










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488On-line version ISSN 2665-4385
Rev. Colomb. Entomol. vol.35 no.1 Bogotá Jan./June 2009
Spodoptera frugiperda: respuesta de distintas poblaciones a la toxina Cry1Ab
Spodoptera frugiperda: response of different populations to the Cry1Ab toxin
INGEBORG ZENNER DE POLANÍA 1, HELBER ADRIAN ARÉVALO MALDONADO 2, RODOLFO MEJÍA CRUZ 3 y JULIÁN L. DÍAZ SÁNCHEZ 4
1 Ph. D. Facultad de Ingeniería Agronómica, Universidad de Ciencias Aplicadas y Ambientales U.D.C.A, Calle 222 No. 54-37, Bogotá, D. C., izenner@udca.edu.co. Autor para correspondencia.
2 Estudiante Facultad de Ingeniería Agronómica, Universidad de Ciencias Aplicadas y Ambientales U.D.C.A, Calle 222 No. 54-37, Bogotá, D. C. jelber2000@gmail.com.
3 M. Sc. Facultad de Ingeniería Agronómica, Universidad de Ciencias Aplicadas y Ambientales U.D.C.A, Calle 222 No. 54-37, Bogotá, D. C. rmejia@udca.edu.co.
4 Estudiante Facultad de Ingeniería Agronómica, Universidad de Ciencias Aplicadas y Ambientales U.D.C.A, Calle 222 No. 54-37, Bogotá, D. C. julianrudeboy86@hotmail.com.
Resumen: Spodoptera frugiperda es la plaga insectil que ocasiona las mayores pérdidas a la producción de maíz en el trópico y neo-trópico. Su control en Colombia, ocupa cerca del 10% de los costos de producción. Plantas transgénicas con la toxina Cry1Ab son propuestas para mermar las pérdidas y evitar el empleo de insecticidas. Para poder manejar la posible resistencia a la toxina y poder monitorear este fenómeno inherente al insecto, se deben realizar estudios relacionados con la determinación de la línea base de susceptibilidad y con el comportamiento de las larvas frente a las plantas transformadas. Esta investigación tuvo como objetivo el establecimiento de la línea base, para diferentes poblaciones, alimentando larvas neonatas con dieta merídica con la toxina incorporada en dosis seriadas, para luego monitorear la susceptibilidad de las larvas al Cry1Ab a través del tiempo. Igualmente, se determinó el efecto del consumo de tejido del híbrido transgénico sobre las larvas. La línea base calculada, para la zona del Tolima correspondió a 5,64ppm y, aquella para una población de la zona cafetera de Cundinamarca a 8,35ppm. A través del tiempo la población del Tolima disminuyó su susceptibilidad, la de Cundinamarca, no sujeta a presión de selección, no varió mayormente. Un 96,66% de mortalidad sufrieron larvas neonatas alimentadas con tejido fresco, follaje verde del maíz transgénico, mientras en aquellas alimentadas con follaje amarillo el porcentaje de mortalidad correspondió a 76,59%. Se concluye que el cogollero del maíz muestra una tendencia marcada de adquirir tolerancia al Cry1Ab en corto tiempo.
Palabras clave: Cogollero del maíz. Línea base de susceptibilidad. MON810. Clorofila. Monitoreo de resistencia.
Abstract: Spodoptera frugiperda is the insect pest that causes the highest losses to corn production in the Tropics and Neotropics. Its control in Colombia accounts for at least 10% of production costs. Transgenic plants with the Cry1Ab toxin are proposed to decrease losses and avoid the use of insecticides. In order to manage possible resistance to the toxin and to monitor the phenomenon inherent in this insect, studies should be conducted on its baseline susceptibility and on larval behavior on transformed plants. The objective of this research was to establish the baseline for different populations, feeding neonate larvae with a meridic diet with a serial dosage of incorporated toxin to monitor the susceptibility of larvae to Cry1Ab through time. In addition, the effect of the consumption of tissue from the transgenic hybrid by larvae was determined. The baseline calculated for the zone of Tolima corresponded to 5.64ppm, while that of the coffee growing zone of Cundinamarca was 8.35ppm. Through time the Tolima population decreased its susceptibility, while that of Cundinamarca, not subjected to selection pressure did not vary considerably. Neonate larvae suffered 96.66% mortality when fed fresh tissue, green foliage of the transgenic hybrid, while those fed with yellow foliage showed a mortality of 76.59%. It was concluded that the fall armyworm shows a marked tendency to acquire tolerance to Cry1Ab within a short time.
Key words: Fall armyworm. Baseline susceptibility. MON810. Chlorophyll. Resistance monitoring.
Introducción
El cogollero del maíz, Spodoptera frugiperda (J. E. Smith, 1797) (Lepidoptera: Noctuidae), insecto nativo del trópico y subtrópico americano es considerado una de las plagas insectiles omnívoras de mayor importancia económica en el sur de Estados Unidos, Centro y Sur América, destacándose entre sus huéspedes el maíz (Mitchell, 1979, citado por Malo et al. (2001); Westbrook y Sparks 1986; Posada 1989; Virla et al. 1999; Waquil et al. 2002). Las larvas, se alimentan de las hojas nuevas del maíz y pueden causar pérdidas hasta por 400 millones de dólares en el Brasil, por ejemplo (Cruz et al., citados por Vilarinho et al. 2006).
Por la resistencia innata a los pesticidas y la disposición en la planta de esta plaga, el control con productos químicos se dificulta, como lo menciona Ávila et al. (1997) para el Brasil, donde es considerada la principal plaga del maíz, causando pérdidas hasta del 100% en la producción. Polanczyk et al. (2004), mencionan esta misma dificultad de control para los Estados Unidos, México, Cuba y Colombia entre otros países. Su importancia económica y ambiental radica en las pérdidas que ocasiona a los cultivos, los costos de control y la contaminación producida por los agroquímicos empleados.
En Colombia, no existen estadísticas referentes a pérdidas confiables causadas por el insecto, pero en maíz tecnificado, se considera que un 5,6% a un 10% de los costos de producción corresponden al control químico de la plaga (URPA, citado por García Roa et al. 1999). Para el Brasil, se reportan pérdidas del 17 al 38,7% de la producción del maíz, dependiendo del estado fenológico de la planta atacada y de la variedad sembrada (Carvalho 1970 y Williams et al. 1990, citados por Waquil et al. 2004). En Colombia el área tecnificada, cultivada con maíz amarillo, el cual incluye aproximadamente 50.000 ha localizadas en al zona cafetera, ocupó 174.599 ha de acuerdo con FENALCE (2008) para el 2007. En la zona cafetera la siembra intercalada, se recomienda en lotes recién sembrados de café o una vez soqueado el cafetal (Vanegas y Polanía 2002). De la superficie total mencionada, por lo menos, un 50% podría ser potencialmente cultivada con el híbrido de maíz transgénico en los departamentos de Córdoba, Tolima y Meta.
