










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488On-line version ISSN 2665-4385
Rev. Colomb. Entomol. vol.36 no.1 Bogotá Jan./June 2010
Parandrinae (Coleoptera: Cerambycidae) de Colombia
Parandrinae (Coleoptera: Cerambycidae) from Colombia
JULIANA CARDONA-DUQUE1, ANTONIO SANTOS-SILVA2 y MARTA WOLFF3
1 Autor para correspondencia: Bióloga, Bs.C. Departamento de Biología, Call Box 9000, Universdad de Puerto Rico, Mayagüez, PR 00681, USA; Grupo de Entomología Universidad de Antioquia (GEUA), jcardonad@gmail.com.
2 Biólogo, B.Sc. Museu de Zoologia, Universidade de São Paulo, Caixa Postal 42594, 04299-970 São Paulo, SP, Brasil, toncriss@uol.com.br
3 Bióloga, Ph.D. Directora Grupo de Entomología Universidad de Antioquia (GEUA). Profesora, Instituto de Biología. Universidad de Antioquia. Calle 67 No. 53-108. Bloque 7-311. Medellín-Colombia, mwolff@matematicas.udea.edu.co.
Recibido: 18-jul-2009 - Aceptado: 2-abr-2010
Resumen: Parandrinae es una subfamilia de Cerambycidae con 63 especies descritas, de las cuales 45 están presentes en América. Se examinaron 266 especímenes recolectados en Colombia, depositados en 17 colecciones colombianas y otros museos en Brasil. Se registran 12 especies, diez de estas a partir de los especímenes examinados y dos a partir de los registros de literatura. Se excluyen Acutandra degeeri (Thomson, 1867), Acutandra punctatissima (Thomson, 1861) y Parandra ubirajarai (Santos-Silva, 2001) de la fauna colombiana. Se corrigen las instituciones depositarias de algunos tipos y se incluyen características que permiten identificar la hembra de Acutandra ubitiara (Santos-Silva & Martins, 2000). Se presentan claves de identificación para los géneros, subgéneros y especies de Parandrinae presentes en Colombia. Se incluyen mapas de distribución.
Palabras clave: Acutandra. Birandra. Escarabajos longicornios. Hesperandra. Parandra.
Abstract: The Parandrinae are a Cerambycidae subfamily with 63 described species, from which 45 occur in America. A total of 266 specimens collected in Colombia was examined, deposited in 17 Colombian collections and other museums in Brazil. Twelve species are reported, ten of them from the specimens examined and two from reports in the literature. Acutandra degeeri (Thomson, 1867), Acutandra punctatissima (Thomson, 1861) and Parandra ubirajarai (Santos-Silva, 2001) are excluded from the Colombian fauna. Depository institutions for some type specimens are corrected and characters that permit identifying the female of Acutandra ubitiara (Santos-Silva & Martins, 2000) are included. Identificatión keys are presented for the genera, subgenera and species of the Parandrinae present in Colombia. Distributión maps are included.
Keywords: Acutandra. Birandra. Longhorned beetles. Hesperandra. Parandra.
Introducción
Los coleópteros son el grupo de organismos más especioso y ampliamente distribuido en el planeta, representan cerca del 25% de todos los organismos. Una de las familias más diversas es Cerambycidae con aproximadamente 9.000 especies descritas para la región Neotropical y cerca de 750 registradas para Colombia (Linsley 1961; Martínez 2000). Junto con Chrysomelidae conforman la superfamilia Chrysomeloidea, que sumada a Curculionoidea conforma el clado Phytophaga (Farrell y Sequeira 2004; Marvaldi et al . 2009).
Parandrinae es un grupo morfológicamente atípico dentro de Cerambycidae. Presentan antenas cortas y segmentos tarsales bien definidos, por lo cual son considerados un grupo inusual dentro de Chrysomeloidea (Lawrence y Britton 1994). Este grupo tiene 63 especies descritas, 45 registradas para el nuevo mundo: 44 en la tribu Parandrini Blanchard, 1845 y una en la tribu monotípica Erichsoniini Thomson, 1861. Para Colombia la información proviene de algunos listados de especies (e.g. Blackwelder 1946; Monné y Giesbert 1994; Martínez 2000; Monné y Hovore 2006; Monné 2006), de las descripciones originales y de algunas revisiones de los grupos (e.g. Arigony 1978; Santos-Silva 2002, 2003a), sin que exista una revisión reciente del grupo en el país. Este trabajo tiene como objetivo actualizar el conocimiento taxonómico de los Parandrinae presentes en Colombia.
Posición Sistemática. Latreille (1802) al describir Parandra (cf. Santos-Silva y Shute 2009) lo colocó en “Trogossitiers” (Trogossitidae, Cleroidea) y allí fue mantenido por Olivier (1803). Posteriormente Latreille (1804) lo situó en Cucujidae, en Cerambycidae (1807), nuevamente en Cucujidae (1810) y finalmente los incluyó en “Platysomes” (1817). Así, la posición sistemática del género variaba e incluso fue considerado un género de “longicornios aberrantes” (e.g. Bates 1879). A partir de Schjödte (1865) la mayoría de los autores pasaron a considerar Parandra como un Prioninae dentro de Cerambycidae (e.g. Bates 1869, 1872, 1879; Gemminger y Harold 1872; Girard 1873; Lansberg 1884; Lameere 1885a, 1885b; Kolbe 1893; Gahan 1895). Sin embargo, algunos citaron Parandra en Cerambycidae sin especificar la relación con Prioninae (e.g. Horn 1875; Sharp 1878; Fairmaire 1881; Kirsch 1889; Blackburn 1895).
Lameere (1901) transfirió los géneros Hystatus Thomson, 1861 y Erichsonia Westwood, 1849 para “Parandrites” e incluyó el grupo en los “Prionides”. Consideró el género Parandra como el ancestro de los Cerambycidae y lo relacionó con los Trogossitidae (Lameere 1902). Posteriormente (1912, 1913, 1919) confirmó la posición del género en Prioninae, destacando la semejanza entre Parandra y el género Passandra Dalman, 1817 de Cucujidae. Aunque Gahan (1906) elevó el grupo a subfamilia, Craighead (1915) y Böving y Craighead (1931) mantuvieron Parandra en Prioninae. Finalmente Linsley (1961) consideró Parandrinae como subfamilia de Cerambycidae.
