










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488On-line version ISSN 2665-4385
Rev. Colomb. Entomol. vol.35 no.2 Bogotá July/Dec. 2009
Análisis morfométrico de la pigmentación en especies colombianas de Drosophila (Diptera: Drosophilidae) del grupo repleta
Morphometric analysis of the pigmentation in colombian species of Drosophila (Diptera: Drosophilidae) from the repleta group
RODRIGO PRIETO S.1 y MARÍA MAGDALENA E. DE POLANCO2
1 M. Sc. Biología. Departamento de Biología. Universidad de Nariño. Ciudadela Universitaria, sede Torobajo. Tel.: 7311449 Ext. 263. San Juan de Pasto, Colombia. bioprieto@yahoo.es. Autor para correspondencia.
2 Ph. D. Genética. Lab. de Citogenética, Fac. de Ciencias, Universidad del Tolima. Cll. 42 - Cra. 4 Santa Helena. Ibagué, Colombia. mmpol@lycos.com.
Recibido: 29-ene-2009 - Aceptado: 21-sep-2009
Resumen: Se comparó la pigmentación abdominal de poblaciones aisladas de las especies del grupo repleta: Drosophila martensis, D. starmeri y D. uniseta (cluster martensis). El propósito fue detectar si la distancia que las separa, aunada a la heterogeneidad ambiental, están relacionadas con un proceso de formación de razas geográficas. De otra parte, se comparó la variación del mencionado carácter en D. aldrichi, del conglomerado mulleri, con la de las especies del conglomerado martensis, tratando de contrastar las relaciones evolutivas entre estas especies. Se digitalizó el contorno de la tercera mancha abdominal con descriptores elípticos de Fourier y análisis canónicos multivariados en las cuatro especies recolectadas en las poblaciones de Camarones (Riohacha-Guajira), Santa Marta (Magdalena) y Rosalía (Desierto de la Tatacoa, Huila). Los resultados mostraron diferencias significativas en la forma de la mancha abdominal entre las especies (P << 0.001). Cada especie conservó su identidad taxonómica. Se observaron diferencias estadísticas (P << 0.001) entre las poblaciones de D. martensis, D. starmeri y D. uniseta. La población de D. aldrichi, de Tatacoa presenta solapamiento con D. martensis lo que sugiere una mayor relación fenotípica entre estas dos especies. Se discuten las implicaciones de estas diferenciaciones.
Palabras clave: Variabilidad morfológica. Morfometría geométrica. Análisis elíptico de Fourier.
Abstract: The abdominal pigmentation of geographically isolated populations of species of the repleta group, D. martensis, D. starmeri and D. uniseta ( martensis cluster), was compared. Our purpose was to detect whether the distance that separated them, together with the environmental heterogeneity, are related to a process of geographic race formation. Moreover, variation in the aforementioned character was compared between D. aldrichi from the mulleri cluster, and species from the martensis-cluster species, attempting to contrast the evolutionary relationships among these species. The outline of the third-abdominal spot was digitalized with elliptic Fourier descriptors and multivariate canonical analysis for the four populations collected from Camarones (Riohacha-Guajira Department), Santa Marta (Magdalena Department) and Rosalía (Tatacoa Desert, Huila Department). Results showed significant differences in the form of the abdominal spot morphology among the species (P << 0.001). Each species preserved its taxonomic identity. Statistically significant differences (P << 0.001) were observed among the populations of D. martensis, D. starmeri and D. uniseta. The population of D. aldrichi from Tatacoa overlapped with D. martensis, which suggests a greater phenotypic relationship between these two species. The implications of these differentiations are discussed.
Key words: Morphological variability. Geometric morphometrics. Fourier elliptical analysis. de razas geográficas (Castañeda 2000; Prieto y De Polanco 2007; Estrada y Prieto 2007; Prieto 2008; De Polanco et al. 2008).
