










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488On-line version ISSN 2665-4385
Rev. Colomb. Entomol. vol.35 no.1 Bogotá Jan./June 2009
Nota cientifica
Superparasitismo de Palmistichus elaeisis (Hymenoptera: Eulophidae) y comportamiento de defensa de dos hospederos
Superparasitism by Palmistichus elaeisis (Hymenoptera: Eulophidae) and defense behaviors of two hosts
MARCUS ALVARENGA SOARES1, CAROLINA TORRES GUTIERREZ2*, JOSÉ COLA ZANUNCIO3, ALINE RODRIGUES PORTO PEDROSA4 y ALEXANDRE SIMÕES LORENZON5
1 Estudiante de Doctorado, Curso de Entomología. Agrónomo, M. Sc. Universidad Federal de Viçosa, UFV. Avenida Peter Henry Rolfs, s/n Campus Universitario 36570-000, Viçosa - MG, Brasil. marcusasoares@yahoo.com.br.
2 Autor para correspondencia: Investigadora Línea Entomología Médica, Bióloga, M. Sc. Programa Estudio y Control de Enfermedades Tropicales - PECET. Universidad de Antioquia. Carrera 53 No. 61-30. Torre 2. Laboratorio 632. Sede de Investigación Universitaria, Medellín, Colombia. carolina.torres@siu.udea.edu.co.
3 Profesor titular, Curso de Entomología. Ing. Forestal, Ph. D. Universidad Federal de Viçosa, UFV. Avenida Peter Henry Rolfs, s/n Campus Universitario 36570-000, Viçosa - MG, Brasil. zanuncio@ufv.br.
4 Estudiante de Biología, Universidad Federal de Viçosa, UFV. Avenida Peter Henry Rolfs, s/n Campus Universitario 36570-000, Viçosa - MG, Brasil. aline_pedrosa@hotmail.com.
5 Estudiante de Ingeniería Forestal, Universidad Federal de Viçosa, UFV. Avenida Peter Henry Rolfs, s/n Campus Universitario 36570-000, Viçosa - MG, Brasil. alelorenzon@yahoo.com.br.
Resumen: Varias familias de parasitoides son capaces de discriminar entre los hospederos parasitados y no-parasitados. Sin embargo, algunos casos de superparasitismo pueden ocurrir en especies de Eulophidae. El objetivo del presente estudio fue investigar si hembras de Palmistichus elaeisis (Eulophidae) superparasitan pupas de Thyrinteina arnobia (Lepidoptera: Geometridae) y de Hylesia sp. (Lepidoptera: Saturniidae) en laboratorio, describiendo mecanismos de defensa de dichos hospederos. El experimento se realizó en el insectario de la Universidade Federal de Viçosa, Brazil. Se utilizaron cajas de Petri como cámaras de oviposición, conteniendo dos pupas de T. arnobia, una de ellas parasitada por P. elaeisis y otra sin parasitar. De igual forma se mantuvieron cajas de Petri con dos pupas de Hylesia sp. Para los dos tratamientos, se liberaron diez hembras de P. elaeisis en la parte central de cada caja de Petri, el cuál constó una repetición. Se usó un total de ocho repeticiones por cada tratamientio. Se observó la preferencia de oviposición de P. elaeisis sobre las pupas hospederas durante seis horas y se analizaron mediante la prueba de chi-cuadrado (X2) (P 0,01). Observando el comportamiento de defensa de los hospederos, las pupas de T. arnobia presentaron movimientos giratorios, lo que indujo el superparasitismo en las pupas de menor movilidad. Los individuos de Hylesia sp. no presentaron comportamiento de defensa, y en este caso la discriminación llegó a una preferencia clara para las pupas no parasitadas.
Palabras clave: Defoliadores. Discriminación. Eucalyptus. Parasitoides. Pupas.