Por otro lado, Crowson (1955, 1960) se opuso a las teorías de Lameere sobre Parandra como la forma ancestral de Cerambycidae y su relación con los Trogossitidae. Argumentó que muchos caracteres de la larva y el imago en Parandra eran “avanzados” con relación a otros Cerambycidae y que probablemente la similitud con los trogosítidos era el resultado de evolución convergente debido a sus hábitos similares, pues viven como adultos bajo la corteza de árboles en descomposición. Sin embargo, resaltó la similitud con Cucujidae en características de las antenas, coxas, tarsos y la distribución geográfica. Para Crowson (1955) la posición sistemática de Parandra era dudosa y aunque había afirmado previamente que parecía ser un verdadero “Prionid”, también sostuvo que estaría bien incluido en Anoplodermatinae dadas sus afinidades con Migdolus (Crowson 1955). Sugirió entonces que Parandra es un Anoploderminae (= Anoplodermatinae) y que ese grupo se incluiría entre los Prioninae. Posteriormente propuso que Parandrinae, Prioninae y Anoplodermatinae forman un grupo natural y que Erichsonia y Parandra vienen de un ancestro “anoplodermatino” (Crowson 1960).
Parandrinae ha sido considerado un grupo de longicornios arcaicos (e.g. Saalas 1936; Linsley 1961, 1962; Chemsak 1996); incluso Linsley (1961) al proponer relaciones dentro de Cerambycidae, describió la morfología del posible ancestro del grupo, coincidente con la que describió para Parandrinae y Prioninae. Así mismo, se cree que el ancestro de los Cerambycidae vivió al interior de ramas, posiblemente alimentándose de madera en descomposición (Crowson 1960; Linsley 1961; Švácha et al . 1997), lo que coincide con la biología de los parandrinos y muchos prioninos.
Otros trabajos en sistemática han relacionado a Parandrinae con Prioninae, siendo Parandrinae el grupo más basal (Nakamura 1981). Igualmente el tipo de cordón nervioso y el canal alimentarío de las larvas de Parandra, son los menos especializados de Cerambycidae, siendo más afines a Prioninae que a Anoplodermatinae (Penteado-Dias 1984; Fonseca- Gessner 1990).
Napp (1994) realizó un análisis cladístico de Cerambycidae con caracteres morfológicos de larvas y adultos, y propuso una clasificación en la que reconoce a Parandrinae como subfamilia. Concluyó que Parandrinae, Anoplodermatinae y Prioninae conforman un grupo monofilético basal al resto de los Cerambycidae, soportado por: reducción de la galea, labro córneo y poco desarrollado, ausencia de lámina lateral del metendosternito y reducción de la vena ‘Sector´. Sin embargo, la restricción en el número de especies incluidas en su análisis (sólo una especie de Parandrinae, una de Anoplodermatinae y dos de Prioninae), la condujo a errores como establecer que la antena es glabra en Parandra, Anoploderma y Prioninae, y determinar que el desarrollo de puntuación y pilosidad en la antena era un caracter derivado del clado de cucújidos, crisomélidos y cerambícidos.
Más recientemente Švácha et al . (1997) y Farrell y Sequeira (2004) reconocieron a los Parandrinae como grupo hermano de Prioninae, incluidos en Cerambycidae. Además Švácha et al . (1997) excluyeron a los Anoplodermatinae, incluyéndolos en la familia Vesperidae. No obstante, la presencia de margen lateral en el pronoto, la ausencia de placa estridulatoria en el mesonoto y la morfología tarsal, agruparían a Parandrinae, Prioninae y Anoplodermatinae. Adicionalmente Švácha et al . (1997) anotan que los Parandrinae son un grupo monofilético soportado por caracteres como los tarsos simples claramente pentámeros, y el pronoto y algunas regiones proesternales en la larva cubiertas con asperitios esclerotizados o granos.
Además, Švácha et al . (1997) proponen que Parandrinae y Prioninae deberían ser agrupados en Prioninae pues no existirían características únicas para Prioninae, lo que resultaría en la parafilia de este grupo. Sin embargo, Švácha (1987) y Švácha et al . (1997) atribuyeron a Parandrini el estatus de tribu basándose exclusivamente en las larvas. Consideramos que proponer el estatus de grupo sólo con base en los caracteres larvales y sin considerar los caracteres imaginales puede ser problemático.
Caracteres como la unión de la vena empusal al ápice de la 1ra vena anal, sin unirse a la plical (Napp 1994) y la presencia de una segunda celda anal, son caracteres únicos a Prioninae. La venación alar en Parandrinae es más simplificada por la ausencia de la celda cerrada en el área anal (Saalas 1936; Linsley 1961; Arigony 1984; Napp 1994; Santos-Silva 2001b); incluso a veces puede presentarse una simplificación en las venas de esa área como en Neandra brunnea (Foster, 1771), o existir una celda vestigial como en algunos especímenes de Erichsonia dentifrons Westwood, 1849.
Adicionalmente, uno de los caracteres principales que distingue a los Parandrinae de Prioninae y las demás subfamilias de Cerambycidae, es la forma de los órganos genitales de las hembras. En el ovipositor de Parandra el paraprocto posee su esternito y terguito separados y aunque son inusualmente rígidos, abrazan la vagina y sus placas con sus partes anteriores (Saito 1990). Los estilos son muy pequeños, de conformación única y están articulados a la parte dorsal de los lóbulos de los coxitos (Saito 1990). Adicionalmente el cordón nervioso de los Parandrinae es único entre los Cerambycidae (Dias 1981; Penteado-Dias 1984). Concordamos con Saito (1990) quien reconoció a Parandrinae como subfamilia en contraste con la clasificación propuesta por Švácha (1987: 14).
