










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488On-line version ISSN 2665-4385
Rev. Colomb. Entomol. vol.34 no.2 Bogotá July/Dec. 2008
Diversidad de invertebrados acuáticos asociados a Bromeliaceae en un bosque de montaña
Diversity of aquatic invertebrates associated to Bromeliaceae in the mountain cloud forest
FABIOLA OSPINA-BAUTISTA1, JAIME V. ESTÉVEZ VARÓN2, EMILIO REALPE3 y FERNANDO GAST4
1 Bióloga, M. Sc. Investigadora asociada al Laboratorio de Zoología y Ecología Acuática-LAZOEA, Universidad de Los Andes, Apartado 4976, Bogotá, Docente Universidad Católica de Manizales. Colombia. fospinab@hotmail.com. Autor para correspondencia.
2 M. Sc. Departamento de Ciencias Biológicas, Universidad de Caldas. Manizales, Colombia. jvestevezv@hotmail.com.
3 Biólogo, M. Sc. Laboratorio de Zoología y Ecología Acuática-LAZOEA, Universidad de Los Andes, Apartado 4976, Bogotá, Colombia. erealpe@uniandes.edu.co.
4 Biólogo, Ph. D. Director Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, Bogotá, Colombia. fgast@humboldt.org.co.
Resumen: Se estudiaron los invertebrados acuáticos asociados a los depósitos de agua de Tillandsia turneri y T. complanata (Bromeliaceae) en un bosque de montaña con el fin de determinar diferencias en la comunidad de invertebrados y relacionarlas con las variables morfología y fisicoquímicos de la bromelia. Se muestrearon 19 individuos de cada especie de Bromeliaceae, y a cada uno se le estableció la comunidad de invertebrados acuáticos, el contenido de agua, de hojarasca y variables morfológicas y fisicoquímicas. Se encontraron 23 morfoespecies, dentro de las cuales el orden Diptera fue el más diverso y abundante; algunos géneros estuvieron presentes en solo una de las especies de Bromeliaceae. La comunidad de invertebrados acuáticos fue similar entre las dos bromelias. Para T. complanata las medidas del ancho de la hoja y contenido de hojarasca se relacionan con el número de especies. Para T. turneri la riqueza de Margalef fue explicada por un modelo que incluye las variables numero de hojas, ancho de la vaina y contenido de agua.
Palabras clave: Bosques de niebla. fitotelmata. Tillandsia turneri. Tillandsia complanata.
Abstract: The aquatic invertebrate community associated with the water tank bromeliads Tillandsia turneri and T. complanata was studied in a mountain cloud forest to determine differences in the invertebrate community and relate those to morphological and physicochemical parameters of the bromeliads. Nineteen individuals of each Bromeliaceae species were sampled, and for each one the aquatic invertebrate community, the water content, litter amount and the morphological and physicochemical parameters were assessed. A total of 23 invertebrate morphospecies were found, of which the order Diptera was the most diverse and abundant; some genera were only found in one of the Bromeliaceae species. The aquatic invertebrate community was similar between the two species. For T. complanata, leaf width and litter amount was related to species number. For T. turneri, the Margalef richness was explained by a model that included the variables of number of leaves, pod width and water content.
Key words: Mountain cloud forests. Phytotelmata. Tillandsia turneri. Tillandsia complanata.
Introducción
Las fitotelmata son pequeños cuerpos de agua que provienen de secreciones líquidas o precipitaciones y que se reservan temporal o permanente en plantas o partes de ellas como tallos, ejes de hojas, frutos o flores (Maguire 1971; Machado- Allison et al. 1986; Greeney 2001). En el caso de las bromelias la formación de estanques se ve facilitada por la disposición de sus hojas en roseta, lo cual permite la reserva agua y detritus (Benzing 1990).
La cantidad de agua y hojarasca que intercepta la bromelia varía con la forma y el tamaño de la planta, así a mayor tamaño de la roseta mayor es la capacidad para recibir el agua proveniente de la lluvia y la hojarasca del dosel (Benzing 1990; Richardson 1999; Zotz y Vera 1999; Ospina-Bautista et al. 2004). Lo anterior, implica más espacio para colonización y recursos más diversos (Lawton 1986; Yanoviak 2001) esto influye en la abundancia, riqueza y diversidad de invertebrados asociados a las bromelias (Richardson 1999; Yanoviak 1999a, b).