Las siembras de plantas transgénicas que expresan toxinas del Bacillus thuringiensis (Bt) han despertado mucho interés y controversia entre los científicos, los agricultores, los ambientalistas, los ingenieros agrónomos, asistentes técnicos y el público en general. A estos organismos genéticamente modificados se les introdujo mediante metodologías de ingeniería genética ADN foráneo de diferente índole. Las plantas a las cuales se ha introducido por ejemplo el gene Cry1Ab del B. thuringiensis var. kurstaki, el cual expresa cristales proteicos δ-endotoxinas, poseen efectividad insecticida contra larvas de algunas especies del Orden Lepidoptera (Waquil et al. 2002; Tabashnik et al. 2003). El híbrido de maíz transgénico, resistente a insectos plagas, más comúnmente cultivado en las Américas expresa precisamente estos cristales proteicos Cry1Ab (evento MON810) (Koziel et al. 1993, citados por Abel y Adamczyk 2004). La toxina que expresa el transgen se produce en tejidos de la planta transformada en concentraciones variables, lo cual puede alterar el nivel de control de algunas de las especies plagas lepidópteros de este cultivo (Abel y Adamczyk 2004) y por lo tanto a la larga ocasionar el fenómeno de resistencia, debido a la supervivencia de individuos heterocigotos y homocigotos resistentes en tejidos con bajas concentraciones de la toxina.
Para el manejo de la resistencia, se recomienda, la siembra de refugios con maíz convencional, cercanos a los cultivos transgénicos (EPA 1998), con el fin de producir suficientes adultos susceptibles, los cuales puedan evitar, por competencia, la cópula entre individuos resistentes. El éxito de esta estrategia radica en la expresión de la toxina en una alta dosis y efectiva en todos los tejidos afectados por la plaga, para que los individuos heterocigotos de la población no alcancen el estado adulto (Abel y Adamczyk 2004). Estas concentraciones fueron determinadas por los mismos autores, encontrándose diferencias significativas entre porciones verdes, amarillo-verdes y blanco-amarillos, tomadas de la hoja número nueve del híbrido de maíz transgénico. Concluyeron que plantas monocotiledóneas y dicotiledóneas no expresan completamente al Cry1A en tejidos con bajo contenido de clorofila. Por lo tanto, se puede asumir que el cogollo, bajo en clorofila, del híbrido del maíz transgénico, donde se desarrolla la larva de S. frugiperda, por lo menos del tercer instar en adelante, no contiene la suficiente concentración del Cry1Ab para ejercer un control satisfactorio, lo cual implicaría una dosis subletal para las larvas sobrevivientes y aceleraría la adquisición de resistencia a la proteína tóxica.
Los estudios realizados en Colombia sobre resistencia y a nivel mundial con individuos del cogollero del maíz procedentes del neotrópico a productos utilizados en su control son escasos. Las primeras evaluaciones, se realizaron en el país con poblaciones de S. frugiperda procedentes de diversas zonas maiceras de los departamentos del Tolima y del César, detectándose resistencia en algunas poblaciones a los insecticidas metomil, clorpirifos, cipermetrina y una menor susceptibilidad de una población del Tolima, en comparación con otras, donde no se había aplicado el entomopatógeno, a una formulación comercial del Bt, lo cual, los autores recomendaron tomar como una advertencia de la posibilidad de adquirir resistencia del insecto a la bacteria (Zenner de Polanía y Borrero Fonseca 1994). La presión de selección ejercida durante cuatro generaciones en el laboratorio, sobre una población de S. frugiperda procedente de Anolaima (Cundinamarca) con una formulación comercial de Bt (mezcla de cinco toxinas, incluyendo Cry1Ac y Cry1Ab), mostró un incremento de 4,6 veces en la CL50 (Borrero y Zenner de Polanía 1998). Posteriormente, Zenner de Polanía et al. (2005) demostraron la ineficiencia de la toxina Cry1Ac sobre poblaciones del cogollero, procedentes del algodonero y de maíz, obteniendo una CL50 similar a la registrada para el insecto con el producto comercial a base de Bt (Zenner et al. 2006; Zenner de Polanía et al.2008).
Bioensayos ejecutados en el Brasil, empleando dietas merídicas con aplicación sobre la superficie de dosis seriadas de la toxina Cry1Ac mostraron un aumento en la tolerancia de la población del cogollero después de selección durante cuatro generaciones (Vilella et al. 2002). Williams et al. (1997) y Williams et al. (1997) estos últimos investigadores citados por Vilella et al. (2002), encontraron niveles significantes de control del S. frugiperda con híbridos de maíz que expresan la toxina Cry1Ab, pero observaron también supervivencias de algunas larvas. Otro estudio, realizado por Waquil et al. (2004), empleando el método de exposición de larvas del cogollero a dieta artificial con superficie tratada con la toxina, mostró que al transferir a los supervivientes a dieta no tratada a los diez días del inicio del ensayo, el 62,8% de ellas recuperó su desarrollo y logró su biomasa normal. Los investigadores infieren implicaciones relevantes para el desarrollo de resistencia al Cry1Ab del S. frugiperda. Investigaciones ejecutadas por Fernándes et al. (2003) realizando evaluaciones en dos estaciones experimentales de Monsanto en el Brasil, concluyeron que el maíz híbrido MON 810 (Yieldgard®) fue eficiente en el control del cogollero del maíz, mientras que Abel y Pollan (2004), evaluando la resistencia de campo de este híbrido Bt de maíz en Stoneville (EEUU), conceptúan que provee una resistencia intermedia al cogollero en comparación con variedades tradicionales. En Colombia, el Instituto Colombiano Agropecuario autorizó la importación y siembra del híbrido de maíz con tecnología Yieldgard® (MON810) mediante la resolución No. 00465 de Febrero 26 del año 2007, la cual menciona que el maíz “contiene el gen Cry 1A (b) que codifica para la proteína insecticida, cry1 A (b) proveniente de la bacteria Bacillus thuringiensis subsp. kurstaki”. La misma resolución asevera que la autorización de las siembras del cultivar transgénico se basa en los estudios de bioseguridad que determinaron que no “hubo efecto negativo de la tecnología Yieldgard® (Mon 810) sobre el agroecosistema donde se desarrollaron los estudios” (ICA 2007a).
Sin embargo, no indica la existencia de estudios relacionados con la línea base de susceptibilidad del S. frugiperda a la toxina del Bt, indispensable para el monitoreo de la resistencia del insecto al Cry, ni menciona otros experimentos publicados que muestran la respuesta de la plaga a diversos tejidos del híbrido transformado.