Por otro lado no hay registro fósil de Parandrinae. Aunque Crowson (1955: 87, 145) y Linsley (1961: 60) mencionaron que Parandrexis parvula Martynov, 1926 era fósil de Parandrinae, aparentemente fue un error de estos autores al interpretar a Martynov (1926), quien afirmó que Parandrexis “no posee descendientes actuales” (traducido de Martinov 1926). Vitali (2006) anotó que Parandrexis está actualmente colocado en Parandrexidae Kirejtshuk, 1994 y transfirió Protospondylis florissantensis (Wickham, 1920) para Parandra (Parandra). Posteriormente Protospondylis Linsley, 1942 fue revalidado y puesto como incertae sedis en Cerambycidae por Santos-Silva y Shute (2009), ya que no es posible determinar el género usando las claves disponibles (Vitali 2006).
Clasificación actual. El género Parandra Latreille, 1802 fue dividido por Lameere (1912) en los subgéneros Archandra, Stenandra, Neandra y Parandra s. str. Posteriormente Quentin y Villiers (1972) elevaron Stenandra a estatus genérico, Arigony (1977) dividió Archandra en los subgéneros Archandra y Hesperandra y este último fue elevado por Villiers (1980) a género. Más adelante Santos-Silva (2002) elevó Archandra y Neandra a género, dividió Parandra en los subgéneros Parandra y Birandra y creó el género Acutandra. El mismo autor (2003a) dividió Hesperandra en los subgéneros Hesperandra, Tavandra y Zikandra, y recientemente, con la resolución de la identidad taxonómica de Parandra laevis Latreille, 1804, Santos-Silva y Shute (2009) realizaron algunos cambios nomenclaturales en la tribu. Actualmente, Parandrinae está compuesta por los géneros y subgéneros: Acutandra, Archandra, Birandra (Birandra), Birandra (Yvesandra), Neandra, Parandra (Hesperandra), Parandra (Parandra), Parandra (Tavandra) y Stenandra.
Características Biológicas y Distribución. Los Parandrinae son fitófagos polífagos, generalmente asociados a madera en descomposición (Linsley 1959). Habitan bajo la corteza de árboles muertos en zonas húmedas y grandes áreas boscosas (Arigony 1978; Triplehorn y Johnson 2005). Las larvas se alimentan exclusivamente de madera en descomposición y pueden encontrarse en el interior del bosque excavando galerías en troncos caídos semi-podridos (Lawrence 1991; Linsley 1961; Costa et al . 1988). Los adultos son nocturnos y con frecuencia son atraídos a la luz (Linsley 1961; White 1983; Tavakilian 2000). Las hembras ponen los huevos sobre troncos de árboles (White 1983) ovipositan sin la ayuda de las mandíbulas en la preparación del nicho, como sí lo hacen otros cerambícidos (Linsley 1959).
Entre los depredadores de Cerambycidae y otros coleópteros barrenadores se encuentran los Cucujidae, Trogossitidae, Cleridae, Colydiidae, Elateridae (larvas), entre otros coleópteros. Además hemípteros, neurópteros y dípteros como Asilidae, Xylophagidae y Rhagionidae. Algunos hymenópteros, nemátodos y hongos pueden parasitarlos. Otros depredadores pueden ser lagartijas, murciélagos y aves de las familias Picidae, Furnariidae, Caprimulgidae, Vireonidae, Parulidae, Sittidae y Tyrannidae (Linsley 1959, 1961).
Por otro lado, las especies de Parandrinae están presentes en los continentes y la mayoría de islas oceánicas, siendo más abundantes en la región Neotropical (Linsley 1961; Villiers 1980). Acutandra Santos-Silva se encuentra en Suramérica, Archandra Lameere se distribuye en la región Paleártica, Birandra Santos-Silva es cosmopolita, Erichsonia Westwood se encuentra en México y Centroamérica, Neandra Lameere en la región Neártica, Parandra Latreille en América y Stenandra Lameere en la región Etiópica.
Morfología de Parandrini. Aunque este trabajo se enfoca en los adultos, la morfología de las larvas y las pupas de Parandrinae ha sido descrita por varios investigadores (e.g. Linsley 1961; Costa et al . 1988; Lawrence 1991; Chemsak 1996; Lingafelter y Micheli 2004). Se conocen las larvas de Neandra brunnea (Fabricius, 1798), Birandra punctata (White, 1853), B. gabonica (Thomson, 1858), B. tavakiliani (Santos- Silva, 2002), Parandra glabra (DeGeer, 1774), P. expectata Lameere, 1902 y Archandra caspia (Ménétriés, 1832).
Los adultos de Parandrinae (Fig. 1A) tienen las antenas relativamente cortas (no se extienden más allá de la base del pronoto), se insertan en frente de los ojos cerca a la base de las mandíbulas (Fig. 1B); los antenómeros presentan áreas sensoriales, pilosidad y puntuación variable (Fig. 1C). El labro es triangular con ápice variable, fusionado al clípeo (Figs. 1D y 1E). Las mandíbulas son agudas, grandes y a menudo dentadas (Figs. 1D y 1E). Las cavidades coxales anteriores pueden ser abiertas o cerradas en la parte posterior (Figs. 1F y 1G). El mesonoto no tiene placa estridulatoria (Fig. 1H). Las patas son cortas y las coxas anteriores son fuertemente transversas con trocantín visible (Figs. 1F y 1G). Los tarsos son delgados, sin cojines ventrales y claramente pentámeros, con el tercer segmento entero o débilmente emarginado y generalmente presentan paroniquio tarsal (Fig. 1I y 1J). Las alas posteriores no tienen celdas cerradas en el sector anal, la celda radial es abierta y la vena 2A2 es ausente o vestigial (Forbes 1922).