El área de la roseta de la bromelia influye indirectamente sobre las condiciones fisicoquímicas del agua ya que modifica las tasas de evaporación y las concentraciones de las sales disueltas y estas variables a su vez influyen sobre la conductividad y el pH. El área y su relación con la cantidad y calidad de hojarasca presentes en la fitotelma influye sobre el pH, concentración de oxigeno y dióxido de carbono disuelto en el agua (Laessle 1961). Esos efectos crean cambios en la riqueza y abundancia de fauna asociada a la bromelia (Frank et al. 1976; Warren y Spencer 1996; Sota 1996, 1998; Richardson et al. 2000a).
Adicionalmente, la riqueza de especies puede estar asociada a la complejidad estructural, es así como la riqueza aumenta a medida que la bromelia se hace más compleja (Machado- Allison et al. 1985). Para establecer la complejidad se puede utilizar el número de hojas y su disposición, porque mientras más hojas el número de compartimentos disponibles para la colonización aumenta (Laessle 1961; Frank 1983) y en cuanto a la disposición, en bromelias con cavidades separadas en la base de cada hoja de la roseta, la complejidad es mayor, mientras que bromelias donde existe una sola cámara central constituida por una espiral de hojas fuertemente imbricada y dispuestas verticalmente la complejidad es menor (Benzing 1980, 1990; Machado-Allison et al. 1985).
En los estudios de fauna asociada a bromelias se han usado indistintamente varias especies de estas plantas sin tener en cuenta las variaciones en el tamaño y complejidad estructural (Laessle 1961; Machado-Allison et al. 1986; Fragoso y Rojas-Fernández 1996; Reid y Janetzky 1996; Richardson 1999). Sin embargo, en pocos estudios se han encontrado que éstas y los factores fisicoquímicos del agua puedan afectar la riqueza y diversidad de invertebrados (Armbruster et al. 2002; Araujo et al. 2007). Conocer los patrones que influyen sobre la estructura de una comunidad determinada es un importante propósito ecológico ya que tiene implicaciones para la conservación de la diversidad debido a las altas tasas de destrucción de hábitats (Bowles et al. 1998).
El objetivo de este trabajo es comparar la comunidad de invertebrados acuáticos asociados a dos especies de Bromeliaceae Tillandsia turneri (Baker, 1888) y Tillandsia complanata (Benth, 1846), relacionar las variables morfológicas y fisicoquímicas del agua de cada especie de bromelia con su comunidad de invertebrados, y además, comparar las variables morfométricas como contenido de hojarasca, contenido de agua, parámetros fisicoquímicos entre las dos especies.
Materiales y Métodos
Área de estudio. El estudio se realizó en un bosque de montaña ubicado en el municipio de Guasca, Cundinamarca, Colombia (4º46’N, 74º10’W), a 3.100 m de altitud. El régimen de precipitación es bimodal con dos épocas de lluvia comprendidas entre los meses de marzo a mayo y octubre y noviembre. La precipitación media anual es de 778 mm, la evaporación media anual de 565 mm y la temperatura media mensual es de 12,3ºC. El bosque presenta un dosel dominado por Weinmannia sp. de 10 m de alto. Los árboles están densamente cubiertos por epifitos y el suelo presenta un manto de musgo continuo donde crece gran diversidad de especies herbáceas (Jaramillo y Cavelier 1998).
Trabajo de campo. El estudio se realizó entre diciembre de 2001 y febrero de 2002, durante los cuales se tomaron muestras pareadas en sitios sombreados de 17 bromelias T. turneri y 17 de T. complanata por ser las especies más abundantes en el lugar de estudio, éstas se encontraban entre 0,5 y 3 m de altura asociadas a árboles de Weinmannia sp. A cada individuo se le realizaron mediciones morfométricas como número de hojas, altura de la roseta, cobertura, área y volumen de la roseta calculada a partir de las fórmulas del área y volumen de un cono. Además, se determinó el largo y el ancho de la lámina y de la vaina de las cinco hojas más grandes con el fin de determinar el tamaño y la capacidad de retención de agua.
De cada bromelia se midió la cantidad de agua retenida y se determinó el pH, la conductividad, la temperatura y la concentración de oxígeno; estas medidas se realizaron en la parte interna y externa de la roseta. El pH se determinó a partir de un medidor de pH marca Shott, la conductividad con un medidor de conductividad Extech y el oxígeno por medio de un medidor de oxígeno Hach La hojarasca retenida por los individuos de T. turneri y T. complanata fue recolectada, secada a 60ºC y posteriormente pesada.
