posteriormente, a cada muestra se le adicionaron 400 ml de cloroformo-alcohol isoamílico en una proporción 24:1 y se centrifugó por 6 minutos a 13.000 rpm a 4ºC. A continuación se tomó el sobrenadante en un nuevo tubo y se repitió el procedimiento con un volumen equivalente de cloroformo. El nuevo sobrenadante se transfirió a un tubo, se adicionó un volumen equivalente de isopropanol y se almacenó a -20ºC por 2 horas. Después de este tiempo, las muestras se centrifugaron por 6 minutos, el sobrenadante se eliminó y el sedimento restante se lavó con 200 µl de etanol al 70% y se centrifugó por 5 minutos; este último procedimiento se repitió, el sedimento restante se secó al vacío y se resuspendió en 50 µl en buffer TE (1X) (TRIS HCL 100 mM y EDTA 10 mM, pH 8.0). Finalmente, se adicionó 1 ml de RNasa y se incubó a 37ºC por 1 µl. Se obtuvo una concentración aproximada de ADN de 50 ng.
El ADN genómico fue amplificado mediante PCR en 25 µl de mezcla de reacción que contenía 2,5 µl de buffer de Taq polimerasa (10X) (Fermentas, California U.S.A), con (NH4)2 SO4, 1,5 µl de MgCl2 25 mM (Fermentas), 0,5 µl de dNTPs (10 µM) (Invitrogen, California U.S.A), 1,0 ml de cebador forward JM76 (5’ GAGCTGAATTAGG(G/A)ACTCCAGG 3’), 1,0 µl de cebador reverse JM77 (5’ATCACCTCC (A/T)CCTGCAGGATC 3’), 0,5 ml (5µ/µl)de Taq ADN polimerasa (Fermentas), 17µl de H20 y 1,0µl de ADN (50 ng).
Para la PCR se realizó un ciclo inicial de 94ºC por 3 minutos, seguido de 30 ciclos a 94ºC por 1 minuto, 62ºC por 1 minuto y 72ºC por 1 minuto, y un ciclo de extensión final de 72ºC por 10 minutos. Posteriormente se realizó una digestión con la enzima de restricción MspI, para esto se adicionó en un nuevo tubo 300 ng de mezcla de PCR, 2 ml de Buffer 10X (Fermentas, California U.S.A) y 10 unidades de la enzima MspI, esto se completo a 30µl con H20, y se incubó a 37ºC por 2 h. Las muestras se analizaron en un gel de agarosa al 2% con bromuro de etidio 0,1% y se visualizaron en un transluminador UV.
Para la amplificación de la región FR en S. frugiperda, se realizaron reacciones de PCR que contenían 2,5 ml de buffer de Taq polimerasa (10X) (Fermentas, California U.S.A) con (NH4)2 SO4, 1,5µl de MgCl2 25 µM (Fermentas), 0,5 µl de dNTPs (10ìM) (Invitrogen), 1,0 µl de cebador FR-a, 1,0 µl de cebador FR-2, 0,5 ml (5u/ml) de Taq ADN polimerasa (Fermentas), 17 ml de H20 y 1,0 ml de ADN (50 ng). Se realizó un ciclo inicial de 94°C por 3 minutos, seguido por 40 ciclos a 94°C por 1 minuto, 62°C por 1 minuto y 72°C por 1 minuto, seguido por un ciclo extra de 72°C por 10 minutos. Las muestras fueron analizadas en un gel de agarosa al 2 % con bromuro de etidio 0,1% y se visualizaron en un transluminador UV.
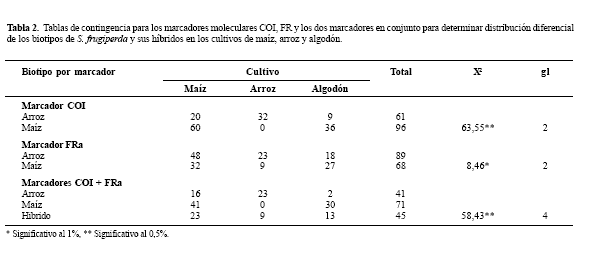
Análisis Estadístico. Debido a que los datos de los biotipos de S. frugiperda y sus respectivos híbridos son categóricos, se realizaron tablas de contingencia (Sokal y Rohlf 1995) para determinar si existe una distribución independiente de los individuos genotipificados (biotipo de maíz, arroz e híbridos) respecto a los cultivos de maíz, arroz y algodón recolectados en las tres micro-regiones (centro, norte y sur) del departamento del Tolima. Estas tablas se dividieron en tres tipos, una primera para el marcador COI, una segunda tabla con el marcador FR y una tercera con la combinación de los dos marcadores. Los análisis de datos se ejecutaron en el programa Genstat 5.0 (2003).