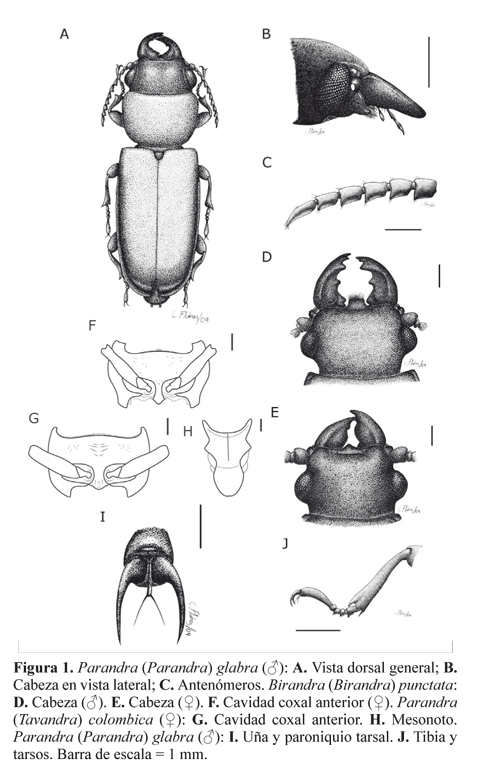
Aunque los Parandrinae no presentan dimorfismo sexual evidente como muchos cerambícidos, hay algunas características que pueden separar los machos de las hembras. En Birandra y Parandra las mandíbulas de las hembras normalmente son más triangulares y sin curvatura hacia la región apical (Fig. 1E); los machos pequeños, tienen mandíbulas falciformes más angostas en la base y un poco curvadas hacia el ápice (Fig. 1D). Además en Birandra el ápice del labro generalmente es truncado en los machos y agudo en las hembras (Figs. 1D y 1E). En los géneros presentes en América el último urosternito generalmente es similar en longitud al penúltimo en los machos y más largo en las hembras. Adicionalmente, el tamaño de la cabeza en relación al protórax normalmente es menor en la hembra, lo que sólo es evidente cuando se tiene una pareja.
Materiales y Métodos
Se revisaron especímenes en 17 colecciones colombianas, en el Museu Nacional de la Universidade Federal do Rio de Janeiro y en el Museu de Zoologia de la Universidade de São Paulo. Además se tomaron los registros de literatura para Colombia en los cuales la identidad taxonómica de las especies no es dudosa.
La distribución de las especies se basa en material examinado. La información de geoposición de las localidades para los especímenes que carecían de ésta, provino del Diccionario Geográfico de Colombia (IGAC 1996) y del Atlas de Colombia (IGAC 2002). En los casos en que la localidad de colección era incierta, se registraron las coordenadas de la cabecera municipal. Los mapas se hicieron usando el programa ArcGis v. 9.3 (ESRI® 1999-2008). Las referencias citadas bajo cada taxón se refieren a las citaciones originales, al catálogo de Monné (2006) y a citaciones posteriores u omisiones del catálogo.
Los siguientes son los acrónimos de las instituciones visitadas: CEUA, Colección de Entomología de la Universidad de Antioquia, Medellín; CFPL, Colección personal de Luís Carlos Pardo Locarno, Palmira; CIBM, Colección de la Corporación de Investigaciones Biológicas, Medellín; CMUC, Centro de Museos de la Universidad de Caldas, Manizales; GMIC, Colección particular de Gilberto Mendoza, Bogotá; IAvH, Instituto Alexander von Humboldt, Villa de Leyva; ICN (algunos ejemplares citados aparecerán con el acrónimo de catálogo ICN-MHN-CO), Colección del Instituto de Ciencias Naturales de la Universidad Nacional de Colombia, Bogotá; LEUC, Colección de Laboratorío de Entomología, Universidad de Caldas, Manizales; MCSJ, Museo del Colegio de San José de La Salle, Medellín; MEPB, Museo Entomológico de Piedras Blancas, Comfenalco, Medellín; MEUV, Museo de Entomología de la Universidad del Valle, Cali; MFLG, Museo Francisco Luís Gallego de la Universidad Nacional de Colombia, Medellín; MHNC, Museo de Historia Natural - Universidad del Cauca, Popayán; MHNS, Museo de Historia Natural, Universidad La Salle, Bogotá; MNRJ, Museu Nacional, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Rio de Janeiro; MUAM, Museo Universidad de Antioquia, Medellín; MUJC, Museo de la Pontificia Universidad Javeriana, Santafé de Bogotá; MZSP, Museu de Zoologia, Universidade de São Paulo, São Paulo, São Paulo; UNAB, Museo Entomológico Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. Y los acrónimos de otras instituciones mencionadas en el texto son: BMNH, The Natural History Museum, Londres, Reino Unido; FIOC, Fundação Instituto Oswaldo Cruz, Rio de Janeiro, Rio de Janeiro, Brasil; ISNB, Institut Royal des Sciences Naturelles de Belgique, Bruselas, Bélgica; MNHN, Muséum National d´Histoire Naturelle, Paris, Francia; MVMA, Museum of Victoria, Melbourne, Australia; NHRS, Naturhistoriska Riksmuseet, Estocolmo, Suiza; USNM, National Museum of Natural History, Washington, D. C., EEUU; SMTD, Staatliches Museum fur Tierkunde, Dresden, Alemania; ZSMC, Zoologische Staatssammlung des Bayerischen Staates, München, Alemania.
Resultados y Discusión
Se examinaron 288 especímenes adultos de Parandrinae, 266 provenientes de Colombia. Se registran especies de Parandrinae para 91 municipios del país, distribuidos en 19 departamentos, entre 0 y 3000 m. Se encontraron un total de 10 especies provenientes de Colombia, se incluyen los registros de tres especies más y se excluyen Acutandra degeeri (Thomson, 1867), Acutandra punctatissima (Thomson, 1861) y Parandra ubirajarai (Santos-Silva, 2001) de la fauna de Colombia.
PARANDRINAE Blanchard, 1845
Parandrites Blanchard, 1845: 134.
Parandrinae; LeConte y Horn, 1883: 265; Monné, 2006: 8.
PARANDRINI Blanchard, 1845
Parandrites Blanchard, 1845: 134.
Parandrini; Monné, 2006: 8.