Para recolectar los invertebrados acuáticos asociados a loas dos especies de bromelias se deshojó la planta, se lavaron sus hojas con agua destilada y se filtró el resultante con papel de 80 g, Este tipo de muestreo ha sido utilizado para evaluar la comunidad invertebrados en otros estudios (Richardson et al. 2000 a, b; Mestre et al. 2001; Stuntz 2002), tiene como ventaja que permite recolectar todos los individuos asociados a la bromelia, principalmente los estados larvales, además, las poblaciones de bromelias no se ven afectadas por este tipo de muestreo ya que presentan altas densidades en los bosques de montaña (Isaza et al. 2004) lo cual permite que se incremente la proporción de propágulos y por lo tanto los potenciales colonizadores (Yeaton y Gladstone 1982). Se identificaron los invertebrados utilizando las claves de Ward y Whipple (1959), Pennak (1978), McCafferty (1982), Epler (1996), Consoli y Lourenço (1998) y Merritt y Cummins (2007).
Análisis de datos. Se utilizó el número de individuos de las especies de invertebrados presentes en cada planta para calcular el índice de diversidad de Shannon y el índice de riqueza de Margalef y se estableció la abundancia relativa de invertebrados por individuo de T. turneri y T. complanata.
Para establecer si existe relación entre las variables morfológicas, contenido de hojarasca, cantidad de agua y parámetros fisicoquímicos y la comunidad de invertebrados asociados, se realizó una regresión múltiple entre dichas variables y la riqueza, la diversidad y la raíz cuadrada de la abundancia de invertebrados más 0,5 para cada especie de bromelia.
Para determinar si había diferencia entre T. turneri y T. complanata en cuanto a su diversidad, riqueza y abundancia relativa de invertebrados por individuo de bromelia, como también entre las variables morfométricas, fisicoquímicos, contenido de agua, hojarasca entre las dos especies de bromelias se realizaron pruebas de t-student previa comprobación de homogeneidad de varianzas y distribución normal.
Resultados
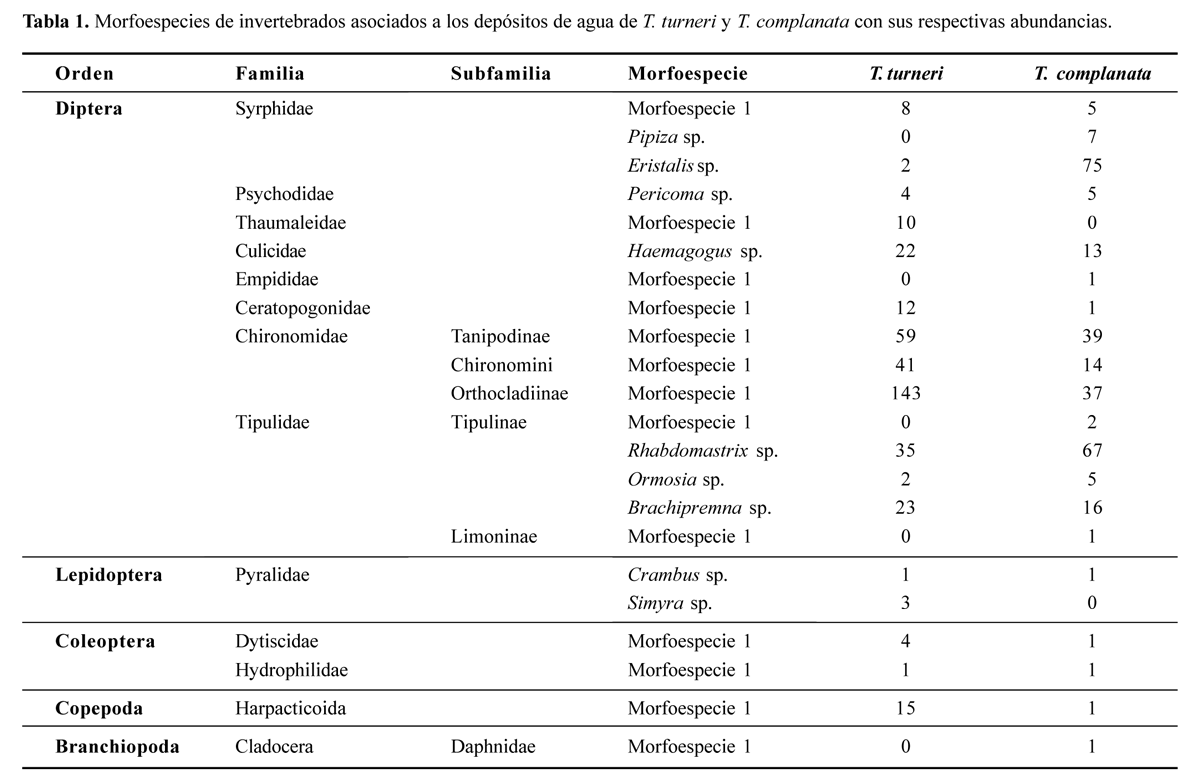
Se encontraron 23 morfoespecies de Arthropoda dentro de las cuales el orden Diptera fue el más diverso con 15 morfoespecies y el más abundante (93,7 % de los individuos en T. turneri, 98,3% en T. complanata). Las morfoespecies de la familia Chironomidae son las más abundantes en T. turneri y las morfoespecies Eristalis sp. y Rhabdomastrix sp. para T. complanata. Además, algunas morfoespecies se encontraron solo en una de las dos especies o en alta proporción en alguna de las bromelias (Tabla 1).