Abstract: Several families of parasitoids are capable of discriminating among parasitized and unparasitized hosts. However, some cases of superparasitism have been recorded in species of Eulophidae. The objective of this study was to investigate whether females of Palmistichus elaeisis (Eulophidae) superparasitize pupae of Thyrinteinaarnobia (Lepidoptera: Geometridae) and Hylesia sp. (Lepidoptera: Saturniidae) in the laboratory and to describe the defense mechanisms of those hosts. The experiment was done in the insectary of the Universidade Federal de Viçosa, Brazil. Petri dishes were used as oviposition chambers and contained two T. arnobia pupae, one of them parasitized by P. elaeisis and the other not. The pupae were placed on each side of the dish. Petri dishes were likewise set up with two pupae of Hylesia sp. For both treatments, ten females of P. elaeisis were released in the center of the dish, which represented a replicate. A total of eight replicates were used for each treatment. The oviposition preference of P. elaeisis on host pupae was observed for six hours and analyzed using the chi-square test (X2) (P 0.01). In observing the defense behavior of the hosts, T. arnobia pupae showed spinning movements which in turn induced superparasitism in less mobile pupae. Individuals of Hylesia sp. did not show any defense behavior and in this case the discrimination led to a clear choice for the unparasitized pupae.
Key words: Defoliators. Discrimination. Eucalyptus. Parasitoids. Pupae.
Introducción
Las especies de eucalipto han sido utilizadas en Brasil, en grandes áreas de reforestación, que incrementan la vulnerabilidad de las plantas a insectos plaga (Soares et al. 2007). Thyrinteinaarnobia (Stoll, 1782) (Lepidoptera: Geometridae) ha sido muy estudiada, considerándose la principal oruga defoliadora de eucalipto en el Brasil (Oliveira et al. 2005). Por otro lado, especies del género Hylesia también han sido registradas en cultivos de eucalipto, con un alto número de individuos voraces (Draudt 1929). El parasitoide Palmistichus elaeisis Delvare y LaSalle, 1993 (Hymenoptera: Eulophidae) se destaca por ser enemigo natural de diversos lepidópteros plaga de especies forestales (Bittencourt y Berti Filho 1999), pudiendo parasitar pupas de T. arnobia y de especies del género Hylesia.
La mayoría de parasitoides son capaces de reconocer los hospederos parasitados por ellos mismos o por coespecíficos, una habilidad llamada “discriminación de hospedero” (van Alphen y Visser 1990; Nufio y Papaj 2001). Sin embargo, algunos parasitoides pueden ovipositar en hospederos previamente parasitados, comportamiento llamado “superparasitismo” (Khafagi y Hegazi 2008). El superparasitismo fue tratado inicialmente como un error de algunos parasitoides, debido a que, aparentemente, el gasto de tiempo y huevos en un hospedero, donde la progenie tiene poca posibilidad de supervivencia, no sería una ventaja adaptativa. Por otra parte, un nuevo ataque recibido por el hospedero podría causarle grave daño y llevarlo a la muerte (Godfray 1994). No obstante, algunos registros han abordado el superparasitismo como una estrategia adaptativa de los parasitoides que, en algunas situaciones, puede ser benéfica (van Alphen y Visser 1990). Este fenómeno ha sido observado cuando los hospederos son escasos y cuando la prole de un segundo parasitoide no es fuertemente afectada ante la competencia por recursos, con la prole del primer parasitoide (van Alphen y Visser 1990).
Algunas especies de lepidópteros poseen mecanismos morfológicos o comportamentales capaces de conferirles protección contra la localización y ataque por parasitoides (Potting et al. 1999). Sin embargo, muchos parasitoides modifican el comportamiento o reducen los movimientos de sus hospederos (Infante et al. 1994; Grosman et al. 2008) y esto podría facilitar la ocurrencia de superparasitismo. En este estudio se investigó si hembras de P. elaeisis superparasitan pupas hospederas de Thyrinteinaarnobia e Hylesia sp. en laboratorio, además de describir los posibles mecanismos de defensa de pupas previamente parasitadas y sin parasitar, contra dichos enemigos naturales.