Introducción
El estudio de las características morfológicas en las especies del género Drosophila reviste un gran interés científico, especialmente por los procesos evolutivos y de especiación que se pueden evidenciar en la variación de algunos fenotipos (Laurie et al. 1997; Ordoñez et al. 2004; Mafla 2005). Los polimorfismos intra e interpoblacionales y las divergencias interespecíficas son criterios básicos para evaluar el avance de procesos de formación de razas geográficas y de especiación, particularmente en poblaciones aisladas, habitando en ecosistemas con diferentes características topográficas, edáficas y climatológicas que favorecen dichos acontecimientos evolutivos. El grupo repleta y en especial el conglomerado martensis de Drosophila, han sido evaluados en Colombia, con resultados que permiten plantear que las poblaciones de la Costa Norte y el desierto de la Tatacoa de Drosophila martensis Wasserman y Wilson, 1957, D.starmeri Wasserman y Koepfer, 1973 y D. uniseta Wasserman y Koepfer, 1973, manifiestan variaciones morfológicas inter-poblacionales, importantes en el momento de la consideración del estatusde razas geográficas (Castañeda 2000; Prieto y De Polanco 2007; Estrada y Prieto 2007; Prieto 2008; De Polanco et al. 2008).
Existe una relación entre la variación de algunas características morfológicas como la pigmentación y los procesos evolutivos que se pueden dar en las especies, la cual se traduce en diversidad fenotípica. La morfometría, una de las herramientas fundamentales en el estudio de las ciencias biológicas y en la prueba de muchas hipótesis evolutivas (Adams et al. 2004; Klingenberg y Monteiro 2005), permite cuantificar dicha variación y separar los componentes genético y ambiental del carácter examinado (Rohlf y Marcus 1993; Castañeda 2000; Jaramillo 2000; Jaramillo y Dujardin 2003). En este trabajo se realizó un análisis elíptico de Fourier a la tercera mancha abdominal de las especies: D. martensis, D. uniseta y D.starmeri recolectadas en Riohacha y Santa Marta en la Costa Norte colombiana y en el desierto de la Tatacoa (Huila) también a D. aldrichi Patterson y Crown, 1940, proveniente de la Tatacoa, con el fin de evaluar su variación, dado que la pigmentación abdominal posee un componente genético que contribuye a la adaptabilidad en ambientes cambiantes y que sirve para la distinción entre razas geográficas (Hollocher et al. 2000; Brisson et al. 2004).
Materiales y Métodos
Se utilizaron 50 individuos, 25 machos y 25 hembras vírgenes, de las especies D. martensis, D. starmeri, D. uniseta y D. aldrichi, que se aislaron y mantuvieron en un medio de cultivo modificado del propuesto por Sans de la Rosa et al. (1987), como lo sugieren De Polanco et al. (2005), hasta que cumplieron tres días de haber emergido de la pupa, evitando así la variación resultante del crecimiento en las primeras horas de vida de estos insectos. Los individuos se recolectaron en la localidad de Camarones en Riohacha (11°31' N, 72°55'W) y Santa Marta en la Costa Norte (11°42'N, 74°12'W) y en Rosalía en el desierto de la Tatacoa (3°13'N,75°10'W) (Fig. 1). El establecimiento de un cepario, para mantener las moscas por una generación después de la recolección, se hizo necesario debido a la dificultad que presenta la determinación de los organismos en campo, como consecuencia de su carácter de especies sinmórficas. Para la determinación de las especies, se tomaron las descripciones de Wasserman y Wilson (1957), Strickberger (1962), Vilela (1983) y De Polanco (1998).