Clave para los géneros y subgéneros de Parandrini presentes en Colombia
1. Cavidades procoxales cerradas en su parte posterior (Fig.1G) (Parandra Latreille, 1802).............................................2 - Cavidades procoxales abiertas en su parte posterior (Fig. 1F).........................................................................................3 2(1). Proyección del borde ocular posterior no sobresaliente (Figs. 1A y 3B). Submento no deprimido, glabro con pelos muy cortos y muy dispersos (si se presentan pelos largos, son muy pocos); puntuación generalmente fina. Carena del submento inconspicua o ancha y poco elevada ............................. .................................... Parandra (Parandra) Latreille, 1802 - Proyección del borde ocular posterior sobresaliente o muy sobresaliente (Figs. 4A, 4B, 5A y 6A). Submento deprimido o no, con pelos largos y abundantes o cortos y dispersos; puntuación gruesa. Carena del submento delgada y elevada, ausente o apenas indicada......……..…...Parandra (Tavandra) Santos-Silva, 2003
3(1). Mandíbulas poco desarrolladas, no falciformes, túmidas en la cara lateral-externa, y semejantes a las de las hembras en los machos de gran tamaño; ápice del labro agudo en los dos sexos (Figs. 1E y 7A); margen anterior del pronoto, en general, fuertemente sinuoso en los dos sexos………… .............................................................................Acutandra Santos-Silva, 2002 [Acutandra ubitiara (Santos-Silva & Martins, 2001)]
- Mandíbulas bien desarrolladas y falciformes en los machos de gran tamaño, con dimorfismo sexual y no túmidas en la cara lateral-externa; ápice del labro de los machos truncado (Fig. 1D); margen anterior del pronoto débilmente sinuoso o no sinuoso en los dos sexos (Birandra Santos-Silva, 2002).... ...............................................................................................4 4(3). Área sensorial del antenómero XI dividida por carena .......................................................Birandra (Yvesandra) Santos-Silva & Shute, 2009 [Birandra (Yvesandra) antioquensis (Cardona-Duque et al . 2007)]
- Área sensorial del antenómero XI no dividida por carena .................................Birandra (Birandra) Santos-Silva, 2002
Parandra Latreille, 1802
Parandra Latreille, 1802: 160; Olivier, 1803: 100.
Parandra (Hesperandra) Arigony, 1977: 163.
Hesperandra; Chalumeau & Touroult, 2005: 41; Monné, 2006: 9 (cat.); Santos-Silva & Shute, 2009: 30 (syn.).
Especie-tipo. Attelabus glaber DeGeer, 1774 (monotipia).
Cabeza grande; borde ocular posterior muy sobresaliente (Figs. 4A, 4B, 5A y 6A) o apenas proyectado (Figs. 1A, 2A, 3B y 3C). Mandíbulas de machos y hembras sin diente apical en la cara externa; en los machos de gran tamaño, falciformes y frecuentemente un poco más largas que la cabeza (Figs. 2A, 3B y 5D); en los machos pequeños y las hembras, levemente curvas en la cara externa y más cortas que la cabeza (Figs. 2B, 3C, 4B y 5A). Antenómero XI sin área sensorial dorsal. Área sensorial de los antenómeros III-XI dividida por una carena longitudinal, en general bien marcada en todos los antenómeros, pero algunas veces ausente en los antenómeros basales, apenas indicada en los antenómeros intermedios y sólo nítida en el antenómero XI. Cavidades coxales anteriores cerradas en la parte posterior (algunas veces, con una pequeña abertura). Paroníquio con setas.
Todas las especies de este género se distribuyen en América.
Parandra (Parandra) Latreille, 1802
Parandra Latreille, 1802: 160; Santos-Silva & Shute, 2009: 30.
Hesperandra (Zikandra) Santos-Silva, 2003a: 120; Chalumeau & Touroult, 2005: 42; Monné & Hovore, 2006: (cat.); Monné, 2006: 12 (cat.); Santos-Silva, 2007: 5 (clave); Santos- Silva & Shute, 2009: 30 (syn.).
Gnathophorus Kirby, 1837: 166; White, 1853: 2 (syn.); Gemminger & Harold, 1872: 2751 (cat.; en syn.).
Isocerus; Germar, 1846: 19 (parte).
Especie-tipo. De Parandra, Attelabus glaber DeGeer, 1774, monotipia; de Hesperandra (Zikandra) y de Gnathophorus, Attelabus glaber DeGeer, 1774 (respectivamente designación original y monotipia).
Proyección del borde ocular posterior no sobresaliente. Submento no deprimido, glabro o con pelos muy cortos y dispersos (aunque pueden presentarse algunos pelos largos); margen anterior poco o moderadamente elevado, precedido o no por un surco. Carena ventral de las genas inconspicua o ancha y poco elevada.
Clave para las especies de Parandra (Parandra) presentes en Colombia
1. Metepisternos con pilosidad conspícua en toda su extensión (Fig. 3G)...........................P. (Parandra) humboldti (Santos-Silva, 2003)
- Metepisternos glabros o con pilosidad restricta al tercio anterior (Fig. 2I)..........................…P. (Parandra) glabra (DeGeer, 1774)
Parandra (Parandra) glabra (DeGeer, 1774)
Attelabus glaber DeGeer, 1774: 352. Localidad-tipo: Indias (?). (NHRS).
Scarites testaceus Fabricius, 1794: 437. Localidad-tipo: Senegallia. (NHRS).
Parandra ferruginea Sturm, 1826: 181. Localidad-tipo: Brasil. (ZSMC).
Parandra lineola Gory in Guérin-Méneville, 1831: estampa 42, figs. 7-7a. Localidad-tipo: no indicada. (Depositarío desconocido).
Parandra mandibularis Perty, 1832: 84. Localidad tipo: Amazonum Flumen. (ZSMC).
Parandra maxillosa Laporte, 1840: 387. Localidad-tipo: Brasil. (MNHN).
Parandra lineolata Gory in Guérin-Méneville, 1844: 207 (error).
Parandra grandis Thomson, 1861b: 79. Localidad-tipo: Colombia. (MNHN).
Parandra barbata Thomson, 1861c: 95. Localidad-tipo: Colombia. (MNHN).
Parandra occipitalis Thomson, 1867: 108. Localidad-tipo: Colombia. (MNHN).
Parandra obsolescens Casey, 1912: 217. Localidad-tipo: Panamá, Chiriquí. (USNM).
Parandra (Archandra) lucida Zikán, 1948: 36. Localidadtipo: Brasil, Rio de Janeiro: Itatiaia. (FIOC).
Parandra (Archandra) glabra var. translucida Zikán, 1948: 37. Localidad-tipo: Brasil, Santa Catarina: Nova Teutonia. (FIOC).
Hesperandra (Zikandra) glabra; Monné, 2006: 13 (cat.). Parandra (Parandra) glabra; Santos-Silva & Shute, 2009: 31 (comb. nov.).