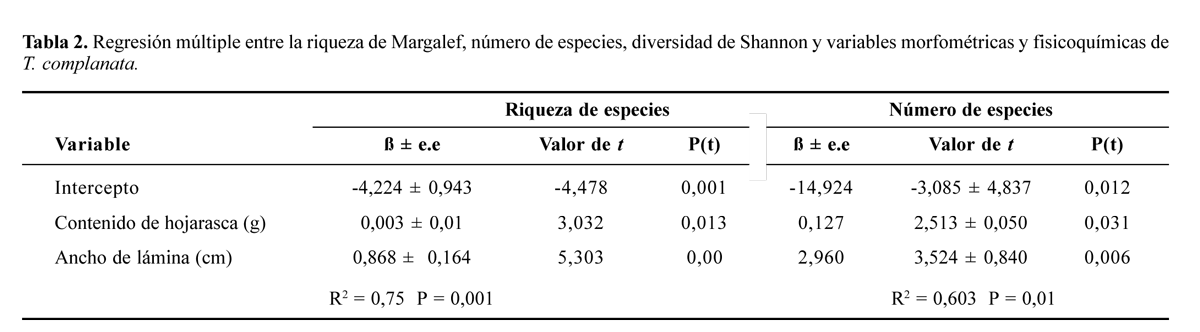
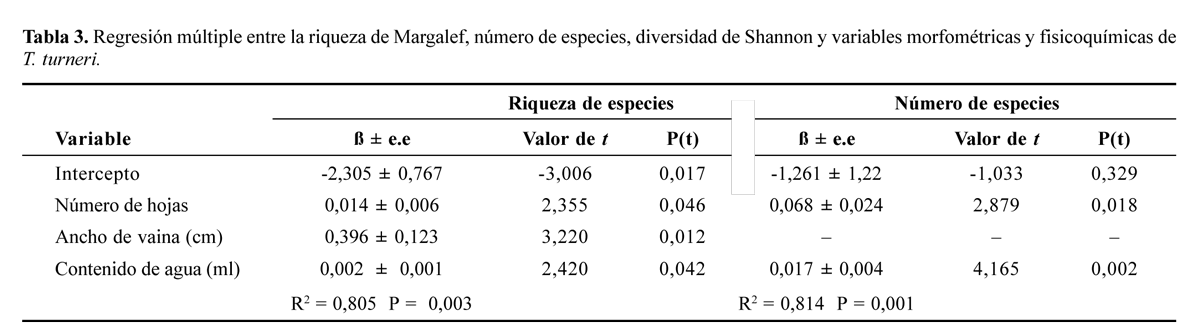
Las variables que mejor describen la relación entre riqueza y los parámetros morfometricos, fisicoquímicos, contenido de agua y hojarasca en T. complanata es el ancho de la hoja y contenido de hojarasca, mientras que no se encontró un modelo que explique la diversidad y la abundancia de invertebrados (Tabla 2). Para T. turneri la riqueza de Margalef es explicada por un modelo que incluye las variables número de hojas, ancho de la vaina y contenido de agua; el número de especies es explicado por el contenido de agua y número de hojas (Tabla 3).