Materiales y Métodos
El experimento se realizó en el laboratorio de Control Biológico de Insectos, del Departamento de Biología animal de la Universidad Federal de Viçosa (UFV), en Minas Gerais, Brasil. Las especies de lepidópteros utilizadas en el experimento fueron criadas entre el período de agosto a octubre del 2007. El experimento fue realizado en el mes de octubre de 2007. Los adultos de T. arnobia, se obtuvieron de colecciones de campo en cultivos de eucalipto, en el estado de Minas Gerais. Estos adultos se mantuvieron en un cuarto climatizado bajo las siguientes condiciones: temperatura 25 ± 2°C, foto-período 12 horas y humedad relativa 70 ± 10%. Los individuos de T. arnobia se separaron por parejas, colocándose en recipientes plásticos (500 ml) con tapa de abertura central cubierta por tela de malla fina. Al interior de los recipientes, se depositaron tiras de papel, adheridas a las tapas, para que en ellas se efectuara la oviposición. Los huevos de T. arnobia se colocaron en mangas entomológicas (0,70 x 0,40 m) de malla fina, envolviendo ramas de Eucalyptus cloeziana F. Muell. (Myrtaceae), en el área experimental del Insectario de la UFV. Cuando las orugas alcanzaron la fase de pupa, se retiraron de las mangas entomológicas, se colocaron en recipientes plásticos y se transfirieron para el laboratorio de Control Biológico de Insectos.
Los huevos de Hylesia sp. se colectaron en cultivos de eucalipto en el estado de Minas Gerais. Debido a la acción urticante de sus individuos, éstos fueron criados en recipientes plásticos (500 ml) en laboratorio, con temperatura de 25 ± 2°C, foto-período de 12 horas y humedad relativa de 70 ± 10%. Después de la emergencia de las larvas, se suministraron diariamente hojas de E. cloeziana, sujetadas por tubos plásticos (2,5 ml - de anestésico odontológico) con agua destilada en su interior.
Ocho pupas de T. arnobia y ocho pupas de Hylesia sp., con menos de 24 horas de edad, se individualizaron en tubos de vidrio (14 x 2,2 cm) y se expusieron al parasitismo por seis hembras de P. elaeisis, con 72 horas de emergencia y sin experiencia previa de oviposición. Las hembras estuvieron en contacto con los hospederos por 24 horas, a una temperatura de 25 ± 1°C, foto-período de 12 horas y humedad relativa de 70 ± 10%, en incubadora. Al final de este período, las hembras de P. elaeisis se retiraron de los tubos y se sacrificaron (Pereira et al., 2008). Una misma cantidad de pupas, de ambas especies, se mantuvo en recipientes plásticos, dentro de la incubadora, sin contacto con parasitoides.
Superparasitismo en pupas de Thyrinteinaarnobia e Hylesia sp. Las hembras de P. elaeisis se sometieron a dos tratamientos, en donde recibieron dos pupas de lepidópteros, una previamente parasitada (T1) y otra sin parasitar (T2). Se realizaron dos bioensayos, utilizando pupas de T. arnobia en el primero e Hylesia sp. en el segundo.
Para los bioensayos, se establecieron unidades experimentales o cámaras de oviposición, que consistieron en cajas de Petri (9,0 x 1,5 cm), conteniendo una pupa hospedera parasitada y una pupa sin parasitar, en sus lados opuestos. Las condiciones de dichas unidades experimentales se estandarizaron, estableciéndose la ubicación de las pupas dentro de las cajas de Petri y evitando que factores ambientales, como la luminosidad, interfirieran en la elección de los parasitoides (Jones et al. 2005). Posteriormente, en el centro de cada unidad experimental, se colocaron diez hembras de P. elaeisis con 72 horas de emergidas, fecundadas y alimentadas con miel. La variable analizada fue la preferencia de oviposición de hembras de P. elaeisis por pupas hospederas parasitadas y sin parasitar. El período de observación fue de seis horas, y en dicho tiempo, se contó el número de parasitoides ovipositando en cada pupa hospedera, en intervalos de ocho minutos. Para cada uno de los bioensayos, con T. arnobia y con Hylesia sp., se establecieron ocho unidades experimentales, cada una representando una repetición.
Se utilizó un diseño experimental aleatorio, incluyendo en el análisis solo las hembras que ovipositaron, y excluyendo aquellas que no ovipositaron en ninguno de los hospederos durante el período evaluado. La oviposición se determinó al observar la inserción del aparato ovipositor de la hembra de P. elaeisis en la superficie de la pupa hospedera y su permanencia en dicha posición por más de un minuto. El número máximo de parasitoides que ovipositó en cada pupa fue sumado, para el cálculo de la frecuencia de oviposición en las diferentes categorías de hospederos. Esta frecuencia se analizó por la prueba de Chi cuadrado (X2) (P ≤ 0,01) (Pearson 1904). Comportamiento de defensa de pupas de Thyrinteinaarnobia e Hylesia sp. El comportamiento de defensa de las pupas de T. arnobia e Hylesia sp. previamente parasitadas y sin parasitar se observó cuando estos hospederos fueron expuestos a las hembras de P. elaeisis en las cámaras de oviposición por seis horas consecutivas.
