










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488On-line version ISSN 2665-4385
Rev. Colomb. Entomol. vol.36 no.1 Bogotá Jan./June 2010
Abundancia de colémbolos (Hexapoda: Collembola) y parámetros edáficos de una selva baja caducifolia
Abundance of springtails (Hexapoda: Collembola) and edaphic parameters from a deciduous dry forest
JOSÉ ANTONIO GÓMEZ-ANAYA1, JOSÉ G. PALACIOS-VARGAS2 y GABRIELA CASTAÑO-MENESES2,3
1 Técnico Departamento de Entomología, Instituto de Ecología, A.C. AP 63, 91000, Xalapa, Veracruz, México. antonio.gomez@inecol.edu.mx.
2 Investigador Ecología y Sistemática de Microartrópodos, Departamento de Ecología y Recursos Naturales, Facultad de Ciencias, Universidad Nacional Autónoma de México, 04510, México, D.F. troglolaphysa@hotmail.com.
3 Técnico Unidad Multidisciplinaria de Docencia e Investigación, Facultad de Ciencias, Universidad Nacional Autónoma de México, Campus Juriquilla, Boulevard Juriquilla 3001, C.P. 76230, Querétaro, Querétaro, México. gcm@hp.fciencias.unam.mx.
Recibido: 22-oct-2009 - Aceptado: 17-abr-2010
Resumen: Se describen y comparan los patrones de abundancia de colémbolos de hojarasca y suelo de dos cuencas hidrológicas de la Estación de Biología Chamela (EBCh, UNAM) en Jalisco, México, con base en colectas mensuales durante un año. Esta localidad, donde el tipo de vegetación dominante es la selva baja caducifolia, presentó un año de precipitación pluvial atípico durante los muestreos realizados, con la mayor cantidad de lluvia durante enero. El muestreo de ambos biotopos fue aleatorio, los núcleos fueron de igual tamaño (95 cm2) y se procesaron in situ mediante embudos de Berlese. Adicionalmente, se cuantificaron diferentes variables edáficas. Se analizó el efecto de los factores cuenca, biotopo y estacionalidad sobre la abundancia de las familias de Collembola. Mediante análisis de conglomerados (AC) se exploran las relaciones faunísticas entre los ensamblajes, y mediante análisis de correspondencia canónica (ACC), las relaciones entre los ensamblajes y las variables fisicoquímicas del suelo. Se obtuvieron 24.253 colémbolos pertenecientes a trece familias. Las más abundantes fueron Isotomidae, Entomobryidae, Hypogastruridae y Sminthuridae. El AC mostró mayor similitud entre biotopos que entre cuencas, mientras que en el ACC la variable más importante para explicar la distribución y abundancia de los Collembola fue la humedad.
Palabras clave: Análisis de Conglomerados. Análisis de Correspondencia Canónica. Chamela. México. Patrones de abundancia.
Abstract: The abundance patterns of leaf litter and soil springtails (Hexapoda: Collembola) from two hydrological watersheds of the Chamela Biological Station (EBCh, UNAM) in Jalisco, Mexico are described and compared based on monthly sampling over one year. This locality, where the dominant vegetation is tropical dry forest, had an atypically high rainfall during the sampling period, with the greatest amount of rain during January. Sampling in both biotopes was random, the areas sampled were of the same size (95 cm2), and these were processed in situ using Berlese funnels. In addition, different edaphic characteristics were quantified. The effect of watershed, biotope and seasonality factors on the abundance of Collembola families was analysed. Using Cluster Analysis (CA), the faunistic relationships between the assemblages were explored, and using Canonical Correspondence Analysis (CCA) the relationships between assemblages and the physico-chemical variables of the soil. A total of 24.253 Collembola were obtained, belonging to 13 families. The most abundant were Isotomidae, Entomobryidae, Hypogastruridae and Sminthuridae. The CA showed more similarity between biotopes than between hydrological watersheds, while in the CCA the most important variable to explain the distribution and abundance of the Collembola was humidity.
Key words: Cluster analysis. Abundance patterns. Canonical Correspondence Analysis. Chamela. Mexico.
Introducción
Los microartrópodos son un componente importante de la fauna del suelo y de otros medios (Palacios-Vargas et al. 2007). En general, los grupos del suelo mejor representados por su diversidad y abundancia son los ácaros oribátidos y los colémbolos (Johnston 2000). Su importancia radica en que su actividad puede influir en la formación y fertilidad de los suelos afectando de esta manera en el nivel de producción primaria del ecosistema (Najt 1973; Tanaka et al. 1978; Faber 1992) y en el ciclo de los nutrientes (Heneghan y Bolger 1998). Por lo anterior, es importante el estudio de su diversidad, distribución, abundancia, biomasa y el carácter de su actividad.
Se sabe que los microartrópodos están relacionados con diferentes factores del medio tales como pH (Loranger et al. 2001), materia orgánica (Merilä y Ohtonen 1997) disponibilidad de nutrientes (Bird et al. 2000), tipo de humus (Hagvar 1982,1983; Hagvar y Abrahamsen 1984; Ponge et al. 1986; Schaefer y Schauermann 1990; Chagnon et al. 2001; Cassagne et al. 2003) y cubierta vegetal (Paquin y Coderre 1997). Pero se sabe poco acerca de la relación entre los Collembola y las diferentes características químicas del suelo (Hagvar y Abrahamsen 1984; Chagnon et al. 2000a; Loranger et al. 2001). Dentro de la ecología, el tema principal ha sido el de los factores que controlan su abundancia; autores como Takeda (1981), Holt (1985), Arbea y Jordana (1985), Gamma et al. (1991) y Díaz y Najt (1990), entre otros, indican que la reducción en la humedad durante la estación seca es lo que más afecta a las poblaciones edáficas. Por otro lado, el efecto del uso del suelo sobre la fauna de colémbolos se ha documentado en algunas ocasiones, señalando que las prácticas agrícolas producen una ruptura total del equilibrio de las comunidades edáficas (Huhta et al. 1967; Izarra y Boo 1980; Reyes y Breceda 1985; Jordana et al. 1987).