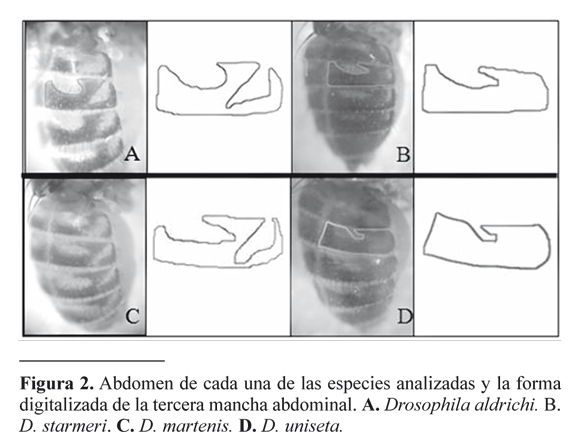
Con el fin de evaluar la forma de la mancha del tercer segmento abdominal se utilizó un análisis geométrico con Series de Fourier Elípticas, el cual se basa en la toma de coordenadas del contorno de la estructura que se analiza o “Outlines” y en el ajuste de los puntos que reproducen gráficamente la forma de la estructura originalmente digitalizada luego de varias repeticiones (Liu et al. 1996; Laurie et al. 1997; De Polanco et al. 2005). Los coeficientes del análisis elíptico de Fourier, producto de las deformaciones o “armónicas” realizadas sobre una elipse inicial, se consignan en una matriz en donde las filas representan los individuos y las columnas cada una de las variables, y finalmente se realiza el análisis multivariado. El método utiliza la forma de la mancha conservando su relación geométrica a lo largo del análisis como una configuración de datos en un espacio invariante con respecto a los efectos de la rotación, la escala y el desplazamiento de la estructura (Lestrel 1997; Adams y Rohlf 2000; Galindo 2004; Adams et al. 2004).
El proceso se inició con la adquisición de las imágenes de la mancha abdominal mediante una cámara digital Digital Colpix 5700 de Nikon unida a un estereoscopio Olympus. Posteriormente se utilizó el programa ACDsee 5.0 para dibujar el contorno de la mancha (Fig. 2) y generar un nuevo archivo que contenga solo el trazo del mismo; a continuación se ejecutaron los programas de la serie Tps, TpsDig y Tpstoefa (http://life.bio.sunysb.edu/morph/), los cuales permiten convertir los puntos del dibujo a coordenadas y crear un archivo compatible con el programa EFAWin respectivamente. De la utilización del programa EFAWin se obtuvo una matriz de datos constituida por los coeficientes generados por ocho armónicas, que representan la forma de la estructura. Finalmente se utilizó el programa Efa-estadística, para convertir los datos de EFAWin a matrices de extensión .TXT que se llevaron a una hoja electrónica para facilitar la aplicación de los programas de análisis estadístico. Las matrices se analizaron con el programa PAST© 1.22 (Hammer et al. 2001), aplicando el análisis canónico discriminante MANOVA-CVA a través de estadísticos como el de Shapiro Wilk`s y Pillai Trace.
Resultados
Las comparaciones inter-específicas mostraron una alta variación en el contorno de la tercera mancha abdominal, presentando una forma particular en cada especie (Fig. 2). Esto se refleja en el análisis canónico discriminante (CVA) de las especies D. martensis, D.starmeri y D. uniseta (Fig. 3), el cual mostró diferencias entre estas (P << 0.05), al igual que el CVA entre las especies del conglomerado martensis y D. aldrichi (P < 0.05) (Fig. 4). En este último se resalta el solapamiento entre D. aldrichi y D . martensis (Fig. 4C).