Resultados y Discusión
La PCR RFLP del gen mitocondrial COI produjo una amplificación de un fragmento de 569 pb y una digestión de 497 pb y 72 pb con la enzima de restricción Mspl identificando individuos pertenecientes al biotipo de maíz exclusivamente en el Tolima. Sin embargo, la visualización del fragmento de 72 pb en geles de agarosa no siempre es posible debido a su pequeño tamaño (Fig. 1). Estos resultados, concuerdan con los trabajos realizados por Levy et al. (2002), Nagoshi y Meagher (2003a) al producir las mismas bandas electroforéticas, al presentar geles en los que se observa solamente el fragmento de 497 pb y otros donde se observan los dos fragmentos. Por otro lado, los cebadores FRa-FR2 de la región en tandem FR del ADN nuclear produjeron una escalera de bandas electroforéticas por encima de los 500 pb en muestras larvales que fueron identificadas como biotipo de arroz (Fig. 2). Este marcador también ha permitido identificar el biotipo de arroz por otros autores como Lu et al. (1994), Nagoshi y Meagher (2003a) y Nagoshi et al. (2007), dado que ellos también obtuvieron bandas electroforéticas de alto peso molecular (entre 1000 y 500 pares de bases) para el biotipo de arroz.
Estos marcadores, también comprobaron la presencia de híbridos entre estos biotipos en el Tolima, definiendo dos tipos de hibridización de S. frugiperda: 1) individuos que generaron productos de digestión con la enzima MspI y a su vez amplificaciones mayores a 500 bp con los cebadores de la región FR y 2) individuos que no presentaron digestión con la enzima MspI y productos de amplificación menores a 500 pb con los cebadores de la región FR. Estos dos tipos de hibridización también fueron evidenciados en los trabajos realizados por Nagoshi y Meagher (2003a, b) quienes realizaron cruces entre estos dos biotipos (hembras de maíz y machos de arroz y viceversa), obteniendo una progenie F1 y F2 que fue genotipificada con estos dos marcadores, por esto, los geles presentados por Nagoshi y Meagher (2003a, b) fueron la base de comparación de los resultados de los geles electroforéticos obtenidos en el presente estudio.
Es importante mencionar que una diferencia entre el trabajo de Nagoshi y Meagher (2003 b) y el presente, se basa en que en el primero se utilizaron dos cebadores forward (FRc y FRa) para la identificación de los dos biotipos, mientras que en el protocolo utilizado en este estudio solamente se implementaron los cebadores FRa-FR2. Estos dos últimos cebadores amplifican la misma región del ADN nuclear que los cebadores FRc- FR2 con 56 pares de bases mas (Nagoshi y Meagher (2003b), lo que implica que el uso de cualquiera de los dos cebadores forward FR puede ser utilizado para la identificación de biotipos de esta especie.
Se genotipificaron dos biotipos de S. frugiperda y sus híbridos de un total de 157 larvas. De éstas, 71 individuos fueron clasificados como biotipo de maíz, al presentar digestión con la enzima de restricción MspI y al amplificar fragmentos menores a las 500 pb con los cebadores FR. Por otra parte, 41 individuos fueron identificados como biotipo de arroz ya que no presentaron corte con la enzima MspI y mostraron una escalera de fragmentos de la región FR de alto peso molecular (mayores a 500 pb). Además, se observaron dos clases de híbridos: 23 individuos que tuvieron digestión con la enzima de restricción MspI de la región COI y amplificación de la región FR por encima de las 500 bp y 22 individuos que no presentaron digestión con la enzima de restricción MspI de la región COI y amplificación de fragmentos menores a las 500 bp en la región FR. Los híbridos identificados en este trabajo también fueron reconocidos por Nagoshi et al. (2007 a), donde reportan que los híbridos positivos para ambos marcadores pueden ser tanto machos como hembras, mientras que los híbridos negativos para los dos marcadores solo son machos.