No obstante, hay trabajos que señalan que muchas especies de colémbolos son sensibles a los cambios en las condiciones edáficas y han sido utilizados como bioindicadores de la salud del suelo (Prat y Massoud 1980; Pinto et al. 1997; Kovác y Miklisová 1997; Greenslade 1997; Frampton 1994, 1997). En este sentido, su utilidad ha sido registrada tanto a nivel de especie (Bengtsson et al. 1983, 1985a, b; Dunger 1986) como familia (Waikhom et al. 2006; Baretta et al. 2008).
Los Collembola pueden resultar más abundantes en ciertos tipos de suelo (Olejniczak 2000). Su diversidad supera actualmente las 7.000 especies a nivel mundial (Palacios- Vargas 2000), siendo mayor hacia los trópicos. En México el conocimiento que se tiene de estos organismos es principalmente taxonómico. Relativamente pocos estudios han aportado información ecológica, de ellos podemos mencionar el de Lavèlle et al. (1981); Palacios-Vargas (1985); Villalobos, (1989, 1990); Miranda y Palacios-Vargas (1992); Mendoza (1995); Palacios-Vargas et al. (1998); Mendoza et al. (1999) y Cutz-Pool et al. (2007). La actualización más reciente al catálogo de los Collembola de México registra más de 600 especies (Palacios-Vargas 1997).
Desafortunadamente, resta mucho por hacer en relación al conocimiento taxonómico de un grupo tan diverso como este (Chagnon et al. 2000b) y, tal vez ante la alta tasa de pérdida de los bosques tropicales, muchas especies nunca serán conocidas (Janzen 1988; Trejo y Dirzo 2000). El objetivo del presente estudio es describir y comparar los patrones de abundancia de las familias de Collembola de dos cuencas similares en cuanto a su producción anual de hojarasca; así como el explorar la relación entre la abundancia del grupo y una serie de variables edáficas.
Materiales y Métodos
Área de estudio. La Estación de Biología Chamela (EBCh) se localiza en la Costa del Pacífico mexicano (19°30’N, 105°03’W), en una región montañosa de relieves dominados por lomeríos y planicies aluviales (Fig. 1). Comprende una superficie de 3.200 hectáreas y es parte integral de la Reserva de la Biosfera Chamela-Cuixmala (RBCC), que actualmente, mediante un convenio entre la UNAM y la Fundación Ecológica de Cuixmala, A.C., protege a más de 13.000 hectáreas de selvas y humedales. Su clima es cálido húmedo con una temperatura promedio de 22 a 26°C y precipitación total anual que varía de 400 a 1.300 mm distribuyéndose en un periodo de cuatro a seis meses, por lo que existe una marcada estacionalidad (Trejo y Dirzo 2000). El periodo de lluvias comprende de julio a octubre y el de secas de noviembre a junio (Maass et al. 2002). La precipitación promedio anual es de 748 mm en la zona de lomeríos y de 782 mm en Cuixmala (García-Oliva et al. 1991). La selva se caracteriza por su diversidad de flora y fauna y por su elevado número de especies endémicas (Lott et al. 1987). La tenencia de la tierra en la Reserva corresponde principalmente a dos propiedades privadas: una de la Universidad Nacional Autónoma de México (UNAM), y otra de la Fundación Ecológica Cuixmala A.C. El nombre del área protegida deriva del pequeño poblado de Chamela, que fue un importante puerto hace varias décadas, y del Río Cuixmala, límite natural del área y el de mayor caudal en esta región (Noguera et al. 2002).
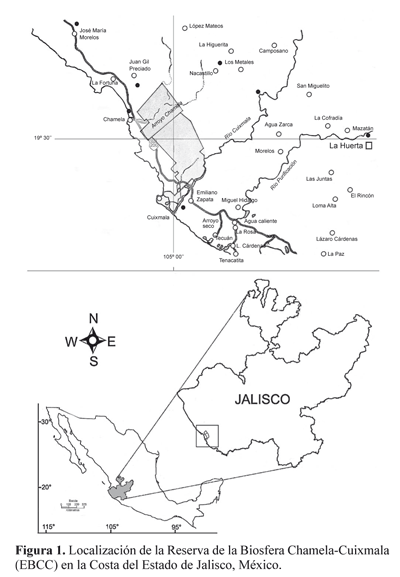
Muestreo. El muestreo de ambos biotopos fue aleatorio, los núcleos de suelo fueron de igual tamaño (95 cm2) y fueron procesados in situ mediante una batería de embudos de Berlese en la Estación de Biología de Chamela (EBCh) durante tres días sin fuente de luz, y los siguientes tres con una bombilla de 25watts por embudo. La fauna extraída se fijó en etanol 75% y fue transportada al Laboratorio de Ecología y Sistemática de Microatrópodos de la Facultad de Ciencias- UNAM. Los colémbolos fueron separados del resto de la fauna y cuantificados a nivel de familia, para lo cual fue necesario realizar numerosas preparaciones semipermanentes en líquido de Hoyer, de acuerdo con la técnica de Christiansen y Bellinger (1980-81).
Adicionalmente, se cuantificaron 17 variables edáficas que fueron medidas en las mismas muestras de suelo de las cuales se extrajo la fauna. Este trabajo se realizó en el Laboratorio de Edafología “Nicolás Aguilera” de la Facultad de Ciencias de la UNAM. Las variables fueron: los porcentajes de fracción fina < 2 mm (FF) y gruesa > 2 mm (FG) mediante el tamizado en seco con malla de abertura 2 mm; arena (ARE); limo (LIM); arcilla (ARC); carbono (C) y materia orgánica (MO) mediante el método modificado de Walkley y Black (Jackson 1982); humedad (HUM) como porcentaje calculado por regla de tres en relación al peso húmedo y peso seco de cada muestra. Densidad real (DR) con un picnómetro y densidad aparente (DA) por el método de Probeta; el porcentaje de porosidad (POR) se calculó mediante el cociente DA/DR. El pH se determinó en una solución de suelo y agua destilada de 1:2.5 con un potenciómetro Corning Modelo 7; el magnesio (Mg) y calcio (Ca) mediante el método de extracción con acetato de amonio 1N pH7 y valoración con varsenato EDTA 0.02N (Jackson 1982); sodio (Na) y potasio (K) por el método de extracción de acetato de amonio 1N pH 7 y determinación con fotómetro de llama Corning 400 (Jackson 1982), y la capacidad de intercambio catiónico (CICT) por el método de saturación con CaCl2, lavado con etanol al 96% y elusión con NaCl 1N pH7 y valoración con varsenato (EDTA 0.02N) y negro de ericromo T (Jackson 1982).