Antes de proponer y de permitir el uso de cualquier nueva herramienta de control, normalmente traída de países tecnológicamente y científicamente más avanzados en estos contextos que los nuestros, se requieren pruebas que demuestren sus bondades y los beneficios para la agricultura del país. La literatura reporta estudios básicos relacionados con S. frugiperda y el híbrido transgénico Yieldgard® realizados bajo condiciones de otros sistemas agropecuarios en otros países, principalmente Estados Unidos, Brasil y Argentina (Waquil et al. 2002, 2004; Chilcutt et al. 2007; Virla et al. 2008). Por esta razón y, para disponer de resultados con poblaciones colombianas, se evaluaron algunos aspectos básicos relacionados con la determinación de la línea base de susceptibilidad del cogollero a la toxina Cry1Ab, la variación de la concentración letal media a través del tiempo y el efecto sobre las larvas de tejidos con contenidos variados de clorofila. En este estudio, se incluyó una población de la zona marginal cafetera de Anolaima (Cundinamarca), tendiendo en cuenta la factibilidad de la migración de los adultos del cogollero de esta zona a departamentos como el Tolima. La capacidad de migrar largas distancias y vencer obstáculos naturales de S. frugiperda fue comprobada, en otros países por diversos autores utilizando una serie de métodos, tales como la evaluación de la susceptibilidad a insecticidas (Pitre 1988) y a un virus de la polihedrosis nuclear (Fuxa 1987), la captura en trampas cebadas con feromonas y la influencia de la velocidad y dirección del viento (Westbrook y Sparks 1986; Malo et al. 2001; Rojas et al. 2004) y, recientemente, el empleo de haplotipos del insecto para monitorear su migración (Nagoshi et al. 2008).
Materiales y Métodos
Teniendo en cuenta algunas de las zonas donde se siembra maíz y las tendencias de migración del insecto, se escogieron tres áreas para obtener el material biológico: una zona del departamento del Tolima incluida en un triángulo hipotético entre El Espinal (4°08’00”N y 74°54’00”OE), El Guamo (4°02’05”N y 74°58’25”OE) y Gualanday (4°16’54”N y 75°01’29”OE); una sola recolección en el área rural de Villavicencio (Departamento del Meta, 4°09’02”N y 73°39’31”OE) y, en la zona marginal cafetera de Cundinamarca, región rural de Anolaima (4°45’15,3”N y 74°29’46,7”OE).
Para iniciar las crías del insecto de cada procedencia, se colectaron, ya sea posturas o larvas en cultivos de maíz no transgénico, exceptuando las tomas de muestra del año 2008, las cuales se efectuaron en lotes del híbrido Yieldgard®, situado entre El Espinal y El Guamo. La raza de la zona cafetera fue colectada en la zona rural de Anolaima en maíz con cinco hojas desplegadas; se crió con la dieta ICRISAT, debido a que en esta dieta se obtuvieron resultados favorables (Arévalo 2008), durante cuatro generaciones, sin someterlo a presión de selección por el Cry1Ab, determinando la CL50 a la toxina para cada generación. Basado en las evaluaciones de Zenner et al. (2006), quienes recomiendan criar a S. frugiperda procedente del área del Tolima durante una generación en el laboratorio para evitar la influencia de un posible parasitismo por Chelonus insularis (Cresson, 1865) (Hymenoptera: Braconidae, Cheloninae), se permitió su desarrollo hasta la siguiente generación, para luego emplear las larvas neonatas para los diversos bioensayos. Las crías y los bioensayos, se efectuaron en vasos plásticos de capacidad de 40 mL con tapa inyectada, utilizando dieta merídica a base de garbanzo, “ICRISAT diet 3” (comunicación personal Dr. Sharma, ICRISAT, Instituto Internacional de Investigación en Cultivos para las Zonas Tropicales Semiáridas Patancheru, Andhra Pradesh, India). Todos los bioensayos, se llevaron a cabo en el laboratorio de Biotecnología Agrícola de la Universidad de Ciencias Aplicadas y Ambientales U.D.C.A, bajo condiciones de temperatura promedio de 24,9ºC y humedad relativa de 70%.
La respuesta dosis mortalidad, se evaluó incorporando dosis seriadas, 0,001-0,01-0,1-1,0-10,0 μg/mL de la toxina Cry1Ab a la dieta. La toxina, se obtuvo de un gene del Bt, donado por el Bacillus Genetic Stock Center (Ohio State University, EE. UU.), extraído de la cepa ECE-54 de E. coli, mediante la metodología recomendada por Akhurst et al. (2003). Se empleó un diseño de bloques al azar con seis repeticiones, cada una de ellas representada por un vaso plástico con dieta sobre la cual se colaron cinco larvas neonatas. La mortalidad, se estimó a los siete días del inicio de la prueba y los datos, se sometieron a análisis Probit (SAS Institute Inc.) para determinar las concentraciones letales.
Para determinar la influencia del contenido hipotético de clorofila del tejido de las plantas transgénicas del híbrido de maíz, basado en la apreciación de la intensidad del color, sobre el desarrollo de las larvas, se alimentaron 30 larvas neonatas, con tejido “verde” (TV), es decir hojas, y otras 30 con tejido “amarillo” (TA), cogollo y capacho y mazorca tierna. La población para esta evaluación se obtuvo de un lote de maíz convencional de El Espinal. El tejido transgénico respectivo para la alimentación se obtuvo de una siembra de maíz Yieldgard® en un invernadero bajo condiciones de bioseguridad. Durante el desarrollo se proporcionó a las larvas alimento fresco cada tercer o cuarto día. Para evitar el canibalismo, se individualizaron las larvas a los siete días de edad. Ambos grupos tuvieron como testigo un número igual de larvas alimentadas con el tejido respectivo procedente de maíz convencional, cultivado bajo las mismas condiciones que el híbrido transgénico. Se realizó un análisis descriptivo, expresado en la duración del estado larval y en el peso de las pupas sobrevivientes. A los datos, se aplicaron una prueba de t y de z, respectivamente.
Resultados y Discusión
La línea base de susceptibilidad calculada para la población procedente de El Espinal, corresponde a 5,64 μg/mL de dieta aunque como se muestra en la Tabla 1, en el primer semestre de evaluación (2006B) se obtuvo una variación considerable entre las dosis letales de dos generaciones evaluadas. El análisis Probit de ambas generaciones mostró una pendiente considerable, significando una buena susceptibilidad de la población a la toxina: a medida que aumenta la dosis de la proteína Cry1Ab, aumenta la mortalidad larval. Comparando las CL50 a través del tiempo para el cogollero procedente de las tres zonas muy cercanas entre sí del departamento del Tolima, (Tabla 1), se observa un incremento de la tolerancia a la toxina, lo cual podría estar relacionado con el aumento del área sembrada de maíz con tecnología Yieldgard® (datos de área de siembra no disponibles a los investigadores). En el primer semestre del segundo año de evaluación, la primera generación sigue susceptible, la segunda muestra un aumento en la tolerancia a la toxina, la cual sigue similar durante el segundo semestre del mismo año. Teniendo en mente la resistencia innata del S. frugiperda a la toxina Cry1Ac (Zenner de Polanía et al. 2008) y, el aumento de la resistencia al estar expuesto a cultivares que expresan esta toxina como el algodón Bollgard® I y II, se puede sospechar de la existencia de una resistencia cruzada a las toxinas del Bt por el cogollero del maíz, lo cual, se manifestaría en una rápida adquisición y aumento de resistencia también al Cry1Ab.