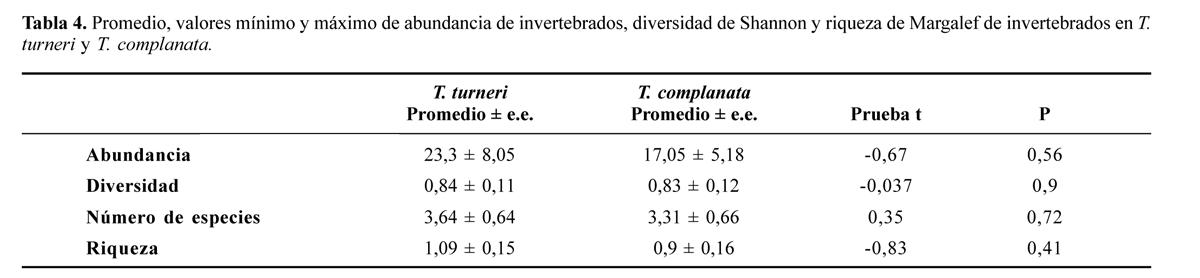
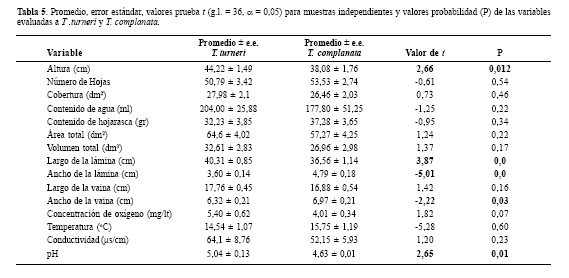
No se encontraron diferencias significativas entre las dos bromelias en cuanto a abundancia, diversidad de Shannon, número de especies y riqueza de Margalef de invertebrados acuáticos asociados (Tabla 4) pero si se encontraron diferencias significativas en las medidas de pH del agua, altura de la roseta, ancho de lámina y vaina y largo de la lámina entre los individuos de las dos especies de Bromeliaceae. T. complanata presenta rosetas con anchos de lámina y vaina mayores que T. turneri, por el contrario las medidas de altura de la roseta y largo de la hoja presentan valores más altos en T. turneri (Tabla 5).
DiscusiónEn las dos especies de bromelias los Artrópodos fueron los más abundantes y diversos, especialmente el orden Diptera; estos resultados son similares a los encontrados para T. turneri en un bosque de montaña de la cordillera oriental colombiana (Ospina- Bautista et al. 2004) y en trabajos realizados en otras especies de bromelias como los de Laessle (1961), Reid y Janetzky (1996), Richarson (1999), Richardson et al. (2000a), Araujo et al. (2007), como los resultados en huecos de árboles Yanoviak (2001).
Los dípteros poseen una alta capacidad de dispersión por el vuelo y de ovoposición en el hábitat donde se desarrollará la larva, estas condiciones permiten que sobrevivan en ecosistemas con condiciones extremas (e.g. bromelia); así para algunos géneros de Diptera existe una relación especie especifica con fitotelmata donde ellos usan estos reservorios naturales como sitios de reproducción (Judd 1998), esto explicaría la gran abundancia y diversidad de los dípteros en T. turneri y T. complanata. La presencia de larvas de Culicidae se puede explicar por la alta riqueza de microorganismos y de detritus en los reservorios de las bromelias (Barrera 1996).
La comunidad de invertebrados acuáticos es similar entre T. turneri y T. complanata, debido a las diferencias en variables que influyen en la riqueza, abundancia y número de especies de invertebrados, según los estudios realizados en otras bromelias (Richarson 1999; Araujo et al. 2007). Por ejemplo, el área y volumen de la planta son medidas relacionadas con la capacidad de la planta de almacenar agua y hojarasca proveniente del dosel; la interacción entre altura de la planta y diámetro podría indicar el espacio posible para colonización (Lawton 1986; Richardson 1999; Yanoviak 2001; Araujo 2007). Adicionalmente, las dos especies de bromelias son similares en cuanto a la disposición de las hojas y forman pequeños reservorios para almacenar agua y detritus, así los procesos de competencia y depredación serían similares.
La presencia de hojas anchas en la roseta de la bromelia permite una mayor cantidad de hojarasca y de espacio para colonización, lo cual resulta en un aumento de la longitud de la cadena trófica; estas características explicarían la relación entre el número y la riqueza de especies con el ancho de la hoja y contenido de hojarasca en T. complanata; la hojarasca es una medida de la productividad del microcosmos, por lo cual a mayor cantidad de hojarasca se esperaría una mayor riqueza especialmente en las cadenas tróficas basadas en detritus. La relación entre esta variable y el número de especies es similar a la encontrada por Ambruster (2001) para especies de los géneros Guzmania, Streptocali y Tillandsia.