Diagnosis. Cabeza grande (un poco más corta que el pronoto en especímenes de tamaño promedio), sin puntuación aparente, un poco visible en la parte posterior de los ojos (Figs. 2A y 2B). Submento con puntuación generalmente muy fina y dispersa (algunos individuos pueden presentar una puntuación levemente gruesa e incluso un poco anastomosada) (Fig. 2C); margen anterior del submento elevado, precedido por surco que puede ser raso; labro no proyectado (no sobrepasando las esquinas anteriores de la cabeza). Mandíbulas de los machos con un diente pequeño un poco después del medio (los machos minor pueden tener las mandíbulas subtriangulares) (Fig. 2A); mandíbulas de las hembras subtriangulares (Fig. 2B). Antenómeros IV-VI más largos que anchos (Fig. 2D), divididos por carena en la cara ventral; antenómero X al menos un tercio más corto que el XI. Ojos de las hembras anchos en vista lateral (Fig. 2E). Protórax de los machos con marginación lateral completa (Fig. 2F); pronoto apenas puntuado (puntos imperceptibles) (Figs. 2G y 2H). Metepisternos glabros o con muy pocos pelos hacia el tercio anterior (Fig. 2I). Proesterno y metaesterno lisos, glabros (algunos individuos pueden presentar algunos pelos dispersos) (Fig.2J). Élitros lisos.

Distribución. Desde México, Centroamérica (incluyendo parte de las Antillas) y Suramérica (Fig. 10A).
Comentarios. Parandra glabra es una especie con una gran variación en tamaño y coloración, y aunque no se han encontrado patrones en esta variación que permitan separarla en especies diferentes, hacen falta estudios más detallados a lo largo de su ámbito (área de distribución) (Bates 1879; Arigony 1978; Villiers 1980; Tavakilian 2000). Esta variación relacionada a la distribución, podría corroborar que se trata de especies muy semejantes o variaciones poblacionales de la misma especie. Algunas de estas formas corresponden a especies descritas que hoy son sinónimos de P. glabra.
Dado que no existe un conjunto de características morfológicas externas que sean conservadas dentro de P. glabra, es necesarío explorar otros caracteres que permitan separarla de las demás especies del subgénero. La mayoría de los caracteres que definen a P. glabra son variables, y las diferentes combinaciones de esta variación son permisibles dentro del concepto amplio que hoy tenemos de esta especie, lo que hace imposible concluir si la especie es extremadamente variable o si se trata de un complejo de especies muy próximas entre sí.
En general la forma proveniente de Colombia y Ecuador corresponde a Parandra grandis Thomson, 1861 (fotografía del lectotipo examinada). Los machos de esta región se diferencian de los del sur y suroeste brasilero por el cuerpo más robusto, el tercio apical de las mandíbulas menos curvado, el borde ocular posterior más sobresaliente (esto se observa en vista dorsal, con relación al cuello), el área posterior a los ojos con puntuación muy gruesa y anastomosada, el submento con puntuación gruesa y abundante (a veces anastomosada), los ángulos posteriores del pronoto más redondeados, el margen del tercio latero-anterior del pronoto más visible en vista dorsal, y los metepisternos frecuentemente con puntos evidentes en toda su extensión (a veces acompañados de pelos cortos). Las hembras se diferencian de los machos por la puntuación más fina y dispersa en el submento; sin embargo, esta puntuación puede ser más gruesa y abundante que en las hembras del sur y sureste de Brasil; la puntuación y pilosidad de los metepisternos es como en los machos. Los machos del sur y sureste brasilero generalmente son más delgados, el tercio apical de las mandíbulas es más curvado, el borde ocular posterior no sobresale, el área posterior de los ojos presenta puntos finos o levemente gruesos y dispersos, la puntuación del submento es fina o muy fina y muy dispersa, los ángulos posteriores del pronoto son más angulados, el tercio lateroanterior del pronoto normalmente no es visible en vista dorsal, y los metepisternos poseen puntos finos y pelos cortos (apenas en el tercio o cuarto basal).
Por otro lado, Parandra barbata Thomson, 1861 y P. occipitalis Thomson, 1867 corresponden a la misma forma que difiere de P. grandis y se aproxima a los machos del sur y sureste brasilero, e incluso a los de Isla del Coco (Costa Rica). Se diferencian principalmente por el borde ocular posterior débilmente sobresaliente, el área posterior de los ojos con puntos más finos y dispersos (principalmente junto al ojo), las mandíbulas claramente más curvadas en el tercio apical y el tercio latero-anterior del pronoto no visible en vista dorsal. Sin embargo, el área posterior de los ojos de la forma de Isla del Coco es intermedia entre la de los especímenes del sur y sureste de Brasil y la de P. grandis: fina junto a los ojos y más rugosa y punteada en dirección al foramen. Igualmente, la puntuación del submento de Parandra barbata y P. occipitalis es muy similar a la encontrada en el centrooeste brasilero: más fina que en los especímenes de Isla del Coco y la forma P. grandis y más abundante que los del sur y sureste brasilero. Los especímenes del noreste de Suramérica (Brasil - estados de Amazonas y Pará - Guyana Francesa y Venezuela) coinciden con la forma de P. grandis al igual que los del noroeste (Colombia y Ecuador) pero tienen las mandíbulas claramente más falciformes (curvatura del tercio apical más acentuada) y la puntuación del submento más fina y abundante.
Parandra maxillosa sensu Thomson (1867) corresponde a la forma del sur y sureste brasileros. Examinamos la fotografía de uno de los especímenes estudiados por Thomson que porta una etiqueta de tipo, como lo afirmó Tavakilian (2000): “...ainsi que l´exemplaire etiquete «Parandra maxillosa Thomson, Type»”; sin embargo, ese especímen no es el tipo de P. maxillosa Laporte, 1840 puesto que el holotipo está depositado en MVMA (Tavakilian 2000). Examinamos la fotografía de ese holotipo y aparentemente corresponde a la forma de la Guyana Francesa, Venezuela y norte de Brasil (Amazonas y Pará). Igualmente examinamos la fotografía del holotipo de P. ferruginea Sturm, 1826 que correspondería a la misma forma.