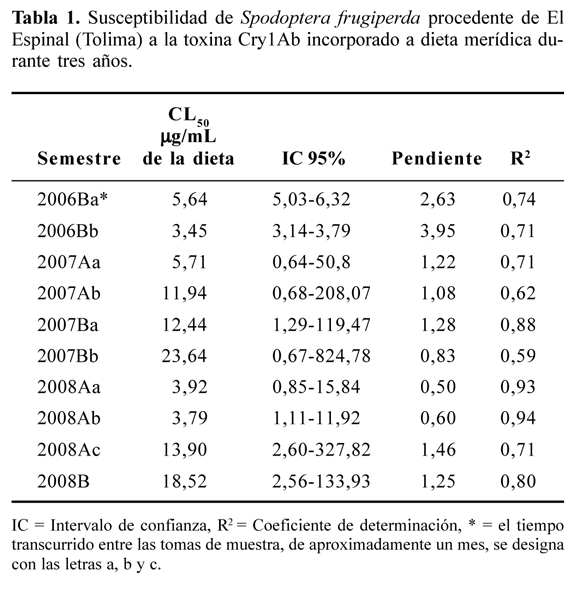
Para el primer semestre del tercer año de análisis (2008Aa y 2008Ab, Tabla 1), se detecta inicialmente una concentración letal media similar a la línea base de susceptibilidad estimada durante el primer semestre de evaluación (2006Ba y 2006Bb); sin embargo 2008Ac, al final del ciclo del cultivo la concentración letal vuelve a subir, de manera similar a lo detectado en el primer semestre del año 2007. En este momento las plantas ya habían producido espigas y las neonatas utilizadas para el bioensayo, provenían de larvas recolectadas precisamente de espigas del híbrido transgénico y criadas hasta la siguiente generación sobre la dieta merídica. Para la primera generación del segundo semestre del año 2008, la CL50 aumenta aún más, ya a proporciones preocupantes e indicadoras de cierta tolerancia a la toxina. En general la variación de la susceptibilidad, se considera errática, 4,19 veces en el 2007Bb y 3,28 para el 2008B (Tabla 1).
La única muestra analizada procedente de maíz convencional del departamento del Meta, arrojó una CL50 de 16,09 μg/mL de dieta, mostrando el análisis probit una pendiente de 1,19, un coeficiente de determinación de 0,76 y un intervalo de confianza, muy amplio, de 1,13 a 228,27. Es probable que la concentración del Cry1Ab determinada en hojas de un cultivo de Yieldgard® de Granada (Meta), 15,37 ppm (datos sin publicar) puedan controlar satisfactoriamente al S. frugiperda de esta zona, la cual se está convirtiendo en un área muy importante para el cultivo de este cereal.
Las larvas, antes de meterse al cogollo se alimentan del follaje, el cual contiene una concentración promedio para el área de El Espinal de la toxina de 8,5 ppm (datos sin publicar), proporcionando por lo tanto un control satisfactorio del primer instar de la plaga. La concentración de la toxina en el follaje del Yieldgard® analizado en el primer semestre del año 2008A en cuyas espigas se recolectaron las larvas vivas mencionadas anteriormente del tercer instar en adelante, mostraron una concentración en hojas de 10,28 ppm (datos sin publicar), adecuada para el control del cogollero, mientras que en las espigas esta concentración del Cry1Ab diminuyó considerablemente y solamente fue de 1,66 ppm (datos sin publicar), dosis demasiado baja para controlar a las larvas del tercer instar en adelante.
Aparentemente, el único estudio realizado para determinar la línea base de susceptibilidad de cuatro poblaciones de S. frugiperda fue el realizado por Lynch et al. (2003) quienes, sin embargo, emplearon otra metodología, la dispersión de la dosis de la toxina sobre la dieta y no incorporada a la dieta y además sometieron larvas de un día de edad y no neonatas. Los resultados mostraron una variación en la susceptibilidad de hasta 15,4 veces en tres años de una de las colonias aumento significativo de la CL50 observada que no fue considerado como un cambio significativo en la susceptibilidad. Empleando una metodología similar a la nuestra, sometiendo larvas de tres días de edad y no neonatas a dosis de 0,456-9,13 y 182,6 μg/mL de dieta, Ramírez-Romero et al. (2007) observaron una reducción significativa de supervivencia a los seis días de iniciado el experimento con la dosis mayor y a los 10 días con la dosis intermedia igualmente, encontraron pesos larvales menores con las tres dosis, pero nunca un 100% de mortalidad larval, lo cual se debe probablemente al uso inicial de larvas ya desarrolladas. Extrapolando, se podría deducir un comportamiento similar de la población del cogollero empleado en el estudio de Ramírez-Romero et al. (2007), quienes evaluaron una raza procedente de Le Magneraud INRA, Francia, como de aquel observado con la raza del insecto procedente del Tolima.
Desde antes de la liberación del híbrido de maíz evento MON810 en otros países, se realizaron investigaciones para determinar el control, mediante la planta transgénica del maíz del S. frugiperda. Buntin et al. (2001), encontraron en el año 1998, en el sur de Georgia (EE. UU.), una considerable reducción en el daño al follaje y al cogollo, pero observaron ataques de larvas en muchas mazorcas. Estas larvas, se desarrollaron muy lentamente, causando más daño a las mazorcas del Yieldgard® que a aquellas del testigo no transgénico. En el Brasil, Vilella et al. (2002), hallaron un aumento significante en la tolerancia del cogollero al cabo de cuatro generaciones de ser sometido a presión de selección por la toxina Cry1Ab. Waquil et al. (2004), evaluando la actividad biológica del Cry1Ab, y basado en la supervivencia de larvas, advirtieron la necesidad de formular estrategias de manejo de resistencia de la planta transgénica. Chilcutt et al. (2007), estableciendo la variación en densidades de la población larval del cogollero en mazorcas del híbrido transgénico, identificaron una densidad menor en la planta transformada, que en aquellas que servían de testigo durante una temporada de baja infestación de la plaga, mientras que bajo condiciones contrarias, las densidades de la plaga fueron similares en los dos híbridos. Finalmente, en otro experimento con razas procedentes del norte de la Argentina el porcentaje de mortalidad de larvas alimentadas con pedazos de hojas de maíz transgénico, NK 120 TDmax®, que expresa al Cry1Ab, solamente alcanzó un 68,8 ± 6,8% (Virla et al. 2008). Así, que en general los resultados de las diversas investigaciones, se pueden considerar hasta cierto punto contradictorios, pero contundentes en afirmar que existe la posibilidad de una rápida adquisición de tolerancia del S. frugiperda a la toxina Cry1Ab.