El número de hojas es una medida de la complejidad de un hábitat porque indica el número de compartimentos disponibles para colonización y la heterogeneidad del microecosistema (Armbruster et al. 2001); esa característica puede crear cambios en la densidad y diversidad de especies (Machado-Allison et al. 1985; Schwartz et al. 2000) al crear nuevos nichos para la especies o reducir las tasas de competencia (Young 2001) y aumentar los organismos detritívoros al reducir la posibilidad de depredación de éstos (Srivastava 2006). Por lo anterior se esperaría una relación entre el número de hojas y las variables del enjambre como la encontrada en T. turneri. Adicionalmente, en esta especie de bromelia el contenido de agua presentó una relación con riqueza de especies, lo cual se explica porque el agua constituye un recurso importante para los organismos que utilizan las bromelias ya sea temporal o permanentemente.
Las diferencias en los valores de pH entre las dos especies de bromelias se pueden explicar por los procesos de descomposición de la materia orgánica acumulada, que implican la liberación de ácidos orgánicos y dióxido de carbono (Wetzel 2001). Las diferencias a nivel de medidas morfológicas de las bromelias tales como altura de la planta y largo de la hoja no se relacionan con la comunidad, es decir que no presentan un efecto significativo sobre la riqueza, diversidad, número de especies y abundancia.
Conclusiones
La morfología de la bromelia puede influir en la comunidad de invertebrados al afectar las variables microclimáticas y los recursos importantes para la comunidad como el detritus, contenido de agua y espacio disponible para colonización, así, en bromelias con morfología parecida la comunidad de invertebrados seria similar si estas plantas se encuentran dentro del mismo ecosistema. Además, para los invertebrados acuáticos existen variables morfológicas importantes como el ancho de la hoja y vaina, en comparación con los invertebrados terrestres donde influye el diámetro y la altura de la bromelia.
En la comunidad también puede influir la competencia intra e interespecifica (Bradshaw y Holzapfel 1983; Fincke 1992), que para el caso de las dos especies estudiadas seria similar debido a que ambas presentan un suministro por pulso donde el espacio y el recurso alimenticio pueden ser limitantes. Las tasas de inmigración y extinción de invertebrados pueden afectar la composición ya que éstas van a depender de la ubicación espacial de la bromelia dentro del bosque, del sitio de origen de propágulos, de los mecanismos de dispersión de cada especie de invertebrado y de los recursos disponibles (Srivastava y Lawton 1998; Yanoviak 1999b) en especial de la hojarasca proveniente del dosel, para el caso de las dos especies de Bromeliaceae estudiadas la fuente principal de hojarasca era la especie de encenillo (Weinmannia sp.).
Agradecimientos
Los autores expresan sus agradecimientos al Instituto de Investigación de Recursos Biológicos Alexander von Humboldt y al programa de Jóvenes Investigadores de COLCIENCIAS por la financiación de este trabajo. A la Universidad de Los Andes por facilitar la realización del presente estudio. Al Centro de Innovación y Desarrollo Tecnológico de la Universidad de los Andes (CITEC) por facilitar los equipos para medir los parámetros fisicoquímicos.