Resultados y Discusión
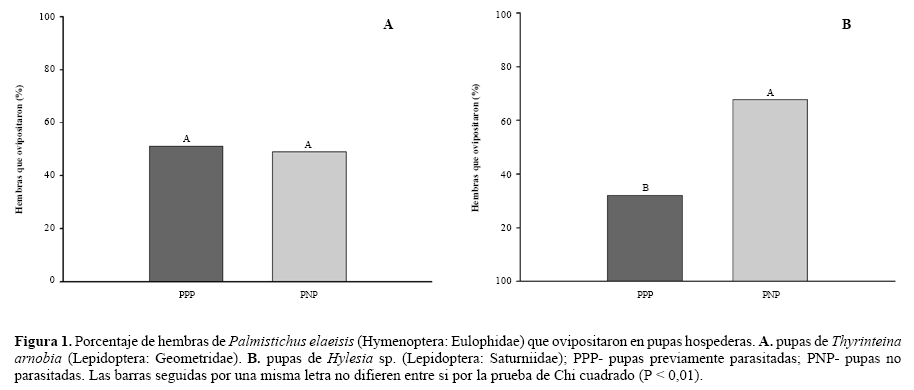
Superparasitismo en pupas de Thyrinteinaarnobia e Hylesia sp. De las 160 hembras de P. elaeisis utilizadas en el experimento, 101 se utilizaron para el cálculo de la frecuencia de oviposición en las pupas hospederas. Las hembras de P. elaeisis que recibieron pupas de T. arnobia no presentaron preferencia de oviposición (X2 = 0, 022, P ≤ 0, 01, n = 45) (Fig.1A). Las hembras de P. elaeisis que recibieron pupas de Hylesia sp. mostraron preferencia por ovipositar sobre pupas no parasitadas (X2 = 7,14, P ≤ 0, 01, n = 56) (Fig.1B).
La ausencia de discriminación de pupas de T. arnobia, observada en el parasitoide, indica superparasitismo en dichos hospederos. En ciertas situaciones, el superparasitismo puede representar una estrategia adaptativa, y un ejemplo de esto es el caso de hembras de parasitoides que atacan hospederos recientemente parasitados, con defensas inmunológicas debilitadas, momento en el cual sus larvas podrán tener mayor probabilidad de supervivencia (Khafagi y Hegazi 2008). Así mismo, cuando la probabilidad de encontrar hospederos no parasitados en ambientes silvestres es baja, el superparasitismo permitiría que hembras de parasitoides dejaran descendientes, a pesar de la ausencia de hospederos adecuados (Godfray 1994). De esta forma, evitar el superparasitismo en algunas situaciones, puede no ser beneficioso para un parasitoide (van Alphen y Visser 1990). Esto explicaría las frecuentes observaciones de superparasitismo en especies de la familia Eulophidae documentadas en la literatura, con registros hechos en laboratorio o campo, para Pediobius foveolatus (Crawford) (Shepard y Gale 1977), Hyssopus pallidus (Askew, 1964) (Zaviezo y Mills 1999), Melittobia australica (Girault, 1912) (Abe et al. 2005) y Phymastichus coffea (LaSalle) (Hymenoptera: Eulophidae)(Castillo et al. 2004; Jaramillo et al. 2006).
Comportamiento de defensa de pupas de Thyrinteinaarnobia e Hylesia sp. Las pupas de T. arnobia no parasitadas, una vez expuestas a las hembras de P. elaeisis, presentaron fuertes movimientos giratorios. En este caso, las pupas giraban 360º sobre su propio eje, y se movían al interior de las cámaras de oviposición (Fig. 2A). Este comportamiento protegió a las pupas del parasitismo, debido a que expulsaban a las hembras de P. elaeisis de la región comprendida por los segmentos abdominales. En estas pupas, solo los parasitoides que intentaron ovipositar en la región de la cabeza obtuvieron éxito, por no entrar en contacto con la caja de Petri durante el movimiento giratorio de la pupa. Las pupas de T. arnobia previamente parasitadas, presentaron tan solo algunos movimientos suaves de los segmentos abdominales (Fig. 2B) o no presentaron ningún movimiento.