Otros resultados contradictorios se pueden observar en la Tabla 2. La CL50 (línea base de susceptibilidad) para la población de Anolaima es un 48,04% mayor, es decir, casi un tercio, que aquella estimada para el área maicera del Tolima. Esto, a pesar de que en el área de colección de la población del cogollero durante más de 12 años no se aplicó ningún producto comercial a base de Bt. La mayor CL50 se determinó para la segunda generación de laboratorio y una intermedia se estimó para la cuarta generación evaluada. La línea de regresión de las tres últimas generaciones muestra una pendiente poco pronunciada y un coeficiente de regresión cada vez más insignificante.
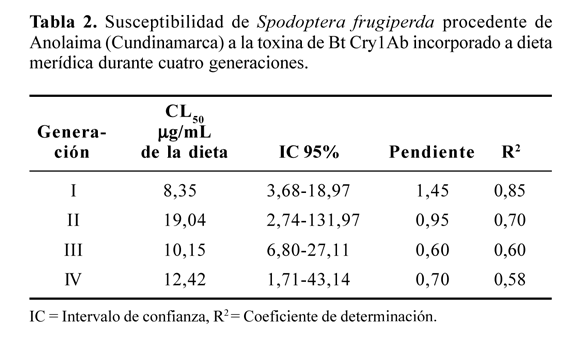
La amplitud del intervalo de confianza puede indicar una población genéticamente heterogénea o inclusive una raza geográfica del insecto, generada por los cruces con la población migratoria del Tolima, y podría ser interpretada como signo de intercambio de genes de poblaciones, dadas las migraciones que se podrían suceder entre el Tolima y la zona cafetera de Cundinamarca a través de las riveras de los ríos (Curí que desemboca en el río Bogotá) que conectan el valle del río Magdalena con el área de Anolaima. Estudios adicionales son imperativos para evaluar si realmente existe esta posibilidad de migración y la posterior adaptación de S. frugiperda procedente de un clima cálido a uno templado o viceversa.
La alimentación con follaje verde de larvas neonatas del cogollero procedentes del Tolima, resultó en un 96,66% de mortalidad al día siete, sólo sobrevivió una larva, la cual alcanzó el estado de pupa con un peso de 202,8 mg a los 27 días. Las larvas del testigo, se desarrollaron normalmente, solo, se observó un 3,33% de mortalidad: La duración promedia del estado larval fue de 23±1 días, con un peso promedio de las pupas de 211,26 ± 18,12 mg. Arévalo (2008), alimentando larvas con tejido foliar verde de maíz, bajo las mismas condiciones de laboratorio, encontró una duración del estado larval muy similar (22,29 ± 2,31 días) y un peso de las pupas de 201,26 ± 28,02 mg, lo cual podría considerarse equivalente, teniendo en cuenta la gran variabilidad reportada en la literatura para estos dos parámetros del cogollero. Márquez et al. (1963 y 1964), citados por Arévalo (2008), mencionan por ejemplo una duración entre 20,3 y 28,5 días, mientras que Álvarez y Sánchez (1983), encontraron que el desarrollo larval, con alimento de hojas de sorgo, varió entre 10 y 16,73 días.
La alta mortalidad proporcionada por el tejido transgénico era de esperarse y corresponde a lo observado en el campo para larvas de los primeros dos instares que consumen follaje antes de meterse al cogollo. Estudios iniciales realizados por Williams et al. (1997) mostraron que los híbridos transgénicos soportaban un daño follar significativamente menor que su contraparte no transgénica y además que larvas alimentadas con dietas que contenían básicamente hojas liofilizadas reducían considerablemente su crecimiento. Buntin et al. (2004) reportan que MON810 evitó consistentemente el daño al cogollo durante los años 1999 y 2000 en Georgia inclusive en siembras tardías de maíz en las cuales normalmente se presentan las infestaciones económicas de S. frugiperda. Virla et al. (2008), al contrario, evaluando 18 poblaciones argentinas del cogollero alimentadas con follaje de maíz transgénico (NK 120 TDmax®) que posee el Cry1Ab incorporado encontraron que a las 78 horas solamente un 40% de las razas mostraron un valor de mortalidad mayor a 75%.
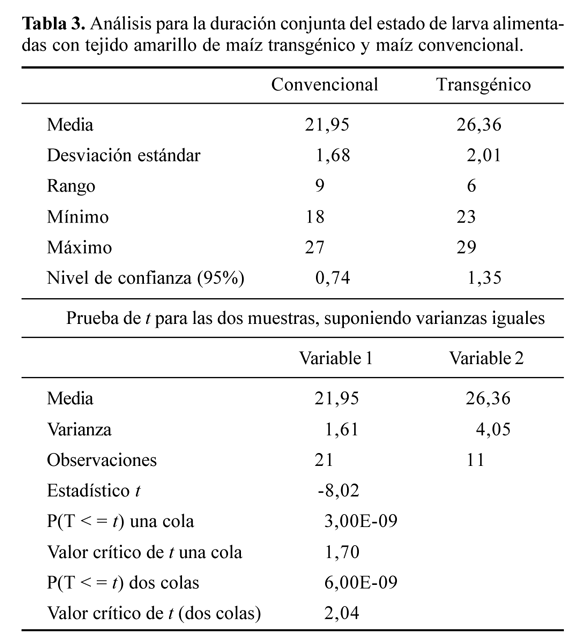
Se pueden considerar los resultados al emplear el tejido amarillo (TA) en la alimentación de las larvas. Tanto para la duración del estado larval como para el peso de la pupa se observaron diferencias significativas entre el tratamiento convencional versus el transgénico (Tablas 3 y 4). Al día siete del inicio del experimento se detectó en el testigo un 4,16% de mortalidad de las larvas y solamente se siguió el desarrollo a 23, de las cuales una se murió a los 14 días de haberse iniciado la investigación. La mortalidad inicial, se atribuyó al canibalismo, característico de la especie. Para evitar este fenómeno, el cual se detecta por los excrementos negros de las larvas que han consumido a otras, se sugiere la individualización de las larvas a más tardar al cuarto día. Para los individuos alimentados con tejido amarillo transgénico (cogollo), al día siete se determinó un 61,70% de mortalidad. A los 14 días se cambió el alimento a pedazos de mazorca tierna, envueltos en el capacho (brácteas que envuelven la mazorca); ya en este momento la mortalidad había aumentado a 76,59%. Los sobrevivientes siguieron su desarrollo normal. La duración del estado larval en el testigo fue de 21,95 ± 0,74 días (media ± D.E.), tiempo considerado normal para las condiciones del ensayo. Para los individuos alimentados con TA transgénico, este tiempo fue estadísticamente diferente al anterior y correspondió a 26,36 ± 1,35 días, lo cual muestra la influencia de la concentración, aunque baja del Cry1Ab en las estructuras de bajo contenido de clorofila.