Análisis estadístico de datos. Para comparar la constitución edáfica del suelo de ambas cuencas y de las temporadas de lluvias y secas se utilizó una prueba t2 de Hotelling, las diferencias, cuando se presentaron, se detectaron mediante los respectivos valores de las pruebas de t incluidos en la rutina. Por otro lado, la abundancia de colémbolos por muestra fue transformada como log (x+1) (Vegter et al. 1988), tanto para los conteos totales, como para aquellos de cada familia, con lo cual se logró normalidad e igualdad de varianzas. Un análisis de varianza de tres vías (ANOVA) fue utilizado para probar el efecto de los factores cuenca, biotopo y mes (tiempo) sobre la abundancia de Collembola. De haber efecto significativo de algún factor o en algún efecto cruzado se procedió a realizar los contrastes de Bonferroni. Asimismo, se calcularon los coeficientes de correlación de Kendall entre las abundancias de las familias y las variables edáficas. Los gráficos y tablas se generaron a partir de los datos sin transformar. Los análisis se realizaron con el paquete STATISTICA ver. 7.1 (StatSoft 2006).
Las relaciones faunísticas de los ensamblajes fueron exploradas mediante la técnica de análisis de conglomerados (AC), para lo cual se construyó una matriz de similitud de Bray-Curtis (similitud cuantitativa), y se utilizó como método de amalgamiento la unión de pares por promedio (UPGMA). Este análisis fue realizado con el programa PC-ORD 5 (McCune y Grace 2002).
Finalmente, analizamos las relaciones que entre las abundancias de Collembola y las variables fisicoquímicas del suelo por medio de un análisis de correspondencia canónica (ACC). La significancia estadística del primer eje y del conjunto de ejes canónicos se comprobó mediante 499 pruebas de permutaciones no restringidas de MonteCarlo. Este análisis fue realizado con el programa CANOCO 4.5 para Windows (ter Braak y Smilauer 1998).
Resultados
Desde el punto de vista edáfico, ambas cuencas resultaron estadísticamente diferentes (T2= 112,9, F17, 176 = 6,1, P < 0.001), sin embargo los valores de la prueba de t evidenciaron que algunas variables fueron iguales entre las cuencas (Tabla 1). De la misma forma existen diferencias edáficas entre las temporadas de lluvias y secas (T2=172,1 F17, 176 = 9,28, P < 0.0001). En este último caso las diferencias más notables estuvieron en las proporciones de fracción gruesa, fina, arena, limo y arcilla, así como en el contenido de humedad que fue en lluvias casi el doble del de la época seca.
Estructura general de los ensamblajes de Collembola. Se recolectaron 24.243 colémbolos pertenecientes a trece familias (Tabla 2). En el biotopo hojarasca se colectaron 10.920 individuos, que representan el 45% del total, siendo la abundancia de ambos ensamblajes de la hojarasca prácticamente igual: 5.493 individuos para la cuenca 1 y 5.427 individuos para la cuenca 4. En el suelo se recolectaron en total 13.323 (55%) colémbolos, de los cuales 6.918 fueron de la cuenca 1 y 6.405 de la cuenca 4. Las familias dominantes fueron Isotomidae (29,3%) y Entomobryidae (28,9%), los Sminthuridae sólo contabilizaron el 16,8% de la abundancia total (Tabla 2).
En ambos biotopos de la cuenca 1 se registraron 12 familias. Los Paronellidae (Salina spp.) sólo se registraron accidentalmente de la hojarasca de la cuenca 4. En la hojarasca de la cuenca 1 (HC1) las familias dominantes fueron Isotomidae (5,9%), Sminthuridae (5,2%) y Entomobryidae (4,8%); en la hojarasca de la cuenca 4 (HC4) lo fueron Isotomidae (8%), Sminthuridae (6,3%) y Entomobryidae (4%); en el suelo de la cuenca 1 (SC1) Entomobryidae (10,4%), Isotomidae (6,6%) y Sminthuridae (3%) y, finalmente, en el suelo de la cuenca 4 (SC4) fueron Entomobryidae (9,7%), Isotomidae (8,9%) y las familias Hypogastruridae y Sminthuridae que tuvieron un porcentaje similar (2,4% y 2,3%, respectivamente). La composición porcentual de las familias en cada ensamblaje (Fig. 2) mostró mayor similitud entre biotopos que entre cuencas.
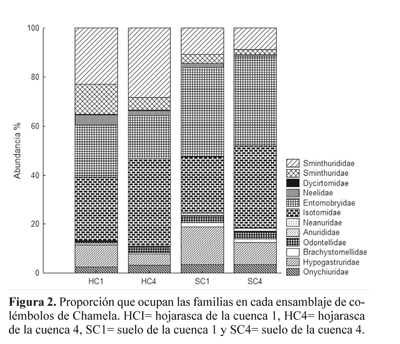
La densidad media (DM) de colémbolos en HC1 fue de 4.818 ind/m2 y para HC4 fue de 4.761 ind/m2. En el SC1 la DM fue de 6.068 ind/m2, mientras que en SC4 fue de 5.618 ind/m2. En general, los Isotomidae (6.233 ind/m2), Entomobryidae (6.142 ind/m2), Sminthuridae (3.570 ind/m2), Hypogastruridae (2.107 ind/m2) y Sminthurididae (1.166 ind/m2) presentaron las mayores densidades (Tabla 2).