Literatura citada
ARAÚJO, V. A., MELO, S. K., ARAÚJO, A. P. A., GOMES, M. L. M., CARNEIRO, M. A. A. 2007. Relationship between invertebrate fauna and bromeliad size. Brazilian Journal of Biology 67 (4): 611-617. [ Links ]
ARMBRUSTER, P.; HUTCHINSON, R. A.; COTGREAVE, 2002. Factors influencing community structure in South America tank bromeliad fauna 3 Oikos 96: 225-234. [ Links ]
BARRERA, R. 1996. Competition and resistance to starvation in larvae of container-inhabiting Aedes mosquitoes. Ecological Entomology 21: 117-127. [ Links ]
BRADSHAW, W. E.; HOLZAPFEL. C. M. 1983. Predator-mediated, non-equilibrium coexistence of tree-hole mosquitoes in southeastern North America. Oecologia 57: 239-256. [ Links ]
BENZING, H.D. 1980. The biology of bromeliads. Mad River Press, Eureka, California. 287 p. [ Links ]
BENZING, H. D. 1990. Vascular epiphytes. General biology and related biota. Cambridge University Press, Cambridge. 354 p. [ Links ]
BOWLES, I. A.; RICE R. E.; MITTERMEIR R. A., FONSECA G.A.1998. Logging and tropical forest conservation. Science 280: 1899-1900. [ Links ]
CONSOLI, R. A. G. B..; LOURENÇO DE OLIVERA, R. 1998. Principais mosquitos de importância sanitária no Brasil. 1ª Reimpressão. Fiocruz. Rio de Janeiro. 228 p. [ Links ]
EPLER, J. H. 1996. Identification manual for the water beetles of Florida. Department of environmental protection. Division of water facilities. Tallahasse. 259 p. [ Links ]
FINCKE, O. M. 1992. Interspecific competition for tree holes: consequences for mating systems and coexistence in neotropical damselflies. American Naturalist 139: 80-101. [ Links ]
FRAGOSO, C.; ROJAS-FERNÁNDEZ, P. 1996. Earthworms inhabiting bromeliads in Mexican tropical rainforests: ecological and historical determinants. Journal of Tropical Ecology 12: 729-734. [ Links ]
FRANK, J. H.; CURTIS, G. A.; EVANS, H. J. 1976. On the bionomics of bromeliad-inhabiting mosquitoes. II. The relationship of bormeliad size to the number of immature Wyeomyia vanduzeei and Wy. medioalbipes. Mosquito News 37: 180-192. [ Links ]
FRANK, J. H. 1983. Bromeliad phytotelmata and their biota, especially mosquitos, pp.101-128. En: Frank, J. H.; Lounibos, P. L. (eds.) Phytotelmata: terrestrial plants as hosts of aquatic insects communities. Plexus Inc., New Jersey. 293 p. [ Links ]
GREENEY, H. F. 2001. The insects of plant-held waters: a review and bibliography. Journal of Tropical Ecology 17: 241-260. [ Links ]
ISAZA, C.; BETANCUR, J.; ESTEVEZ-VARÓN, J. 2004. Vertical distribution of bromeliads in a montane forest in the eastern cordillera of the Colombian andes. Selbyana 25 (1): 126-137. [ Links ]
JARAMILLO, M.; CAVELIER, J. 1998. Fenología de dos especies de Tillandsia (Bromeliaceae) en un bosque montano alto de la cordillera oriental colombiana. Selbyana 19: (1) 44-51. [ Links ]
JUDD, D. D.1998. Review of a bromeliad-ovopositing lineage in Wyeomyia and the resurrection of Hystatomyia (Diptera: Culicidae). Annals of the Entomological Society of America 91: 572-589. [ Links ]
LAESSLE, A. M. 1961. A micro-limnological study of Jamaican bromeliads. Ecology 42: 499-517. [ Links ]
LAWTON, J. H. 1986. Surface availability and insect community structure: the effects of architecture and fractal dimension of plants, pp. 317-331. En: Juniper B. E.; Southwood T. R. E. (eds.). Insects and the plant surface, Edward Arnold, London, England. 360 p. [ Links ]
MACHADO-ALLISON, C. E. R. BARRERA, FRANK, J. H., DELGADO L., GOMEZ-COVA, C. 1985. Mosquito communities in Venezuela phytotelmata, pp. 79-93. En: Lounibos, L. P.; Rey, J. R.; Frank, J. H. (eds.). Ecology of mosquitoes. Vero Beach, Florida. USA. [ Links ]
MACHADO-ALLISON, C. E.; BARRERA, R.; DELGADO, L.; GÓMEZ-COVA, C.; NAVARRO, J. C. 1986. Mosquitos (Diptera: Culicidae) de los fitotelmata de Panaquire, Venezuela. Acta Biológica Venezolana 2 (12): 1-12. [ Links ]
MAGUIRE, B. 1971. Phytotelmata: biota and community structure determination in plant-held waters. Annual Review of Ecology and Systematics 2: 439-464. [ Links ]
McCAFFERTY, W. P. 1982. Aquatic Entomology. Science Books International. Boston. 448 p. [ Links ]