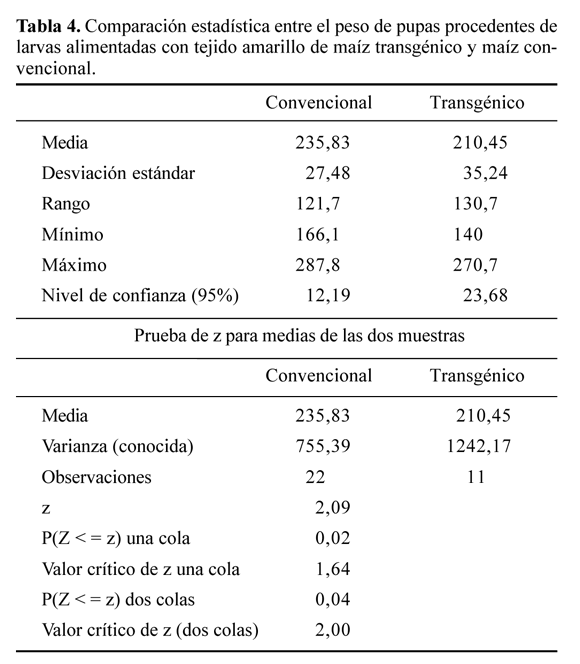
El peso de las pupas 24 horas después de formarse este estado, fue para el TA convencional de 235,82 ± 12,18 mg y para las pupas procedentes de larvas alimentadas con TA transgénico de 210,45 ± 23,67 mg. La prueba de z (Tabla 4) detecta una diferencia significativa entre los pesos y un rango mayor y significativo, al comparar las medias de los resultados de los dos grupos. Se atribuye esta diferencia a la existencia de la concentración del Cry1Ab en el tejido del maíz transgénico que no causa un 100% de mortalidad, la cual, aunque sí afectó tanto el desarrollo inicial larval y el peso de las pupas sobrevivientes, no influyó en general en el desarrollo de los sobrevivientes.
La concentración de la toxina Cry1Ab en la mazorca tierna fue de 0,82 ppm con un rango de 0,59-1,01 (datos sin publicar), es decir una concentración subletal, la cual bajo condiciones de campo podría tener una influencia marcada sobre la evolución de la resistencia al Cry1Ab del insecto. El comportamiento observado y la supervivencia de S. frugiperda en tejido amarillo del Yieldgard®, bajo condiciones de laboratorio explica la existencia observada en el campo, de larvas de III y IV instar, alimentándose de espigas, cubiertas de vainas amarillas. La dosis subletal en el cogollo y en otras estructuras con bajo contenido de clorofila, puede permitir a través del tiempo la generación de poblaciones resistentes y además resultar contradictorio para la teoría de los refugios. Abel y Adamczyk (2004) obtuvieron resultados similares al evaluar diferentes perfiles de color de hojas de maíz transgénico.
Determinaron, que a medida que disminuye el color, de verde a blanco amarillo, de la hoja número nueve de la planta, disminuye la concentración de la toxina y aumenta el peso de las larvas del cogollero alimentados con el tejido respectivo. Sus resultados demostraron una diferencia significativa entre las concentraciones halladas en el tejido verde, verde-amarillo y blanco amarillo. Concluyeron estos autores que tejidos con bajo contenido de clorofila no expresan completamente la proteína tóxica. Otros estudios, pero con el algodonero y Helicoverpa zea (Boddie, 1850), encontraron también expresiones diferenciales de la δ-endotoxina en diversos tejidos y una alta correlación entre la supervivencia y el desarrollo del insecto, alimentado con estas estructuras (Adamczyk et al. 2001a, 2001b). Igualmente, el Grupo Fitosanidad U.D.C.A (2008), determinó una muy baja concentración del Cry1Ac, 0,65 ppm, en flores sin fecundar, blanco-amarillas del algodonero, en comparación con 4,42ppm detectados en hojas terminales.
Wu et al. (2008) basado en un estudio realizado durante 15 años evaluando las fluctuaciones poblacionales de Helicoverpa armigera (Hübner, 1808), cinco años antes y 11 después de la introducción del algodón transgénico en diferentes provincias de la China, postuló que la disminución en general de la población de la plaga, se debe al control que ejerce sobre ella la toxina incorporada a la planta. Sugieren, además estos autores que la planta transgénica no solamente controla la plaga sino que puede reducir su presencia en otros huéspedes y disminuir la necesidad de control químico. Este caso hace referencia a la existencia de una concentración adecuada de la toxina en el tejido afectado, lo cual aparentemente no ocurre ni con el híbrido de maíz, ni con el algodonero transgénico. Se asume por lo tanto que a través del tiempo, no disminuiría la población de S. frugiperda en el área del Tolima, a pesar de siembras continuas del híbrido que contiene el Cry1Ab. La población del cogollero se mantendrá al alimentarse las larvas sobrevivientes, ante todo de las espigas del maíz que no contienen una concentración letal. Además, se considera la posibilidad de resistencia cruzada a los diversos Crys del Bt, Cry1Ab y Cry1Ac en este caso, fenómeno, cuya existencia se debería evaluar en nuestro medio con nuestras razas de S. frugiperda. Esta última consideración sugiere la urgencia de evaluaciones similares con el maíz transgénico que contiene el gen Cry1F, Herculex I (TC-1507), ya aprobado para su siembra en Colombia (ICA 2007b) y con otras toxinas incorporadas, para incluir en un plan de manejo de resistencia el uso alterno de maíces que poseen otras toxinas con receptores diferentes a las toxinas mencionadas.
Agradecimiento
A COLCIENCIAS, a la Universidad de Ciencias Aplicadas y Ambientales U.D.C.A por la financiación del proyecto y a FENALCE por su colaboración logística y técnica.