Abundancia. La abundancia de colémbolos muestra efecto significativo de la cuenca (F1, 432 = 7,9, P = 0,005), del biotopo (F1, 432 = 120,6, P < 0,05), del mes de captura (F11, 432 = 59,15, P < 0,05) y del efecto cruzado biotopo*mes (F11, 432 = 6,62, P < 0,05), pero no de los efectos cruzados biotopo*cuenca (F1, 432 = 1,1, P = 0,30), cuenca*mes (F11, 432 = 0,97, P = 0,47) y cuenca*biotopo*mes (F11,432 = 1,14 , P = 0,33). De esta forma, es mayor la abundancia, y por ende la densidad, de colémbolos en la cuenca 1 (12.411 colémbolos, 5.443 ind/m2) que en la cuenca 4 (11.832 colémbolos, 5.190 ind/m2). También es estadísticamente mayor la densidad de colémbolos en suelo (5.843 ind/m2) que en hojarasca (4.789 ind/m2). La significancia en el efecto cruzado biotopo*mes implica que existe diferencia en la abundancia registrada entre los biotopos hojarasca y suelo en algunos meses, independientemente de la cuenca que se trate. Este efecto presentó un patrón bien definido, donde la abundancia promedio en el suelo es mayor a lo largo del año (excepto en enero). Las diferencias evidenciadas por los contrastes de Bonferroni corroboraron que la abundancia promedio mensual en suelo es superior a la abundancia promedio mensual de la hojarasca de cualquier mes. Por otro lado, no hubo diferencia en la abundancia de ninguna familia entre las cuencas (P = 0,68, sin considerar a los Paronellidae en esta comparación), mientras que entre los biotopos, los Hypogastruridae, Brachystomellidae, Anurididae, Neanuridae y Entomobryidae fueron más abundantes en el suelo.
Fluctuación de la abundancia de las familias. Los patrones de abundancia de los Hypogastruridae fueron erráticos en los cuatro ensamblajes y sólo mostraron picos de mayor abundancia en el suelo de ambas cuencas después de las lluvias atípicas de enero (Fig. 3A). Los patrones de los Entomobryidae fueron similares y mostraron mayor abundancia en ambos ensamblajes del suelo; también similares a lo largo del año y mostraron menor abundancia en hojarasca. En diciembre y enero ocurrieron las menores abundancias de manera simultánea en los cuatro ensamblajes (Fig. 3B). Los Isotomidae mostraron patrones similares en los cuatro biotopos; en diciembre todos los ensamblajes mostraron disminución en la abundancia, pero luego en enero se incrementó notablemente (Fig. 3C). Los Onychiuridae también incrementaron su abundancia durante enero principalmente (Fig. 3D). Otras familias como los Brachystomellidae fueron más abundantes en enero y febrero y sus menores abundancias ocurrieron de marzo a junio; los Odontellidae tuvieron su mayor abundancia entre septiembre y febrero, exceptuando diciembre, el resto de los meses presentaron abundancias mínimas; los registros de los Anurididae fueron mayores entre julio y febrero, de marzo a junio no hubo representantes de esta familia; los Neanuridae fueron más abundantes durante julio y febrero, exceptuando diciembre, y con menores registros entre marzo y junio; los Neelidae (Megalothorax minimus Willem, 1900) tuvieron mayores registros entre octubre y febrero, exceptuando diciembre, de marzo a junio su abundancia fue nula; los Dicyrtomidae incrementaron notablemente su abundancia en enero y febrero, también existió registro en septiembre y octubre, el resto de los meses no hubo representantes de esta familia; los Sminthurididae se registraron principalmente en enero, los muestreos restantes no los registraron. Finalmente, los Sminthuridae mostraron su mayor incremento en noviembre y enero, su registro fue mínimo de julio a octubre.
Dominancia temporal. Varias familias están bien representadas en los ensamblajes de hojarasca durante los meses de recolección (Figs. 4A, B) especialmente los Entomobryidae e Isotomidae dominan ampliamente en el suelo a través del año. Los ensamblajes de suelo (Figs. 4C, D) guardan una mayor similitud en su dominancia temporal.
Análisis de conglomerados. Se conforman dos grupos a la mayor distancia (Fig. 5). Sin embargo, los ensamblajes mensuales son mixtos, esto es, los conforman hojarasca y suelo de diferentes meses. Esto supone que no existe un patrón de relaciones faunísticas definido en esta dimensión. El establecimiento de ensamblajes tan diferentes de ambas cuencas y ambos biotopos a través del tiempo no permite establecer claramente los grupos. Sin embargo, de manera general es posible apreciar que son más parecidos los ensamblajes entre biotopos iguales (Fig. 6). Resultan más similares ambos ensamblajes de suelo que los de hojarasca, sin duda, el suelo es más estable, mientras que la hojarasca puede acumularse de manera diferencial en ambas cuencas y permitir la conformación de ensamblajes un poco más diferenciados.
Variables del suelo y colémbolos. Casi todas las familias se correlacionaron positivamente con la humedad del suelo, excepto Hypogastruridae (Tabla 3); sin embargo, la abundancia de esta familia se correlacionó positivamente con la porosidad, materia orgánica, carbono, magnesio y potasio, y negativamente con las densidades aparente y real y con el pH. Los coeficientes de correlación significativos entre abundancia y humedad oscilaron entre 0,16 (Neanuridae) y 0,84 (Isotomidae). La abundancia de los Entomobryidae se correlacionó significativamente (P < 0,05) con la mayoría de las variables edáficas. Siendo los más marcados, además de la humedad, la densidad aparente, arcilla, capacidad de intercambios catiónico, porosidad, materia orgánica, carbono y calcio.
Análisis de correspondencia canónica. Los resultados del análisis de correspondencia canónica fueron globalmente significativos (traza = 0,54, F = 3,80, P = 0,002, Tabla 4). Los primeros tres ejes ofrecieron una buena solución a la ordenación de las variables fisicoquímicas y de las familias de Collembola ya que de la variabilidad total en los datos de abundancia de las familias (inercia= 2,35) fue explicable al 91,6% mediante el conjunto de dichos ejes. El primer eje canónico fue también estadísticamente significativo (eigenvalue = 0,29, F=29,84, P = 0,002).