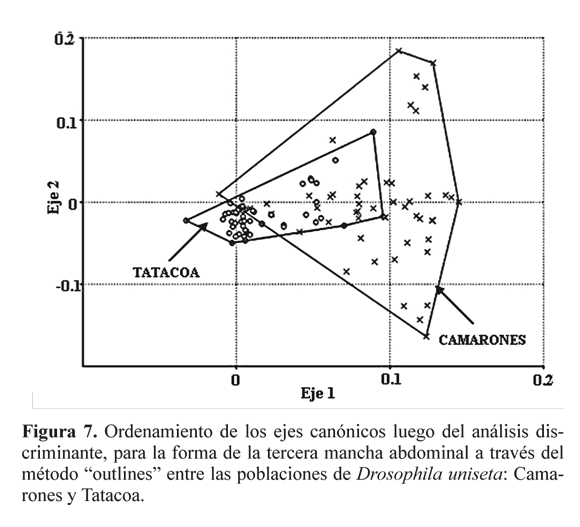
El CVA de todas las agrupaciones muestra diferencias inter- poblacionales. En D. martensis el CVA mostró diferencias entre las poblaciones de Tatacoa, Santa Marta y Camarones (Pλ wilk = 9.647 E-14, Pλ wilk= 6.55 E-14, Pλ wilk = 0.0002) (Figs. 5A, 5B y 5C). La misma prueba en D.starmeri para Tatacoa, Santa Marta y Camarones, permitió establecer diferencias en la forma de la mancha abdominal (Pλ wilk= 8.016E-25, Pλ wilk= 5.028E-6 Pλ wilk= 6.415E-27) (Figs. 6A, 6B y 6C). Las poblaciones de Camarones y el desierto de la Tatacoa en D. uniseta presentan diferencias (Pλ wilk= 3.391E-13), sin embargo los dos agrupamientos se solapan (Fig. 7). De igual modo, los individuos de la población de Tatacoa frente a los de Camarones tienen una escasa variación en la forma de la mancha abdominal. De las tres especies, D. uniseta es la que menor variación refleja.
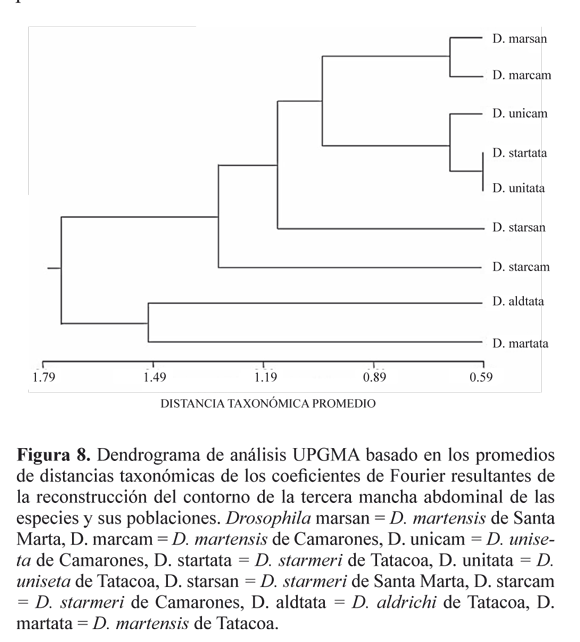
El análisis de agrupamiento UPGMA (Fig. 8), muestra una relación de similitud entre las especies D. martensis y D. aldrichi, al ser comparadas con todas las agrupaciones y confirma la diferencia entre estas dos especies y D. uniseta y D. starmeri. Así mismo, se puede determinar la identidad taxonómica de cada especie y la semejanza existente entre las poblaciones de la Costa Norte en D. martensis.
Discusión
Las especies mantienen su identidad taxonómica. Al compararlas en cada una de las poblaciones, se puede determinar la existencia de una forma base (o primitiva) en la tercera mancha abdominal, evidenciada en el hecho de que existen individuos de cada especie que comparten semejanzas con respecto a esta característica, tal como puede observarse en las intersecciones de las gráficas del MANOVA. El hecho de que las especies sean consideradas gemelas o hermanas, determina la existencia de un ancestro común (Carvajal-Rodríguez et al. 2005), del cual aún pueden compartir muchas características. Por tal razón es probable que estas semejanzas resulten de su estrecha relación filogenética y de la similitud en las condiciones ecológicas que requieren.
La similitud entre las muestras de D. martensis, de la Costa Norte coincide con la semejanza fenotípica y genética, que a nivel enzimático, fue reportada por Ordoñez et al. 2004, en estas localidades. Esta situación es frecuente en las razas de las especies continentales en las que los límites de cada población no son muy precisos observándose en algunos casos zonas de integración que pueden mantener un mínimo flujo de genes entre las poblaciones (García y Kirkpatrick 1997).