Literatura citada
ABEL, C. A.; POLLAN, M. C. 2004. Field resistance of Bacillus thuringiensis Berliner transformed maize to fall armyworm (Lepidoptera: Noctuidae) and southwestern corn borer (Lepidoptera: Crambidae) leaf feeding. Journal of Entomological Science 39 (3): 325-336. [ Links ]
ABEL, C. A.; ADAMCZYK, Jr., J. J. 2004. Relative concentration of Cr1A in Maize leaves and cotton bolls with diverse chlorophyll content and corresponding larval development of fall armyworm (Lepidoptera: Noctuidae) and southwestern corn borer (Lepidoptera: Crambidae) on maize whorl leaf profiles. Journal of Economic Entomology 97 (5): 1737-1744. [ Links ]
ADAMCZYK, Jr., J. J.; HARDEE, D. D.; ADAMS, L. C.; SUMMERFORD, D. V. 2001a. Correlating differences in larval survival and development of bollworm (Lepidoptera: Noctuidae) and fall armyworm (Lepidoptera:Noctuidae) to differential expression of Cry1A(c) a-endotoxin in various plant parts among commercial cultivars of transgenic Bacillus thuringiensis cotton. Journal of Economic Entomology 94 (1): 284-290. [ Links ]
ADAMCZYK, Jr., J. J.; ADAMS, L. C.; HARDEE, D. D. 2001b. Field efficacy and seasonal expression profiles for terminal leaves of single and double Bacillus thuringiensis toxin cotton genotypes. Journal of Economic Entomology 94 (4): 1589-1593. [ Links ]
AKHURST, R. J.; JAMES, W.; BIRD, L. J.; BEARD, C. 2003. Resistance to the Cry1Ac a-endotoxin of Bacillus thuringiensis in the cotton bollworm Helicoverpa armigera (Lepidoptera: Noctuidae). Journal of Economic Entomology 96 (4): 1290- 1299. [ Links ]
ÁLVAREZ, J. A.; SÁNCHEZ, G. 1983. Variación en el número de instares de Spodoptera frugiperda (J. E. Smith). Revista Colombiana de Entomología. 9 (1, 2, 3 y 4): 43-49. [ Links ]
ARÉVALO M., H. A. 2008. Evaluación de dietas merídicas para la cría de Spodoptera frugiperda (J. E. Smith) y Heliothis virescens (Fabricius) (Lepidoptera; Noctuidae). Trabajo de Grado para obtener el título de Ingeniero Agrónomo. Facultad de Ingeniería Agronómica. U.D.C.A Bogotá. [ Links ]
ÁVILA, C. J.; DEGRANDE, P. E.; GÓMEZ, S.A. 1997. Insetosprega: reconhecimento, comportamento, danos e controle, pp. 157-180. In: Empresa Brasileira de Pesquisa Agropecuária, EMBRAPA. Milho informações técnicas. Dourados. [ Links ]
BORRERO, F.; ZENNER DE POLANÍA, I. 1998. Resistencia potencial de Spodoptera frugiperda a Bacillus thuringiensis subesp. kurstaki. Manejo Integrado de Plagas (Costa Rica) 47: 18-23. [ Links ]
BUNTIN, G. D.; LEE, B. D.; WILSON, D. M.; McPHERSON, R. M. 2001. Evaluation of Yieldgard transgenic resistance for control of fall armyworm and corn earworm (Lepidoptera: Noctuidae) on corn. Florida Entomologist 84 (1): 37-42. [ Links ]
BUNTIN, G. D.; ALL, J. N.; LEE, R. D.; WILSON, D. M. 2004. Plant-incorporated Bacillus thuringiensis resistance for control of fall armyworm and corn earworm (Lepidoptera: Noctuidae) in corn. Journal of Economic Entomology 97 (5): 1603-1611. [ Links ]
CHILCUTT, CH. F.; ODVODY, G.N.; CORREA, J. C.; REMMERS, J. 2007. Effects of Bacillus thuringiensis transgenic corn on corn earworm and fall armyworm (Lepidoptera: Noctuidae) densities. Journal of Economic Entomology 100 (2): 327-334. [ Links ]
EPA (U. S. Environmental Protection Agency). 1998. The Environmental Protection Agency´s white paper on Bt plant-pesticide resistance management. Biopesticides and Pollution Prevention Division, Office of Pesticide Programs. Washington, DC. No. 739-S-98-001. [ Links ]
FENALCE (Federación Nacional de Cultivadores de Cereales y Leguminosas). 2008. Informe de Gestión 2007. Ed. Produmedios, Bogotá. 103 p. [ Links ]
FERNÁNDES, O. D.; PARRA, J. R. P.; NETO, A. F.; PÍCOLI, R.; BROGATTO, A. F.; DEMÉTRIO, C. G. B. 2003. Efeito do milho geneticamente modificado Mon810 sobre a lagarta-do-cartucho Spodoptera frugiperda (Smith, 1797) (Lepidoptera: Noctuidae). Revista Brasileira de Milho e Sorgo 2 (2): 25-35. [ Links ]
FUXA, J. R. 1987. Spodoptera frugiperda susceptibility to nuclear polyhedrosis virus isolates with reference to insect migration. Environmental Entomology 18 (1): 218-223. [ Links ]
GARCÍA ROA, F., MOSQUERA, A. T.; VARGAS S., C. A.; ROJAS, A. L. 1999. Manejo integrado del gusano cogollero del maíz Spodoptera frugiperda (J. E. Smith). CORPOICA, Palmira, Boletín Técnico No. 7. 18 p. [ Links ]
GRUPO FITOSANIDAD U.D.C.A. 2008. Propuesta de manejo en cultivos transgénicos. Folleto Divulgativo. Universidad de Ciencias Aplicadas y Ambientales U.D.C.A. 11 p. [ Links ]
ICA (Instituto Colombiano Agropecario). 2007a. Resolución No 00465 (26 feb 2007) por la cual se autorizan siembras de maíz con la tecnología Yieldgard® (Mon 810). 7 p. [ Links ]
ICA (Instituto Colombiano Agropecario). 2007b. Resolución No 00464 (26 feb 2007) por la cual se autorizan siembras de maíz con la tecnología Herculex I (TC-1507). 7 p. [ Links ]
LYNCH, R. E.; HAMM, J. J.; MYERS, R. E.; GUYER, D.; STEIN, J. 2003. Baseline susceptibility of the fall armyworm (Lepidoptera: Noctuidae) to Cry1Ab toxin: 1996-2000. Journal Entomological Science 38 (3): 377-385. [ Links ]
MALO, E. A.; CRUZ-LÓPEZ, L.; VALLE-MORA, J.; VIRGEN, A.; SÁNCHEZ, J. A.; ROJAS, J. C. 2001. Evaluation of commercial pheromone lures and traps for monitoring male fall armyworm (Lepidoptera: Noctuidae) in the coastal region of Chiapas, Mexico. Florida Entomologist 84: 659-664. [ Links ]