Por otro lado, Perty (1832) en la descripción de P. mandibularis registró como localidad típica: “Habitat in sylvis ad Amazonum flumen”. Examinamos la fotografía del holotipo que está depositado en ZSMC, cuya etiqueta corresponde a aquella utilizada por Joseph Anton Maximilian Perty, presentada en Horn y Kahle (1937). Esta forma corresponde a la del sur y sureste brasileros y no a la de la Amazonía brasilera. Aunque la figura presentada por Perty (1832) se asemeja a los especímenes del norte de Brasil (Amazonas y Pará), es probable que el holotipo haya sido incorrectamente etiquetado en relación a la localidad de colecta.
Los especímenes del centro-oeste de Brasil tienen estados intermedios entre los del sur y sureste brasilero y aquellos de la Guyana Francesa, Venezuela y norte brasilero; igualmente entre los de Colombia y Ecuador y los del sur y sureste brasileros. Los pocos ejemplares estudiados de Paraguay y Argentina aparentemente corresponden a la misma forma del sur y sureste brasilero. Por otro lado, los machos de Costa Rica (excepto Isla del Coco) y Panamá, en general son muy semejantes a los de Colombia; sin embargo las hembras se asemejan más a las del sur y sureste brasileros, principalmente en la puntuación y pilosidad de los metepisternos. Esa forma corresponde a P. obsolescens Casey, 1912 (fotografía del holotipo examinada) y probablemente es igual a P. lineola Gory, 1831 proveniente de la isla de Guadalupe. Nuestra suposición se basa únicamente en la descripción de Gory (1844) [en Gory (1831) la especie fue sólo figurada], pues no examinamos especímenes procedentes de Guadalupe.
Por otro lado, Parandra lineola Gory en Guérin-Méneville, 1831 es un nombre válido pues hay una asociación de un nombre válido a una figura (ICZN 1999: artículo 12.2.7) y Parandra lineolata Gory en Guérin-Méneville, 1844 no tiene estatus nomenclatural pues es una ortografía subsecuente incorrecta (ICZN 1999: artículo 33.3) y no una enmienda para P. lineola. Cowan (1971) estudió las fechas de publicaión de la “Iconographie du règne animal de G. Cuvier...” de Félix Édouard Guérin-Méneville y concluyó que la parte con la plancha número 42 (con las figuras de P. lineola), fue publicada el 21 de mayo de 1831 y el texto completo el 7 de septiembre de 1844. Aunque la localidad típica de P. lineola no fue mencionada, se infiere que es Guadalupe (aquella de la descripcción de P. lineolata que no tiene localidad típica por ser un error de ortografía). La institución depositaria del(los) tipo(s) de P. lineola es desconocida (Tavakilian 2000; Monné 2006).
Thomson (1861b) al redescribir Parandra colombica White, 1853 escribió incorrectamente “Parandra columbica”. Sin embargo examinamos la fotografía de uno de los especímenes de la colección Thomson y conforme lo observó Lameere (1902), esa redescripción no corresponde a la especie de White (1853) sino a Parandra glabra. Tavakilian (2000) escribió al respecto: “L´exemplaire mâle étiqueté comme columbica par Thomson (MNHN) [il signale en posséder 12 exemplaires] est bien identique à l´espèce de Degeer,…”. La puntuación elitral <más fuerte> mencionada por Thomson (1861b), es mucho más fina que la de la verdadera Parandra colombica White, 1853. Curiosamente los especímenes incorrectamente interpretados como la especie de White por Thomson (1861b), han sido erróneamente considerados sintipos y la localidad como localidad típica (e.g. Monné 2006). Parandra colombica sensu Thomson (1861b) corresponde a la forma de P. grandis.
Por otro lado, Tavakilian (2000) registró en relación a Scarites testaceus Fabricius, 1794: “Au sujet de Scarites testaceus, d´après Zimsen (1964: 17), aucune espèce décrite por Fabricius de la collectión Gustav von Paykull (aujourd´hui au NRS), n´est étiqueté comme type. Sa patrie «Senegalia», doit être la conséquence d´une provenance mal interprétée”. Sin embargo, desconocemos a qué forma de P. glabra corresponde.
Examinamos fotografías del lectotipo hembra de Attelabus glaber DeGeer, 1774, concordando con Tavakilian (2000) quien dijo que el lectotipo es perfecrtamente idéntico a las hembras recolectadas en Guyana Francesa y que probablemente Surinam debe ser la verdadera localidad típica de Attelabus glaber y no “Indes” (traducido Indias Occidentales) como escribió DeGeer. La puntuación moderadamente gruesa y abundante en el submento a veces es mayor que en los especímenes procedentes de Colombia y corresponde a la forma encontrada en la Guyana Francesa, Venezuela y norte de Brasil (Amazonas y Pará). Así mismo, el macho presentado por DeGeer (1774) (espécimen extraviado) parece corresponder a la forma de esa región.
Sin duda P. glabra es la especie más ampliamente distribuida y comúnmente encontrada en las colecciones, conforme lo observado por Tavakilian (2000). En las colecciones colombianas encontramos 123 especímenes y ha sido recolectada en las tres cordilleras, los valles interandinos, las costas Atlántica y Pacífica, y la región Amazónica, desde el nivel del mar hasta los 2600 m. En los especímenes colombianos la puntuación de los élitros y el submento puede variar, entre especímenes con puntos relativamente gruesos y concentrados (principalmente en el submento) y otros sin puntuación aparente. Aunque en general las especies de Parandra poseen las cavidades coxales anteriores cerradas en la parte posterior, algunos individuos de P. glabra pueden presentarlas levemente abiertas.
Biología. Tavakilian (2000) y Monné (2002, 2004) listaron varias plantas hospederas para esta especie: Spondias mombin (Anacardiaceae), Aspidosperma sp. (Apocynaceae), Araucaria angustifolia, Araucaria brasiliana (Araucariaceae), Piptocarpha angustifolia (Asteraceae), Alnus acuminata subsp. acuminata (=Alnus jorulliensis) (Betulaceae), Ochroma pyramidale (Malvaceae), Cecropia sp. (Urticaceae), Phoebe sp. (Lauraceae), Acacia decurrens, Mimosa scabrella (Fabaceae), Morus alba (Moraceae), Eucalyptus globulus (Myrtaceae) y Salix humboldtiana (Salicaceae). De estas encontramos en Colombia: Spondias mombin (0-1000 m), Aspidosperma spp., Araucaria angustifolia y A. brasiliana (cultivadas), Alnus acuminata var. acuminata (1500-3000 m), Ochroma pyramidale (ampliamente cultivada, 0-1500 m), Cecropia spp., Phoebe spp. (distribuidas en el Neotrópico entre 800-2500 m), Acacia decurrens (1000-1500 m), Morus alba (1000-1500 m), Eucalyptus globulus y Salix humboldtiana (cultivadas).