La variación inter-poblacional que se manifestó en cada una de las especies a través de la reconstrucción geométrica de la pigmentación abdominal, podría considerarse una evidencia del proceso de formación de razas geográficas, lo que eventualmente puede desembocar en un proceso de microevolución. Teniendo en cuenta el componente genético asociado con la pigmentación en las especies de Drosophila (Brisson et al. 2004; Carbone et al. 2005) y el politipismo aquí encontrado a este nivel, es posible que cada variante en la forma de la mancha sea adaptativa en la población en donde aparece. No obstante, también es posible que las diferencias pobla cionales en las manchas abdominales no sean adaptativas en absoluto y que se deban a mutaciones que generan variación y se han fijado en las poblaciones por deriva genética. A pesar de esto, la existencia de variación particular en cada población, sustentada en cualquiera de las dos hipótesis y en la imposibilidad de ser compartida por el aislamiento geográfico, se ajusta a las primeras etapas del modelo de especiación alopátrica.
Otro aspecto relevante, que vale la pena considerar, es la posibilidad de encontrar algún tipo de selección sexual, asociada con la pigmentación, que se esté reflejando en las preferencias de apareamiento entre individuos de la misma población y especie, lo cual permitiría que, en especies estrechamente relacionadas los apareamientos fueran sesgados, favoreciendo el mantenimiento de las barreras reproductivas (Rojas 2001; Rodriguero et al. 2002; Prud'homme et al. 2006); en este sentido, es probable que las diferencias encontradas en la forma de la mancha abdominal puedan favorecer un reconocimiento de los miembros de la misma especie o población.
La variación morfológica puede estar evidenciando diferentes arquitecturas genéticas y procesos adaptativos microevolutivos en las especies y poblaciones estudiadas de D. martensis, D.starmeri y D. uniseta. De otra parte, el origen de los mecanismos de aislamiento necesarios en la formación de nuevas especies se halla generalmente asociado con la diversificación morfológica y ecológica de las poblaciones. En consecuencia, es posible plantear la existencia de un proceso activo de formación de razas que, eventualmente, puede desembocar en un proceso incipiente de especiación.
Finalmente, sin pretender con los resultados establecer filogenias, es preciso decir que al comparar la pigmentación de D. aldrichi con las demás especies se encontró una mayor similitud con D. martensis. Este resultado es particularmente interesante por que refuerza la hipótesis planteada por De Polanco et al. (2008), quienes plantean que D. aldrichi y D. martensis, actualmente ubicadas en conglomerados diferentes mulleri y martensis respectivamente, deben ser reubicadas juntas en uno solo.
Literatura citada
ADAMS, D. C.; ROHLF, F. J. 2000. Ecological character displacement in Plethodon: Biomechanical differences found from a geometric morphometric study. Proceedings of the National Academy of Sciences USA 97 (8): 4106-4111. [ Links ]
ADAMS, D. C.; ROHLF, F. J.; SLICE, D. E. 2004. Geometric morphometrics: ten years of progress following the'Revolution'. Special issue of the workshop in Rome. Italian Journal of Zoology 71: 5-16. [ Links ]
BRISSON, J.; TEMPLETON, A.; DUNCAN, I. 2004. Population genetics of the developmental gene optomotor-blind (omb) in Drosophila polymorpha: Evidence for a role in abdominal pigmentation variation. Genetics 168: 1999-2010. [ Links ]