Por otro lado, las tres tablas de contingencia muestran que existe una distribución diferencial de los biotipos de maíz y arroz respecto a la planta hospedera debido a que en cada una de ellas se rechazó la hipótesis nula teniendo en cuenta cada marcador molecular por separado y en conjunto (Tabla 2). Se puede observar que el biotipo de maíz se encuentra exclusivamente en los cultivos de maíz y algodón al analizar las muestras larvales con los marcadores COI y FR por separado y en conjunto, mientras que el biotipo de arroz se encuentra en mayor número en el cultivo de arroz y en menor cantidad en los otros dos cultivos. Resultados similares fueron indicados por Pashley (1986, 1998) en Estados Unidos, donde observó que las larvas del biotipo de maíz se encontraron más frecuentemente alimentándose en plantas de maíz y algodón, mientras que las larvas de arroz, en plantas de arroz y pastizales. En un trabajo posterior, Pashley et al. (2004) obtuvo las mismas distribuciones de los biotipos a partir de recolecciones de adultos y larvas en cultivos de maíz, arroz y pastizales en Estados Unidos (Florida y Lousiana), Puerto Rico, Republica Dominicana, Jamaica, Guadalupe, México y Ecuador. Igualmente, Nagoshi y Meagher (2003a) observaron que machos adultos de S. frugiperda de la Florida (Estados Unidos) formaban dos grupos de individuos identificados con la enzima Msp1 en la región COI y la región FR; el primer grupo lo conformaba el biotipo de maíz y se recolectó únicamente en cultivos de maíz y un segundo grupo de individuos clasificado como biotipo de arroz, que se recolectó en pastizales y hábitat mixtos principalmente.
Los resultados de estos análisis son interesantes, al reflejar diferencias en el comportamiento de oviposición de las hembras híbridas respecto a las hembras pertenecientes a los biotipos puros, puesto que las primeras se equivocan al ovipositar en sus plantas hospederas, ya que heredan los genes de ambos biotipos parentales, por ello ubican sus posturas indiscriminadamente en cualquier cultivo; mientras que las hembras de cada biotipo son más especificas respecto a su hospedero, ya que el biotipo de maíz solo se encuentra asociado al maíz y algodón, y el biotipo de arroz principalmente al arroz (Pashley et al. 2004; Nagoshi et al. 2007a). Por otro lado, las diferencias en la distribución de los biotipos de S. frugiperda también podría explicarse por una tasa de supervivencia diferencial de las larvas de estos biotipos en cada uno de los cultivos (Pashley et al. 2004). Estas dos estrategias de la biología del insecto deberán ser tenidas en cuenta en futuras investigaciones para determinar si la asociación insecto hospedero evolucionó por preferencia a su planta hospedera o por resistencia diferencial de aleloquímicos de sus larvas a las sustancias toxicas producidas por sus hospederos (Pashley et al. 1992, 2004).
Conclusiones
S. frugiperda ha sido uno de los insectos más ampliamente estudiados por diversos investigadores en el intento de generar alternativas de manejo integrado hacia éste con el uso de controles químicos y biológicos. Sin embargo, estos métodos no han tenido en cuenta un conocimiento básico de la biología de esta polilla, debido a que se encuentra en un periodo de divergencia evolutiva, generando biotipos que se están especializando hacia dos plantas hospederas principales: el maíz y arroz. Estos biotipos representan especies incipientes que deben ser tratadas con cautela dado que, al menos en Estados Unidos, presentan un comportamiento diferencial de resistencia hacia insecticidas y Bacillus thuringensis, lo cual debe conducir futuras investigaciones a la realización de estudios de comportamiento de resistencia de estos biotipos en Colombia, para así lograr un mejor manejo de este insecto en las regiones cuyo impacto es mayor.