NAGOSHI, R. N.; MEAGHER, R. L.; FLANDERS, K.; GORE, J.; JACKSON, R.; LÓPEZ, J.; ARMSTRONG, J. S.; BUNTIN, G. D.; SANSONE, C.; LEONARD, B. R. 2008. Using haplotypes to monitor migration of fall armyworm (Lepidoptera: Noctuidae) corn-strain population from Texas and Florida. Journal of Economic Entomology 101 (3): 742-749. [ Links ]
PITRE, H. N. 1988. Relationship of fall armyworm (Lepidoptera. Noctuidae) from Florida, Honduras, Jamaica, and Mississippi: susceptibility to insecticides with reference to migration. Florida Entomologist 71 (1): 56-61. [ Links ]
POLANCZYK, R. A.; SILVA, R. F.; FIUZA, L. M. 2004. Isolamento de Bacillus thuringiensis Berliner a partir de amostras de solos e sua patogenicidade para Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae). Revista Brasileira de Agrociência, Pelotas 10 (2): 209-214. [ Links ]
POSADA-O., L. 1989. Lista de insectos dañinos y otras plagas en Colombia. Instituto Colombiano Agropecuario, ICA. Boletín Técnico No. 43. Produmedios, Bogotá. 662 p. [ Links ]
RAMÍREZ-ROMERO, R.; BERNAL, J. S.; CHAUFAUX, J.; KAISER, L. 2007. Impact assessment of Bt-maize on a moth parasitoid, Cotesia marginiventris (Hymenoptera: Braconidae), via host exposure to purified Cry1Ab protein or Bt-plants. Crop Protection 26: 953-962. [ Links ]
ROJAS, J. C.; VIRGEN, A.; MALO, E. A. 2004. Seasonal and nocturnal flight activity of Spodoptera frugiperda males (Lepidoptera. Noctuidae) monitored by pheromone traps in the coast of Chiapas, Mexico. Florida Entomologist 87 (4): 496-503. [ Links ]
TABASHNIK, B. E.; CARRIERE, Y.; DENNCHY, T. J.; MORIN, S.; SISTERSON, M. S.; ROUSH, R. T.; SHELTON, A. M.; ZHAO, J. Z. 2003. Insect resistance to transgenic Bt crops: lessons from the laboratory and fields. Journal of Economic Entomology 96 (4): 1031-1038. [ Links ]
VANEGAS-A., H.; POLANÍA-F., F. 2002. Maíz en la zona cafetera. Boletín Informativo TECNIFenalce 3 (4): 1-8. [ Links ]
VILARINHO, E. C.; FERNANDES, O. A.; OMOTO, C.; HUNT, T. E. 2006. Oil-soluble dyes for marking Spodoptera frugiperda (Lepidoptera: Noctuidae). Journal of Economic Entomology 99 (6): 2110-2115. [ Links ]
VILELLA, F. M. F.; WAQUIL, J. M.; VILELA, E. F.; SIEGFIRED, B. D.; FOSTER, J. E. 2002. Selection of the fall armyworm, Spodoptera frugiperda (Smith) (Lepidoptera: Noctuidae) for survival on Cry1A(b) Bt toxin. Revista Brasileira de Milho e Sorgo 1 (3): 12-17. [ Links ]
VIRLA, E. G.; COLOMBO, M. V.; BERTA, D. C.; VALVERDE, L. 1999. El complejo de parasitoides del "gusano cogollero" del maíz, Spdoptera frugiperda, en la República Argentina (Insecta, Lepidoptera). Neotropica 45: 3-12. [ Links ]
VIRLA, E. G.; ÁLVAREZ, A.; LOTO, F.; PERA, L.M.; BAIGORI, M. 2008. Fall armyworm strains (Lepidoptera: Noctuidae) in Argentina, their associate host plants and response to different mortality factors in laboratory. Florida Entomologist 91 (1): 63- 69. [ Links ]
WAQUIL, J. M.; VILLELA, F. M. F.; FOSTER, J. E. 2002. Resistencia do milho (Zea mays L.) transgénico a lagarta-do-cartucho, Spodoptera frugiperda (Smith) (Lepidoptera: Noctuidae). Revista Brasileira de Milho e Sorgo 1 (3): 1-11. [ Links ]
WAQUIL, J. M.; VILLELA, F. M. F.; SIEGFRIED, B. D.; FOSTER, J. E. 2004. Actividade biológica das toxinas do Bt, Cry1A(b) e Cry1F em Spodoptera frugiperda (Smith) (Lepidoptera: Noctuidae). Revista Brasileira de Milho e Sorgo 3 (2): 153-163. [ Links ]
WESTBROOK, J. K.; SPARKS, A. N. 1986. The role of atmospheric transport in the economic fall armyworm (Lepidoptera. Noctuidae) infestation in the southeastern United States in 1977. Florida Entomologist 69 (3): 494-502. [ Links ]
WILLIAMS, W. P.; SAGERS, J. B.; HANTEN, J. A.; DAVIS, F. M. BUCKLEY, P. M. 1997. Transgenic corn evaluated for resistance to fall armyworm and southwestern corn borer. Crop Science 37 (3): 957-962. [ Links ]
WU, K-M.; LU, Y-H.; FENG, H-Q.; JIANG, Y-Y.; ZHAO, J-Z. 2008. Suppression of cotton bollworm in multiple crops in China in areas with Bt toxin-containing cotton. Science 321: 1676-1678. [ Links ]
ZENNER, I.; ÁLVAREZ, A.; BARRETO, S. 2006. Influence of parasitism by Chelonus insularis Cresson (Hymenoptera: Braconidae, Cheloninae) on the susceptibility of Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) to insecticides. Neotropical Entomology 35 (6): 818-822. [ Links ]
ZENNER DE POLANÍA, I.; BORRERO FONSECA, F. 1994. Resistencia del cogollero del maíz, Spodoptera frugipeda (J. E. Smith) a algunos insecticidas y su manejo, pp. 35-41. En: Memorias Seminario Internacional sobre el cultivo de maíz y sorgo sus principales plagas y enfermedades. Comité Interinstitucional sobre los cultivos de sorgo y maíz. C.I. Tibaitatá. Enero 25-30 de 1993. Produmedios (Bogotá). [ Links ]
ZENNER DE POLANÍA, I.; ÁLVAREZ R., J. A.; MEJÍA C., R.; BAYONA R., M. A. 2005. Influencia de la toxina Cry1Ac del Bacillus thuringiensis sobre el desarrollo del cogollero del maíz, Spodoptera frugiperda (J. E. Smith). Revista U.D.C.A Actualidad & Divulgación Científica 8 (2): 129-139. [ Links ]
ZENNER DE POLANÍA, I.; ÁLVAREZ R., J. A.; ARÉVALO M., H. A.; MEJÍA C., R.; BAYONA R.; M. A. 2008. Susceptibilidad de cuatro nóctuidos plaga (Lepidoptera) al gene Cry1Ac del Bacillus thuringiensis incorporado al algodonero. Revista Colombiana de Entomología 34 (1): 41-50. [ Links ]
Recibido: 15-nov-2008 - Aceptado: 30-mar-2009