CARBONE, M.; LLOPART, A.; DEANGELIS, M.; COYNE, J. 2005. Quantitative trait loci affecting the difference in pigmentation between Drosophila yakuba and D. santomea. Genetics 171: 211-225. [ Links ]
CARVAJAL-RODRÍGUEZ, A.; CONDE-PADÍN, P.; ROLÁNALVAREZ, E. 2005. Decomposing shell form into size and shape by geometric morphometric methods in two sympatric ecotypes of littorina saxatilis. Journal of Molluscan Studies 71: 313-318. [ Links ]
CASTAÑEDA, H. C. 2000. Análisis morfológico comparativo en Drosophilastarmeri de las poblaciones de Camarones (Guajira), Santa Marta (Magdalena) y Tatacoa (Desierto de la Tatacoa, Huila). Bogotá. Tesis Maestría. Universidad de los Andes, Instituto de Genética 89 p. [ Links ]
DE POLANCO, M. M. E. 1998. Estudios cromosómicos comparados de Drosophila repleta (Cepa Siboney, Cuba) vs. Drosophila repleta (Wharton, 1942) y Drosophila martensis (Valledupar, Barrancas y Riohacha), Tesis doctoral, Universidad de los Andes. Tomo I, p. 30-45. [ Links ]
DE POLANCO, M.; ORDOÑEZ, M.; GALINDO, L. 2005. Morfometría del contorno (borde de la forma) con Series de Fourier, paso a paso. Ibagué: Universidad del Tolima, p. 6-25. [ Links ]
DE POLANCO, M. M. E.; PRIETO, R.; GALINDO, L.; LOZANO, L.; ORDÓÑEZ, M. 2008. Análisis morfométrico en especies de Drosophila (Diptera: Drosophilidae) del grupo repleta de ecosistemas semiáridos colombianos. Revista Colombiana de Entomología 34 (1): 105-109. [ Links ]
ESTRADA, A.; PRIETO, R. 2007. Análisis de la variación genética de dos poblaciones de Drosophila (Grupo repleta) provenientes de ecosistemas semiáridos aislados geográficamente (Desierto de la Tatacoa - Huila y Valle del Patía Nariño) mediante cruces interpoblacionales y morfometría del Aedeago. Revista de la Asociación Colombiana de Ciencias Biológicas. Memorias XLII Congreso Nacional de Ciencias Biológicas 19 (1): 1-188. [ Links ]
GALINDO, L. F. 2004. Evaluación del estatus sistemático de Heliconius tristero Brower (Lepidoptera: Nymphalidae): datos morfométricos, observaciones ecológicas y experimentos de cruces genéticos. Tesis de Maestría en Biología, Universidad de los Andes, Bogotá, Colombia, pp. 18-30. [ Links ]
GARCÍA, R. G.; KIRKPATRICK, M. 1997. Genetic models of adaptation and gene flow in peripheral populations. Evolution 51 (1): 21-28. [ Links ]
HAMMER, O.; HARPER, D.; RYAN, P. 2001. Paleontological statistics software package for education and data analysis. Paleontología Electrónica 4 (1): 9. [ Links ]
HOLLOCHER, H.; L, JULIET; G. ERIC. 2000. Evolution of abdominal pigmentation differences across species in the Drosophila dunni subgroup. Evolution 54: 2046-2056. [ Links ]
JARAMILLO, N. 2000. Caracterización en tamaño y forma de los caracteres métricos y su interés en los estudios poblacionales aplicados a los Triatominae. Universidad de Antioquia, Tesis de doctorado en Ciencias Básicas Biomédicas, Medellín. 145 p [ Links ]
JARAMILLO, N.; DUJARDIN, J. P. 2003. Análisis morfométrico: Significado biológico del tamaño y la conformación, pp. 151- 166. In: Guhl, F.; Schofield, C. J. (Eds.). Proceedings Fourth International Workshop on Population Genetics and Control of Triatominae, Cartagena de Indias (ECLAT 4). CIMPAT, Universidad de los Andes, Corcas Editores Ltda., Bogotá, Colombia. [ Links ]
KLINGENBERG, P. C.; MONTEIRO, L. R. 2005. Distances and directions in multidimensional shape spaces: Implications for morphometric applications. Systematic Biology 54 (4): 678-688. [ Links ]