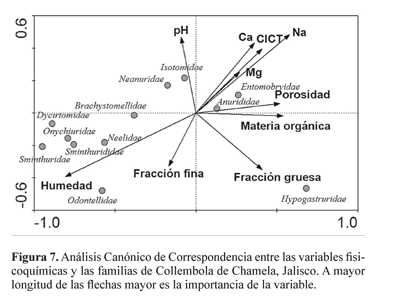
En general, en el ACC las variables humedad y sodio (Na) parecen tener mayor importancia en cuanto a la distribución de las familias de Collembola (Fig. 7). Sin embargo, las familias de Symphypleona, así como Odontellidae, Onychiuridae y Brachystomellidae, tienden a asociarse bien con la humedad y la fracción fina. Por su parte los Hypogastruridae se asocian más con la fracción gruesa del suelo. Finalmente, otras familias como Isotomidae y Neanuridae se asocian más con el pH.
Discusión y conclusiones
Aspectos generales. Los Collembola son un grupo dominante de la fauna edáfica en Chamela, y junto con los ácaros pueden conformar más del 90% de la abundancia de los artrópodos (Palacios-Vargas et al. 2007); igualmente, en el dosel de la misma selva pueden representar hasta el 95% de la abundancia (Palacios-Vargas et al. 1998) donde conforman un ensamblaje contrastante en cuanto a riqueza y composición con los que se establecen en suelo y hojarasca (Palacios- Vargas y Gómez-Anaya 1993).
Las familias dominantes y más constantes a través del año, tanto en hojarasca como en suelo, fueron Entomobryidae e Isotomidae. Mientras que otras, como los Paronellidae, tuvieron registros esporádicos y bien puede ser considerada como una familia rara en suelo y hojarasca de esta región, a pesar de que en el dosel de la misma selva puede conformar más del 90% de la abundancia de los Collembola (Palacios- Vargas et al. 1998).
Las familias Sminthurididae, Sminthuridae, Onychiuridae y Brachystomellidae, presentaron sus mayores registros en periodos definidos y al parecer como una respuesta positiva a la precipitación atípica registrada en enero. A diferencia de ello, los Entomobryidae disminuyeron notablemente su abundancia durante enero. Probablemente como la mayoría de ellos son formas epiedáficas, fueron removidos del suelo con las lluvias, y su registro se incrementó nuevamente en febrero (Fig. 3B). Finalmente, los Hypogastruridae no tuvieron relación alguna con los cambios hídricos.
Densidad. La densidad registrada en este estudio es superior a la que reportan Lavèlle et al. (1981) de 7.025 ind/m2 para el suelo de la selva baja caducifolia de la región de Laguna Verde, Veracruz durante la época de lluvias, y también fue superior a la reportada por Waikhom et al. (2006) de 180x102 ind/m2 en los 10 primeros centímetros del suelo de un bosque subtropical en Phayeng, Manipur (India). En el caso de las familias, las de mayor densidad para Chamela fueron Isotomidae, Entomobryidae, Sminthuridae, Hypogastruridae y Sminthurididae (Tabla 2). La densidad de los Isotomidae de Chamela es similar a la registrada por Lavèlle et al. (1981) de 5.453 ind/m2. Sin embargo, en el caso de los Entomobryidae la densidad de Chamela es mucho mayor que la de Laguna Verde (1.754 ind/m2) y también mayor que la de los Onychiuridae (333 ind/m2) de la misma localidad. Como contraste, la densidad de los Isotomidae de Chamela es mucho menor a la reportada por Waikhom et al. (2006) de 389x102 ind/m2, también menor a la de Hypogastruridae (222x102 ind/m2) y Brachystomellidae (16x102 ind/m2), pero es parecida a la de los Sminthuridae (46x102 ind/m2).
Análisis de conglomerados. Cuando se incluyen los ensamblajes mensuales por biotopo y cuenca en el análisis de conglomerados (AC), los grupos que se forman son mixtos tanto en localidad como en mes (Fig. 5). Lo anterior supone que no existe un patrón bien definido de relaciones faunísticas. Por otro lado, cuando no se utilizan los ensamblajes mensuales y se usan sólo los cuatro ensamblajes generales de las cuencas y biotopos (Fig. 6), se aprecia que los ensamblajes de suelo son un poco más similares (BC = 87,47%) que los de hojarasca (BC = 82,54%), mientras que la menor similitud se localiza entre hojarasca de la cuenca 1 y suelo de la cuenca 4 (BC = 68,17%). La similitud entre cuencas parece mayor que la similitud entre biotopos.
Variables del suelo y colémbolos. Casi todas las familias respondieron positivamente a la humedad del suelo, excepto Hypogastruridae (Tabla 3). La correlación positiva con la humedad sugiere que a mayor humedad edáfica mayor será la abundancia. La respuesta de los Isotomidae fue la más fuerte (Fig. 3C) ya que tuvieron el mayor coeficiente de correlación. Respecto a la relación de la abundancia con otras variables edáficas, algunas familias parecen relacionarse negativa o positivamente con diferentes variables y aunque los coeficientes de correlación provienen de un número relativamente grande de observaciones (N= 240), los valores en general son débiles, por lo que su relación no es clara. También existe la posibilidad de que el efecto de estas variables pudiera ser indirecto, afectando esencialmente la producción de hifas (hongos) que es la fuente principal de su alimento. La inclusión de otras variables, como la cantidad y contenido de esporas e hifas en el tracto digestivo de los colémbolos y en el suelo, es recomendable en futuros estudios para esclarecer estas hipótesis.
En el ACC la mayor influencia sobre la abundancia de las familias de Collembola fue dada por la humedad. Este factor ha sido mencionado en varias ocasiones (Salamon et al. 2004). La estacionalidad al parecer también influye en cuanto a la incidencia de los Collembola, sin duda debido a que el régimen hídrico del suelo depende de la temporada de lluvias y a su vez de las características propias del suelo y su capacidad para retener la humedad por determinados periodos. La cubierta vegetal, así como la cubierta de hojarasca o mantillo, pueden jugar papeles importantes en cuanto al establecimiento y dispersión de los colémbolos al evitar la pérdida de humedad.