Agradecimientos
Los autores expresan su agradecimiento a la Universidad Nacional de Colombia, sede Medellín y a la Corporación de Investigaciones Biológicas (UNALMED-CIB), Unidad de Biotecnología Vegetal por el desarrollo de este trabajo. A CORTOLIMA por otorgar el permiso de captura y acceso a los recursos genéticos de S. frugiperda en el Tolima (resolución 843, agosto 2007) y a Jhon Alexander Agudelo por la recolección de las larvas de S. frugiperda. Este proyecto fue financiado por la Universidad Nacional de Colombia al investigador principal Clara Saldamando. Código Quipu 201010006109.
Literatura citada
ADAMCZYK, J. R.; H OLLOWAY, J. J.; LEONARD, J. W.; GRAVES, J. B. 1997. Susceptibility of fall armyworm collected from different plant hosts to selected insecticides and transgenic Bt cotton. Journal of Cotton Science 1 (1): 21-28. [ Links ]
ÁLVAREZ, J. A.; SÁNCHEZ, G. 1983. Variación en el número de instares de Spodoptera frugiperda (J. E. Smith). Revista Colombiana de Entomología 9 (1): 43-49. [ Links ]
BUSATO, G. R.; GRUZTMACHER, A. D.; DE OLIVEIRA, A. C.; VIEIRA, E. A.; ZIMMER, P. A.; KOPP, M. M.; DE BANDEIRA, J.; MAGALAHES, T. 2004. Analise da estrutura e diversidade molecular de poblacioes de Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) asociadas as culturas de milho e arroz no Rio Grande do Sul. Neotropical Entomology 33: 709-716. [ Links ]
BUSTILLO, A. E.; POSADA, F. J. 1983. Patogenicidad de un aislamiento de Nomuraea rileyi sobre larvas del cogollero del maíz Spodoptera frugiperda. Revista Colombiana de Entomología 12(1): 5-15. [ Links ]
CLARK, P. L.; MOLINA-OCHOA, J.; MARTINELLI, S.; SKODA, S. R..; ISENHOUR, D. I.; LEE, D. J.; KRUMM, J., T.; FOSTER, J. E. 2007. Population variation of the fall armyworm, Spodoptera frugiperda, in western hemisphere. Journal of Insect Science 7: 05. [ Links ]
DRES, M.; MALLET, J. 2002. Host races in plant-feeding insects and their importance sympatric speciation. Philosophical Transactions of the Royal Society of Science 357: 471-492. [ Links ]
GARCÍA, F.; MOSQUERA, M. T.; VARGAS, C.; ROJAS, L. A 2002. Control biológico, microbiológico y físico de Spodoptera frugiperda (Lepidoptera: Noctuidae) plaga del maíz y otros cultivos en Colombia. Revista Colombiana de Entomología 28 (1): 53-60. [ Links ]
GENSTAT. 2003. GENSTAT 5.0 Release 4.23DE, Lawes Agricultural Trust, Rothamsted Experimental Station. [ Links ]
LEVY, C. H.; GARCÍA-MARUNIAK, A.; MARUNIAK, J. 2002. Strain identification of Spodoptera frugiperda (Lepidoptera: Noctuidae) insects and cell line: PCR-RFLP of cytocrome oxidase c subunit I gene. Florida Entomologist 85 (1): 186-190. [ Links ]
LÓPEZ-EDWARDS, M.; HERNÁNDEZ-MENDOZA, J. L.; PESCADOR- RUBIO, A.; MOLINA-OCHOA, J.; LEZMAGUTIERREZ, R.; HAMM, J. J.; WISEMAN, B. R. 1999. Biological differences between five populations of fall armyworm (Lepidoptera: Noctuidae) collected from corn in Mexico. Florida Entomologist 82 (2): 254-262. [ Links ]
LU, Y. J.; KOCHERT, G. D.; ISENHOUR, D. J.; ADANG, M. J. 1994. Molecular characterization of a strain-specific repeated DNA sequence in the fall armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae). Insect Molecular Biology 3: 123–130. [ Links ]
MARTINELLI, S.; CLARK, P. L.; ZUCCHI, M.I.; SILVAFILHO, M.C.; FOSTER, J. E. OMOTO, C. 2007. Genetic structure and molecular variability of Spodoptera frugiperda (Lepidoptera: Noctuidae) collected in maize and cotton field in Brazil. Bulletin of Entomological Research 97: 225-231. [ Links ]