LAURIE, C.; TRUE, J. R.; LIU, J.; MERCER, J. M. 1997. An introgression analysis of quantitative trait loci that contribute to a morphological difference between Drosophila simulans and D. mauritiana. Genetics 145: 339-348. [ Links ]
LESTREL, P. (Ed.). 1997. Fourier descriptors and their applications in Biology. Cambridge Univ. Press. 466 p. [ Links ]
LIU, J.; MERCER, J. M.; STAM, L.; GIBSON, G.; ZENG, Z.; LAURIE, C. 1996. Genetic analysis of a morphological shape difference in the male genitalia of Drosophila simulans and D. mauritana. Genetics 142: 1129-1145. [ Links ]
MAFLA, A. 2005. Ciclos de vida y componentes de la aptitud de Drosophila inca y D. yangana (Diptera, Drosophilidae). Iheringia Série Zoología 95 (1): 89-91. [ Links ]
ORDOÑEZ, M.; ARENA, A.; BETANCOURT, A.; POLANCO, M. de. 2004. Análisis electroforético de las relaciones evolutivas en las especies de Drosophila (Diptera: Drosophilidae) del enjambre martensis: D. martensis, D.starmeri y D. uniseta. Revista Colombiana de Entomología 30 (1): 65-74. [ Links ]
PRIETO, R. 2008. Análisis morfométrico de cuatro especies del grupo repleta: D. martensis, D. starmeri, D. uniseta y D. aldrichi de dos ecosistemas semiáridos, desierto de la Tatacoa y Costa Norte colombiana. Tesis de Maestría. Universidad del Tolima. 110 p. [ Links ]
PRIETO, R.; DE POLANCO M. M. 2007. Análisis morfométrico con series de Fourier del ala en 4 especies del grupo repleta de Drosophila provenientes de dos ecosistemas semiáridos colombianos. Revista de la Asociación Colombiana de Ciencias Biológicas. Memorias XLII Congreso Nacional de Ciencias Biológicas 19 (1): 1-188. [ Links ]
PRUD'HOMME, B.; GOMPEL, N.; ROKAS A. KASSNER, V.; WILLIAMS, T.; YEH S.; TRUE J.; SEAN, B. 2006. Repeated morphological evolution through cis-regulatory changes in a pleiotropic gene. Nature 440: 1050-1053 [ Links ]
RODRIGUERO, M.; VERA, M. T.; CALCAGNO, G.; ALLINGHI, A.; CLADERA, J.; VILARDI, J. C. 2002. Selección sexual sobre el fenotipo multivariado en la mosca de los frutos sudamericana Anastrepha fraterculus (Diptera Tephritidae). 31 Congreso Argentino de Genética 68 p. [ Links ]
ROHLF, F. J.; MARCUS, L. F. 1993. A revolution in morphometrics. Trends in Ecology and Evolution 8 (4): 129-132. [ Links ]
ROJAS, M. A. P. 2001. Análisis del aislamiento reproductivo en las especies cactofílicas del enjambre martensis, en dos regiones Colombianas (Costa Norte y Desierto de la Tatacoa). Bogotá. Tesis Maestría. Universidad de los Andes, Instituto de Genética. 105 p. [ Links ]
SANS DE LA ROSA, M; MORENO-ROSSI, A.; BUSTOS, E. 1987. Nuevo medio de cultivo para las especies de Drosophila starmeri, Drosophila uniseta, Drosophila martensis, y Drosophila venezolana del grupo repleta del enjambre martensis, recientemente colectadas en los desiertos de la Guajira Colombiana. Evolución Biológica 2: 305-306. [ Links ]
STRICKBERGER, M. N. 1962. Experiments in genetics with Drosophila. John Wiley; New York. 345 p. [ Links ]
VILELA, R. C. A. 1983. Revision of the Drosophila repleta species group (Diptera, Drosophilidae). Revista Brasileira de Entomologia 27 (1): 78-81. [ Links ]
WASSERMAN, M.; WILSON, F. D. 1957. Further studies on the repleta group. University of Texas Publications 5721: 132-156. [ Links ]
WASSERMAN, M.; KOEPFER, H. R. 1973. Cytogenetics of the South American Drosophila mulleri complex: the martensis cluster, more interespecific shaving of inversions. Genetics 93: 935- 946. [ Links ]