Estacionalidad. Como caso excepcional, la familia Hypogastruridae, donde las especies más abundantes fueron Xenylla humicola Fabricius, 1780 y Tafallia insulares Bonet, 1947, no mostró asociación ni correlación significativa con la humedad (Fig. 7, Tabla 4). Se sabe que algunas especies de Xenylla son formas tolerantes a los ambientes secos (Hagvar 1982). En contraste, otras familias, como Sminthurididae tienen su única incidencia, y por ende su mayor abundancia, localizada en los periodos de mayor precipitación. Otras, como Isotomidae, con mayor riqueza que Sminthurididae, Sminthuridae y Onychiuridae, presentaron también su mayor abundancia en los picos de mayor precipitación (Fig. 3).
Los Entomobryidae, generalmente conformados por formas epígeas parecen tener un patrón definido entre julio y noviembre y una respuesta negativa en enero, cuando hubo un caso atípico de precipitación, y luego una recuperación en febrero, principalmente en suelo (Fig. 3B). A diferencia de otras familias, la respuesta de los Entomobryidae a la precipitación es múltiple, debido a que en su composición se incluyen nueve especies. La mayoría de los entomóbridos suelen trepar a la vegetación arbustiva, como lo demuestran las capturas de Entomobrya (Drepanura) ca. californica, Seira bipunctata (Packard, 1873), S. dubia Christiansen et Bellinger, 1980, Lepidocyrtus finus Christiansen et Bellinger, 1980 en trampas Malaise (Palacios-Vargas y Gómez-Anaya 1993), mientras que Megalothorax minimus (Neelidae) tiene poco movimiento vertical.
El estudio muestra la estrecha funcionalidad de las dos cuencas con mayor similitud en producción de hojarasca en Chamela y puede ser utilizado como patrón conservado en estudios de contraste con localidades con diferente tiempo y uso de suelo.
Agradecimientos
Los autores agradecen a la Dirección General de Asuntos del Personal Académico, UNAM, su apoyo al proyecto: “Efectos del Uso de una Selva Baja Caducifolia sobre la Fauna de Artrópodos” (IN2078/91) y a la M.Sc. Alicia Rodríguez Palafox, por su gran ayuda y compañerismo durante el trabajo de campo.
Literatura citada
ARBEA, J. I.; JORDANA, R. 1985. Estudio ecológico de la colembofauna de los suelos del Macizo de Quinto Real (Pinineos Occidentales) y descripción de dos especies nuevas: Anurida flagellata sp. n. y Onychiurus subedinensis sp. n. (Insecta, Collembola). Boletín de la Estación Central de Ecología (Madrid) 74 (28): 57-80. [ Links ]
BARETTA, D.; SEABRA F., C.; SOUSA, J. P.; NOGUEIRA C., E. J. B. 2008. Colêmbolos (Hexapoda: Collembola) como bioindicadores de qualidade do solo em áreas com Araucaria agustifolia. Revista Brasileira de Ciência do Solo 32 (n. spec.): 2693-2699. [ Links ]
BENGTSSON, G.; GUNNARSSON, T.; RUNDGREN, S. 1983. Growth changes caused by metal uptake in a population of Onychiurus armatus (Collembola) feeding on metal polluted fungi. Oikos 40 (2): 216-225. [ Links ]
BENGTSSON, G.; GUNNARSSON, T.; RUNDGREN, S. 1985a. Influence of metals on reproduction, mortality and population growth in Onychiurus armatus (Collembola). Journal of Applied Ecology 22: 967-978. [ Links ]
BENGTSSON, G.; OHLSSON, L.; RUNDGREN, S. 1985b. Influence of fungi on growth and survival of Onychiurus armatus (Collembola) in a metal polluted soil. Oecología 68 (1): 63-68. [ Links ]
BIRD, S.; COULSON, R. N.; CROSSLEY, D.A. JR. 2000. Impacts of silvicultural practices on soil and litter arthropods diversity in a Texas pine plantation. Forest Ecology and Management 131 (1-3): 65-80. [ Links ]
CASSAGNE, N.; GERS, CH.; GAUQUELIN, T. 2003. Relationships between Collembola, soil chemistry and humus type in forest stands (France). Biology and Fertility of Soils 37 (6): 355- 361. [ Links ]
CHAGNON, M..; HÉBERT, C.; PARÉ, D. 2000a. Community structure of Collembola in sugar maple forests: relation to humus type and seasonal trends. Pedobiologia 44 (2): 148-174. [ Links ]
CHAGNON, M..; PARÉ, D.; HÉBERT, C. 2000b. Relationships between soil chemistry, microbial biomass and the collembolan fauna of southern Québec sugar maple stands. Ecoscience 7 (3): 307-316. [ Links ]
CHAGNON, M.; PARÉ, D.; HÉBERT, C.; CAMIRE, C. 2001. Effects of experimental living on collembolan communities and soil microbial biomass in a southern Québec sugar maple (Acer saccharum Marsh.) stand. Applied Soil Ecology 17 (1): 81-90. [ Links ]
CHRISTIANSEN, K.; P. BELLINGER 1980-81. The Collembola of North America North of the Rio Grande. A taxonomic analysis. Grinnell College, Iowa. 1322 p. [ Links ]
CUTZ-POOL, L. Q.; PALACIOS-VARGAS, J. G.; CASTAÑO MENESES, G.; GARCÍA-CALDERÓN, N. E. 2007. Edaphic Collembola from two agroecosystems with contrasting irrigation type in Hidalgo State, México. Applied Soil Ecology 36 (1): 46-52. [ Links ]
DÍAZ, A. ; J. NAJT 1990. Étude des peuplements de microarthropodes dans deux Páramos de l´État de Mérida (Venezuela). II. Structure des peuplements. Revue d´Écologie et de Biologie du Sol 27 (2): 331-340. [ Links ]