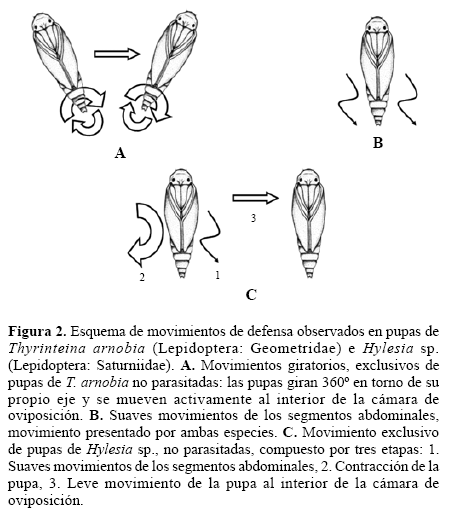
Las pupas de Hylesia sp., no parasitadas, una vez expuestas a hembras de P. elaeisis presentaron movimientos suaves en los segmentos abdominales o se contraían generando un pequeño movimiento al interior de la cámara de oviposición (Fig. 2C). Las pupas parasitadas de este género presentaron movimientos leves de los segmentos abdominales (Fig. 2B), o no registraron movimientos. La elección de P. elaeisis por pupas de T. arnobia, previamente parasitadas, puede ser influenciada por la presencia de mecanismos de defensa en las pupas hospederas no parasitadas. Las pupas de algunas especies de lepidópteros, incorporaron algunos sistemas de defensa, incluyendo una cutícula dura, movimientos giratorios (Cole 1959) y fuertes contracciones de la región abdominal (Gross 1993). De forma semejante, el movimiento giratorio desarrollado por las pupas de T. arnobia no parasitadas, evitó el ataque de muchos parasitoides. Por otro lado, las pupas de T. arnobia parasitadas no presentaron movimiento giratorio, mostrando que sus reflejos estaban reducidos (Infante et al. 1994). La ausencia de movimiento probablemente favorece el superparasitismo del hospedero, dado que representa una presa fácil para el ataque de enemigos naturales. Las pupas de Hylesia sp. presentaron menor capacidad de movimiento al compararlas con las pupas de T. arnobia y en este caso, tan solo la discriminación de hospederos influyó en la elección de las hembras de P. elaeisis, explicando así la fuerte preferencia de estos parasitoides por las pupas no parasitadas. Los resultados del presente estudio señalan los mecanismos de defensa de pupas no parasitadas, como uno de los factores que influencian el superparasitismo de T. arnobia y sugieren la capacidad de las hembras de P. elaeisis de discriminar pupas hospederas de Hylesia sp.
Agradecimientos
A Elizabeth C. Pedroso por las ilustraciones. Al Consejo Nacional de Desarrollo Científico y Tecnológico (CNPq) (Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq)), por las becas y auxilios económicos concedidos.
Literatura citada
ABE, J.; KAMIMURA, Y.; SHIMADA, M. 2005. Individual sex ratios and offspring emergence patterns in a parasitoid wasp, Melittobia australica (Eulophidae), with superparasitism and lethal combat among sons. Behavioral Ecology and Sociobiology 57: 366-373. [ Links ]
BITTENCOURT M. A. L; BERTI FILHO, E. 1999. Preferência de Palmistichus elaeisis por pupas de diferentes lepidópteros praga. Scientia Agricola 56: 1281-1283. [ Links ]
CASTILLO, A.; INFANTE, F.; VERA-GRAZIANO, J.; TRUJILLO, J. 2004. Hostdiscrimination by Phymastichus coffea, a parasitoid of the coffee berry borer. BioControl 49: 655-663. [ Links ]
COLE, L. R. 1959. On the defenses of lepidopterous pupae in relation to the oviposition behavior of certain Ichneumonidae. Journal of the Lepidopterists Society 13: 1-10. [ Links ]
DRAUDT, M. 1929. Familie Saturniidae, pp. 713-827. En: Seitz, A. (Ed.). Die Gross-Schmetterlinge der Erde. 6. Die Amerikanischen Spinner und Schwärmer. Stuttgart, Alfred Kernen, 508 p. [ Links ]
GODFRAY, H. C. J. 1994. Parasitoids, behavioral and evolutionary ecology. Princeton: Princeton University Press, 473 p. [ Links ]