MCMICHAEL, M.; PASHLEY, D. P. 1999. Differences in amplified fragment-length polymorphism in fall armyworm (Lepidoptera: Noctuidae) host strains. Annals of the Entomological Society of America 92 (2): 175-181. [ Links ]
MEAGHER, R. L.; GALLO-MEAGHER M. 2003. Identifying host strains of fall armyworm (Lepidoptera: Noctuidae) in Florida using mitochondrial markers. Florida Entomologist 86 (4): 450- 455. [ Links ]
NAGOSHI, R. D.; MEAGHER, R. L. 2003a. Fall armyworm FR sequences map to sex chromosomes and their distribution in the world indicate limitations in interstrain mating. Insect Molecular Biology 12 (5): 453-456. [ Links ]
NAGOSHI, R. D.; MEAGHER, R. L. 2003b. FR Tandem-Repeat Sequence in Fall armyworm (Lepidoptera: Noctuidae) Host Strains. Annals of the Entomological Society of America 96 (3): 329-335. [ Links ]
NAGOSHI, R. D.; MEAGHER, R. L. 2004. Behaviour and distribution of the two fall armyworm host strains in Florida. Florida Entomologist 87 (4): 440-448. [ Links ]
NAGOSHI, R. D.; MEAGHER, R. L.: NUESSSLY, G.; HALL, D. G. 2007a. Effects of fall armyworm (Lepidoptera: Noctuidae) interstrain mating in wild populations. Environmental Entomology 35 (2): 561-568. [ Links ]
NAGOSHI, R. D.; SILVIE, P.; MEAGHER, R. L.; LOPEZ, L.; MACHADO, V. 2007b. Identification and comparison of fall armyworm (Lepidoptera: Noctuidae) host strains in Brazil, Texas, and Florida. Annals of the Entomological Society of America 100: 394- 402. [ Links ]
PASHLEY, D. P. 1986. Host-associated genetic differentiation in fall armyworm (Lepidoptera: Noctuidae) a sibling species complex? Annals of Entomological Society of America 79: 898-904. [ Links ]
PASHLEY, D. P.; MARTIN, J. A. 1987. Reproductive incompatibility between host strains of the fall armyworm (Lepidoptera: Noctuidae). Annals of Entomological Society of America 80: 731– 733. [ Links ]
PASHLEY, D. P.; HAMMOND, A. M.; HARDY, T. N. 1992. Reproductive isolating mechanisms in fall armyworm host strains (Lepidoptera: Noctuidae). Annals of Entomological Society of America 85: 400-405. [ Links ]
PASHLEY, D. P. 1998. Sex linkage and speciation in Lepidoptera. pp. 309-319. In Howard, D.; Berlocher, S. (eds). Endless forms: species and speciation. Oxford. NY. [ Links ]
PASHLEY, D. P.; MCMICHAEL M.; SILVAIN J. F. 2004. Multilocus genetic analysis of host use, introgression and speciation in host strains of fall armyworm (Lepidoptera: Noctuidae). Annals of Entomological Society of America 97 (5): 1034-1044. [ Links ]
PINZÓN, G. 1992. Especies sibling de Spodoptera frugiperda. Departamento de desarrollo y registros CIBA-GEYGI COLOMBIANA. 38-41 p [ Links ]
TORRES, L.; COTES, A. M. 2005. Efecto de la crioconservación sobre la viabilidad y actividad biocontroladora de Nomuraea rileyi contra Spodoptera frugiperda (Lepidoptera: Noctuidae). Revista Colombiana de Entomología 31 (2): 133-138. [ Links ]
SAMBROOK, J.; RUSELL, D. 2001. Molecular Cloning. A Laboratory Manual. 3ª. Ed. Cold Spring Harbor Laboratory Press. SOKAL, R. R.; ROLHF, F. J. 1995. Biometry. W. H. Freeman and company, NY. [ Links ]
ZENNER - DE POLANÍA, I.; MADONADO, H. A.; MEJIA CRUZ, R. A.; BAYONA, M. 2007. Susceptibilidad de algunas plagas (Lepidoptera, Noctuidae) del algodonero al gene Cry1ac del Bacillus thuringiensis, pp. 147. En: Resúmenes del XXXIV Congreso Colombiano de Entomología. [ Links ]
Recibido: 5-oct-2007- Aceptado: 25-ago-2008


{kind=link}