DUNGER, W. 1986. Observations on the ecological behaviour of some species of the Tullbergia krausbaueri- group. p. 111-115. En: Dallai, R. (ed.). 2nd International Seminar on Apterygota. Siena. [ Links ]
FABER, J. 1992. Soil fauna stratification and decomposition of Pine litter. Ph.D. thesis, Free University Amsterdam, Amsterdam. 131 p. [ Links ]
FRAMPTON, G. K. 1994. Sampling to detect effects of pesticides on epigeal Collembola (springtails). Aspects in Applied Biology 37: 121-130. [ Links ]
FRAMPTON, G. K. 1997. The potential of Collembola as indicators of pesticide usage: evidence and methods from de UK arable ecosystem. Pedobiologia 41 (1-3): 179-184. [ Links ]
GAMMA, M. M. Da; NOGUEIRA, A.; MÚRIAS, A. F. A. 1991. Effets du reboisement par Eucalyptus globulus sur les Collemboles édaphiques. Revue d´Écologie et de Biologie du Sol 28 (1): 9-18. [ Links ]
GARCÍA-OLIVA, F. ; EZCURRA, E.; GALICIA, L. 1991. Pattern of rainfall decomposition in the Central Pacific coast of México . Geografiska Annaler, Series A, Physical Geography 73 (3-4): 179-186. [ Links ]
GREENSLADE, P. 1997. Are Collembola useful as indicators of the conservation value of native grasslands? Pedobiologia 41 (1-3): 215-220. [ Links ]
HAGVAR, S. 1982. Collembola in Norwegian coniferous forest soils I. Relations to plant communities and soil fertility. Pedobiologia 24 (5-6): 255-296. [ Links ]
HAGVAR, S. 1983. Collembola in Norwegian coniferous forest soils II. Vertical distribution. Pedobiologia 25 (6): 383-401. [ Links ]
HAGVAR, S.; ABRAHAMSEN, G. 1984. Collembola in Norwegian coniferous forest soils. III. Relations to soil chemistry. Pedobiologia 27 (5): 331-339. [ Links ]
HENEGHAN, L.; BOLGER, T. 1998. Soil microarthropod contribution to forest ecosystem processes: the importance of observational scale. Plant and Soil 205 (2): 113-124. [ Links ]
HOLT, J. A. 1985. Acari and Collembola in the litter and soil of three north Queensland rain forest. Austral Ecology 10 (1): 57-65. [ Links ]
HUHTA, V., KARPPINEN. E., NURMINEN, M.; A. VALPAS 1967. Effect of silvicultural practices upon arthropods, annelid and nematode populations in coniferous forest soil. Annales Zoologici Fennici 4: 87-135. [ Links ]
IZARRA DE, D. C.; BOO, R. 1980. Los efectos de una reforestación con plantas introducidas sobre los microartrópodos del suelo. Ecología (Argentina) 5: 59-70. [ Links ]
JACKSON, M. 1982. Análisis Químicos de Suelos. 4ª. Omega, Madrid. 662 p. [ Links ]
JANZEN, D. 1988. Tropical dry forest: the most endangered mayor tropical ecosystem. p. 130-137. En: Wilson, E. O. (ed.). Biodiversity. National Academy Press, Washington, D. C. USA. 521 p. [ Links ]
JOHNSTON, J. M. 2000. The contribution of microarthropods to aboveground food webs: a review and model of belowground transfer in a coniferous forest. The American Midland Naturalist 143 (1): 226-238. [ Links ]
JORDANA, R.; ARBEA, J. I.; MORAZA, L.; MONTENEGRO, E.; MATEO, M. D.; HERNÁNDEZ, M. A.; HERRERA, L. 1987. Effect of reforestation by conifers in natural biotopes of middle and South Navarra (Northern Spain). Revue Suisse de Zoologie 94 (3): 491-502. [ Links ]
KOVÁC, L.; MIKLISOVÁ, D. 1997. Collembola communities (Hexapoda, Collembola) in arable soils of East Slovakia. Pedobiologia 41 (1-3): 62-68. [ Links ]
LAVÈLLE, P.; MAURY, E.; SERRANO, V. 1981. Estudio cuantitativo de la fauna del suelo en la región de Laguna Verde, Veracruz. Época de lluvias, pp. 65-100. En: Reyes-Castillo, P. (Ed.). Estudios Ecológicos en el Trópico Mexicano. Instituto de Ecología A.C., Xalapa. México. 105 p. [ Links ]
LORANGER, G.; BANDYOPADHYAYA, I.; RAZAKA, B.; PONGE, J.-F. 2001. Does soil acidity explain altitudinal sequences in collembolan communities? Soil Biology and Biochemistry 33 (3): 381-393. [ Links ]
LOTT, E.J.; BULLOCK, S. H.; SOLÍS-MAGALLANES, J. A. 1987. Floristic diversity and structure of upland and arroyo forests of Coastal Jalisco. Biotropica 19 (3): 228-235. [ Links ]
MAASS, J. M.; JARAMILLO, V.; MARTÍNEZ-YRIZAR, A.; GARCÍA-OLIVA, F.; PÉREZ-JIMÉNEZ, A.; SARUKHÁN, J. 2002. Aspectos funcionales del ecosistema de Selva Baja Caducifolia en Chamela, Jalisco, México. p. 525-542. En: Noguera, F. A; Vega, J. H; García-Aldrete, A. N.; Quesada, M. (eds.). Historia Natural de Chamela. Instituto de Biología, UNAM, México. 568 p. [ Links ]
MCCUNE, B.; GRACE, J. 2002. Analysis of ecological communities. MjM Software Design, Gleneden Beach, Oregon. USA. 300 p. [ Links ]
MENDOZA A., M. S. 1995. Los insectos colémbolos y la sucesión secundaria del bosque mesófilo de la reserva de la biósfera"El Cielo", Tamaulipas. Tesis de Licenciatura. ENEP-Iztacala, UNAM. México. 92 p. [ Links ]
MENDOZA, S.; VILLALOBOS, F. J.; RUIZ, L.; CASTRO A. E. 1999. Patrones ecológicos de los colémbolos en el cultivo de maíz en Balún Canal, Chiapas, México. Acta Zoológica Mexicana (n.s.) 78: 83-101. [ Links ]
MERILÄ, P.; OHTONEN, R. 1997. Soil microbial activity in the coastal Norway spruce [Picea abies (L.) Karst.] forest of the Gulf of Bothnia in relation to humus-layer quality, moisture and soil types. Biology and Fertility of Soils 25 (4): 361-365. [ Links ]