GROSMAN, A. H.; JANSSEN, A.; DE BRITO, E. F.; CORDEIRO, E. G.; COLARES, F.; FONSECA, J. O.; LIMA, E.; PALLINE, A.; SABELIS, M. W. 2008. Parasitoid increases survival of its pupae by inducing hosts to fight predators. PloS ONE 3: e2276. [ Links ]
GROSS, P. 1993. Insect behavioral and morphological defenses against parasitoids. Annual Review of Entomology 38: 251-273. [ Links ]
INFANTE, F.; MURPHY, S. T.; BARRERA, J. F.; GÓMEZ, J. W.; DAMON, A. 1994. Cría de Phymastichus coffea parasitoide de la broca del café y algunas notas sobre su historia de vida. Southwestern Entomologist 19: 313-315. [ Links ]
JARAMILLO, J.; BORGEMEISTER, C.; SETAMOU, M. 2006. Field superparasitism by Phymastichus coffea, a parasitoid of adult coffee berry borer, Hypothenemus hampei. Entomologia Experimentalis et Applicata 119: 231-237. [ Links ]
JONES, G. A.; SIEVING, K. E.; AVERY, M. L.; MEAGHER, R. L. 2005. Parasitized and non-parasitized prey selectivity by an insectivorous bird. Crop Protection 24: 185-189. [ Links ]
KHAFAGI, W. E.; HEGAZI, E. M. 2008. Does superparasitism improve host suitability for parasitoid development? A case study in the Microplitis rufiventris - Spodoptera littoralis system. BioControl 53: 427-438. [ Links ]
NUFIO, C. R.; PAPAJ, D. R. 2001. Host marking behavior in phytophagous insects and parasitoids. Entomologia Experimentalis et Applicata 99: 273-293. [ Links ]
OLIVEIRA, H. N.; ZANUNCIO, J. C.; PEDRUZZI, E. P.; ESPINDULA, M. C. 2005. Rearing of Thyrinteina arnobia (Lepidoptera: Geometridae) on guava and eucalyptus in laboratory. Brazilian Archives of Biology and Technology 48: 801-806. [ Links ]
PEARSON, K. 1904. On the theory of contingency tables and its relation to association and normal correlation. Draper's Co. Res, pp. 443-475. En: Mem. Biometric Ser. 1. Reprinted (1948) in Karl Pearson's Early Papers, Cambridge University Press. [ Links ]
PEREIRA, F. F.; ZANUNCIO, T. V.; ZANUNCIO, J. C.; PRATISSOLI, D.; TAVARES, M. T. 2008. Species of Lepidoptera defoliators of Eucalyptus as new host for the parasitoid Palmistichus elaeisis (Hymenoptera: Eulophidae). Brazilian Archives of Biology and Technology 51: 259-262. [ Links ]
POTTING, R. P. J.; VERMEULEN, N. E.; COLONG, D. E. 1999. Active defense of herbivorous hosts against parasitism: Adult parasitoid mortality risk involved in attacking a concealed stemboring host. Entomologia Experimentalis et Applicata 91: 143-148. [ Links ]
SHEPARD, M.; GALE, G. T. 1977. Superparasitism of Epilachna varivestis [Col.: Coccinellidae] by Pediobius foveolatus [Hym.: Eulophidae]: influence of temperature and parasitoid-host ratio. Entomophaga 22: 315-321. [ Links ]
SOARES, M. A.; LEITE, G. L. D.; ZANUNCIO, J. C.; ROCHA, S. L.; DE SÁ, V. G. M.; SERRÃO J. E. 2007. Flight capacity, parasitism and emergence of five Trichogramma (Hymenoptera: Trichogrammatidae) species from forest areas in Brazil. Phytoparasitica 35: 314-318. [ Links ]
VAN ALPHEN, J. J. M.; VISSER, M. E. 1990. Superparasitism as an adaptive strategy for insect parasitoids. Annual Review of Entomology 35: 59-79. [ Links ]
ZAVIEZO, T.; MILLS, N. 1999. Aspects of the biology of Hyssopus pallidus (Hymenoptera: Eulophidae), a parasitoid of the codling moth (Lepidoptera: Olethreutidae). Environmental Entomology 28: 748-754. [ Links ]
Recibido: 26-abr-2008 - Aceptado: 12-feb-2009


{kind=link}