MERRITT, R. W.; CUMMINS, K. W. 2007. An introduction to the aquatic insects of North America. Segunda edición. Kendall/Hunt Dubuque, Iowa.722 p. [ Links ]
MESTRE, L. A.; ARANHA, J. M.; ESPER, M. 2001. Macroinvertebrate fauna associated to the bromeliad Vriesea inflata of the atlantic forest (Paraná State, Southern Brazil). Brazilian Archives of Biology and Technology 44 (1): 89-94. [ Links ]
OSPINA-BAUTISTA, F.; ESTEVEZ, J.V.; BETANCUR J.; REALPE E. 2004. Invertebrados acuáticos asociados a Tillandsia turneri Baker (bromeliaceae) en un bosque altoandino (Cundinamarca, Colombia). Acta Zoológica Mexicana 20 (1): 153-166. [ Links ]
PENNAK, R. W. 1978. Fresh-water invertebrates of United States. Second edition. John Wiley & Sons, New York. 803 p. [ Links ]
REID, J. W.; JANETZKY, W. 1996. Colonization of Jamaican bromeliads by Tropocyclops jamaicensis n. sp. (Crustacea: Copepoda: Cyclopoida). Invertebrate Biology 115 (4): 305-320. [ Links ]
RICHARDSON, B. A. 1999. The bromeliad microcosm and the assessment of fauna diversity in a Neotropical forest. Biotropica 31 (2): 321-336. [ Links ]
RICHARDSON, B.; ROGERS, C.; RICHARDSON, M. J. 2000a. Nutrients, diversity and community structure of two phytotelm systems in a lower mountain forest, Puerto Rico. Ecological Entomology 25: 348-356. [ Links ]
RICHARDSON, B.; RICHARDSON M. J.; SCATENA F. N.; McDOWELL W. H. 2000b. Effects of nutrient availability and other elevation changes on bromeliad populations and their invertebrate communities in a humid tropical forest in Puerto Rico. Journal of Tropical Ecology 16 (2): 167-188. [ Links ]
SCHWARTZ, M. W.; BRIGHAM, C. A.; HOEKSEMA, J. D.; LYONS, K. G.; VANMANTGEM, P. J. 2000. Linking biodiversity to ecosystem function: implications for conservation ecology. Oecologia 122: 297-305. [ Links ]
SOTA, T. 1996. Effects of capacity on resource input and the aquatic metazoan community structure in phytotelm. Researches on Population Ecology. 38: 65-73. [ Links ]
SOTA, T. 1998. Microhabitat size distribution affects local difference in community structure: metazoan community in treeholes. Researches on Population Ecology 40: 249-255. [ Links ]
SRIVASTAVA, D. S.; LAWTON. H. 1998. Why more productive sites have more species: experimental test of theory using treehole communities. American Naturalist 152: 510-529. [ Links ]
SRIVASTAVA D. S. 2006. Habitat structure, trophic structure and ecosystem function: interactive effects in a bromeliad–insect community. Oecologia 149: 493-504. [ Links ]
STUNTZ S.; ZIEGLER C.; SIMON U.; ZOTZ G. 2002. Diversity and structure of the arthropod fauna within three canopy epiphyte species in central Panama. Journal of Tropical Ecology 18: 161- 176. [ Links ]
WARD, H. B; WHIPPLE, G. C. 1959. Fresh water biology. Second edition. W. T. Edmondson. New York. [ Links ]
WARREN, P. H; SPENCER, M. 1996. Community and food-web responses to the manipulation of energy input and disturbance in small ponds. Oikos 75: 407-418. [ Links ]
WETZEL, R. G. 2001. Limnology: lake and river ecosystems. 3rd ed. Academic Press, California. 1006 p. [ Links ]
YANOVIAK, S. P. 1999a. Community structure in water-Filler tree holes in Panama: effects of hole height and size. Selbyana 20: 106-115. [ Links ]
YANOVIAK, S. P. 1999b. Effects of leaf litter species on macroinvertebrate community properties and mosquito yield in neotropical tree hole microcosms. Oecologia 120: 265-271. [ Links ]
YANOVIAK, S. P. 2001. The macrofauna of water- filled tree holes on Barro Colorado island, Panama. Biotropica 33 (1): 110-120. [ Links ]
YEATON, R. I.; GLADSTONE, D. E. 1982. The pattern of colonization of epiphytes on calabash tress (Crescentia alata HBK) in Guanacaste Province, Costa Rica. Biotropica 14: 137- 140. [ Links ]
YOUNG, K. 2001. Habitat diversity and species diversity: testing the competition hypothesis with juvenile salmonids. Oikos 95: 87-93. [ Links ]
ZOTZ, G.; VERA, T. 1999. How much water is in the tank? Model calculations for two epiphytic bromeliads. Annals of Botany 83: 183-192. [ Links ]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}