MIRANDA, R. A.; PALACIOS-VARGAS, J. G. 1992. Estudio comparativo de las comunidades de colémbolos edáficos de bosque de Abies religiosa y cultivo de haba (Vicia faba). Agrociencia serie Protección Vegetal 3 (4): 7-18. [ Links ]
NAJT, J. 1973. Algunos conceptos sobre la biología de los suelos como ciencia de nuestro tiempo. IDIA Suplemento Forestal 29: 97-105. [ Links ]
NOGUERA, F. A.; VEGA-RIVERA, J. H.; GARCÍA-ALDRETE, A. N.; QUESADA-AVENDAÑO, M. 2002. Historia Natural de Chamela. Instituto de Biología, UNAM, México, D.F. 568 p. [ Links ]
OLEJNICZAK, I. 2000. Communities of soil microarthropods with special reference to Collembola in midfield shelterbelts. Polish Journal of Ecology 52 (2): 123-133. [ Links ]
PALACIOS-VARGAS, J. G. 1985. Los Microartrópodos del Popocatépetl. Aspectos ecológicos y biogeográficos de los Oribátidos (Acarida) y los colémbolos (Apterygota). Tesis doctoral. Fac. Ciencias UNAM. 114 p. [ Links ]
PALACIOS-VARGAS, J. G. 1997. Catálogo de los Collembola de México. Coordinación de servicios editoriales, Facultad de Ciencias, UNAM, México, D. F. 102 p. + 20 fotos. [ Links ]
PALACIOS-VARGAS, J. G. 2000. Los Collembola (Hexapoda: Entognatha) de Jalisco, México. Dugesiana 7 (1): 23-36. [ Links ]
PALACIOS-VARAGAS, J. G.; GÓMEZ-ANAYA, J. A. 1993. Los Collembola (Hexapoda: Apterygota) de Chamela, Jalisco, México (Distribución Ecológica y Claves). Folia Entomológica Mexicana 89: 1-34. [ Links ]
PALACIOS-VARGAS, J.G.; CASTAÑO-MENESES, G.; GÓMEZANAYA, J. A. 1998. Collembola from the canopy of a Mexican tropical deciduous forest. Pan Pacific Entomologist 74 (1): 47- 54. [ Links ]
PALACIOS-VARGAS, J. G.; CASTAÑO-MENESES, G.; GÓ MEZ-ANAYA, J. A.; MARTÍNEZ-YRIZAR, A.; MEJÍA-RECAMIER, B. E.; MARTÍNEZ-SÁNCHEZ, J. 2007. Litter and soil arthropods diversity and density in a tropical dry forest ecosystem in Western México . Biodiversity and conservation 16 (13): 3703-3717. [ Links ]
PAQUIN, P.; CODERRE, D. 1997. Changes in soil macroarthropod communities in relation to forest maturation through three successional stages in the Canadian boreal forest. Oecología 112 (1): 104-111. [ Links ]
PINTO, C.; SOUSA, J. P.; GRAÇA, M. A. S.; GAMA DA, M.M. 1997. Forest Soil Collembola. Do tree introduction make a difference? Pedobiologia 41 (1-3): 131-138. [ Links ]
PONGE, J. F.; VANNIER, G.; ARPIN, P.; DAVID, J. F. 1986. Caractérisation des humus et des matières par la faune du sol. Intérêt sylvicole. Revue Forestiére Française, 38 (6):509-516. [ Links ]
PRAT, B.; MASSOUD, Z. 1980. Étude de la communauté des Collemboles dans un sol forestier. I. Structure du peuplement. Revue d´Écologie et de Biologie du Sol 17 (2): 199-216. [ Links ]
REYES, G. G.; BRECEDA, A. M. 1985. Análisis de la composición florística y estructura de la vegetación secundaria derivada de un bosque mesófilo de montaña en Gómez Farías, Tamaulipas (México). Tesis de licenciatura, Facultad de Ciencias, UNAM, México, 168 p. [ Links ]
SALAMON, J. A.; SCHAEFER, M.; ALPHEI, J.; SCHMID B.; SCHEU, S. 2004. Effects of plant diversity on Collembola in an experimental grassland ecosystem. Oikos 106 (1): 51-60. [ Links ]
SCHAEFER, M.; SCHAUERMANN, J. 1990. The soil fauna of beech forests: comparison between a mull and a moder soil. Pedobiologia 34 (5): 299-314. [ Links ]
STATSOFT. 2006. STATISTICA data analysis software system and computer manual. Versión 7.1. StatSoft Inc., Tulsa. [ Links ]
TAKEDA, H. 1981. Effects of shifting cultivation on the soil mesofauna with special reference to collembolan population in the north-east Thailand . Memoirs of the College of Agriculture , Kyoto University 118:45-60. [ Links ]
TANAKA, M., Y. SUGI, S. TANAKA, Y. MISHIMA ; R. HAMADA, 1978. Biological Production in a warm-temperate evergreen oak forest of Japan. JIBP Synthesis, Japanese Committee for the International Biological Program. 18: 147-165. [ Links ]
TER BRAAK , C. J. F.; SMILAUER, P. 1998 CANOCO Reference Manual and User´s guide to Canoco for Windows: Software for canonical Community Ordination, versión 4. Ithaca, New York, USA, 352 p. [ Links ]
TREJO, I.; DIRZO, R. 2000. Deforestation of seasonally dry tropical forest: a national and local analysis in México. Biological Conservation 94 (2): 133 -142. [ Links ]
VEGTER, J. J.; JOOSSE, E. N. G.; ERNSTING, G. 1988. Community structure, distribution and population dynamics of Entomobryidae (Collembola). Journal of Animal Ecology 57 (3): 971-981. [ Links ]
VILLALOBOS, F. J. 1989. Los colémbolos Poduromorpha (Apterygota: Insecta) y la sucesión secundaria del Bosque Mesófilo de Montaña. Biotam, Tamaulipas, México 1: 45-52. [ Links ]
VILLALOBOS, F. J. 1990. Estudio preliminar sobre la abundancia y diversidad de los Collembola (Apterygota) de un bosque tropical del Noreste de México. Folia Entomológica Mexicana 80: 5-29. [ Links ]
WAIKHOM, M.D., SINGH B.; JOYMATI D.L. 2006. Vertical distribution pattern of Collembola in a sub tropical forest floor of Manipur. Zoos´Print Journal 21 (7): 2331-2332. [ Links ]