Los adultos de P. glabra se encuentran debajo de corteza de sus hospederos a veces en pequeños grupos (cerca de una decena de individuos) (Villiers 1980). Han sido observados en la madera y corteza del balso, Ochroma pyramidale (Cav. ex Lam.) Urb., durante la noche (Villiers 1980). En los machos de esta especie, las antenas parecen tener una gran sensibilidad a alguna feromona de las hembras (Lingafelter 1998).
Material examinado. COLOMBIA. Amazonas. Río Tacana, 1 (H), en la selva, 30.X.1946, L. Richter (ICN-MHN-CO 2337, ICN); PNN Amacayacu, Caño Mata Mata, 100 m, 1 (H), II.1989, F. Fernández (IAvH 01286); PNN Amacayacu, Mata Mata, 3°23´01”S; 70°06´01”W, 150 m, 1 (H), Malaise, 2. III.2000-3.IV.2000, A. Parente (IAvH -3422). Antioquia. Santa Fe de Antioquia, 1 (H), manual en hoja, IX.1994, C. Echeverri (CEUA - 36716); Río Claro, 1 (M), manual en suelo, 3.I.1998, E. Muñoz, (MUA-IN 2931, MUA); Murrí, 1 (M), manual en bosque, I.1982, G. Galeano (MUA-IN 00836, MUA); Ciudad Bolívar, 1 (M), IV. 1953, F. L. Gallego (No. 8771, MFLG); Yolombó, 1 (M), en madera, V.1955, F. L. Gallego (No. 8771, MFLG); Turbo, 1 (M), en maleza, X.1947, F. L. Gallego (No. 8771, MFLG); Villarteaga, 1 (H), IV.1953, F. L. Gallego (No. 8771, MFLG); Medellín, 1 (H), en madera, IV.1959, F. L. Gallego (No. 8771, MFLG); Sopetrán, 1 (H), II.194?, F. L. Gallego (No. 8771, MFLG); Urrao, 1 (H), en pared, 27.VI.1989, J. Marío G. (No. 8771, MFLG); San Luís, 440 m, 1 (M), 3.III.1994, L-S (MUJ - 5923); Maceo, Vereda Santa Bárbara, Hacienda Santa Bárbara, 6°32´38,8”N; 74°38´24,8”W, 450 m, 1 (H), dentro de tronco, XII.2007, Campuzano, Cardona, Flórez y Sánchez (CEUA). Caldas. Manizales, 1 (H), en rosas, VIII.1948, F. L. Gallego (No. 8771, MFLG); Palestina, Vereda Santágueda, 1200 m, 1 (H), en pasto, 7.V.2006, Hoyos y Vásquez (LEUC); Manizales, Gallinazo (Río Blanco), 2100 m, 1 (M), bosque en árbol, 12.V.2006, Charry y Gonzáles (LEUC); Manizales, Km 41, 1 (H), en suelo, 28.VIII.1993, Cast. Osp. (CI-001-044-6, CMUC). Caquetá. San Vicente del Caguán, PNN Picachos, 2°47´51”N; 74°51´18”W, 1450 m, 1 (H), fuente de luz, alrededores casa escuela, XI.1997, F. Escobar (IAvH - 01285). Casanare. Yopal, 5º21´N; 72º24´W, 350 m, bmh-T, 1 (H), maleza de hoja ancha, 3.V.1970, M. Camacho (FAUNIC 04, UNAB). Cauca. Morales, 2°45´27”N; 76°37´56”W, 1635 m, 1 (M), 10. VI.1988, L. Mera (UNAB); PNN Munchique, 1750 m, 2 (M), bosque, 2.II.1998, P. Rengifo (IAvH - 01372, 01363); El Tambo, Tambito, 1 (M), XI.1990, A. J. Negret (EN 000233 R, MHN-UC); Popayán, Barrío Los Hoyos, 1 (M), 5.IV.1982, H. Granados (EN 000234 R, MHN-UC). Chocó. Nuquí, Jurubidá, Comunidad Embera, 2 (M), manual en bosque primario, 4.X.1995, B. Amagara (0102-951004-0020, 0102-951004- 0019, CIB); Nuquí, Jurubidá, Morro-Mico, 1 (H), manual intradomicilio, 23.VII.1995, G. Tabares, (0101-950723-0004, CIB); Nuquí, Antacorí, 1 (M) y 1 (H), jama en rastrojo, 15.I.1996, H. Jaramillo (0100-960115-0012, -0013, CIB); Nuquí, Tribugá, Río Tribugá arriba, 1 (M), jama en rastrojo, 6.XII.1995, J. H. Murillo (0102-951206-0009, CIB); Nuquí, Estación El Amargal, 30 m, 2 (H), 22.VIII.1996, C. C. Ramírez (ICNMHN- CO 4657, 4654, ICN); Nuquí, El Amargal, 5 m, 1 (H), Malaise, II.1994, A. Bonilla (IAvH - 01280); Lloró, Quebrada Boraudó, 1 (M) y 1 (H), trampa de luz, 17-20.III.2002, Neita J. C. (ICN-MHN-CO 7563, 7564, ICN); Rio Sucio, Caño Seco, 1 (M), 26.III.1999, Carlos Sierra (ICN-MHN-CO 7557, ICN); B. San Juan, Togoromá, 1 (H), Colecta diurna en follaje, 22.VII.1990, L. C. Pardo Locarno (CFPL). Cundinamarca. Santandercito, 1 (H), 10.I.1965, S. Restrepo (MUJ - 5921); Santandercito, 1600 m, 1 (M), III.1964, sin colector (MUJ - 5919); Santandercito, 1 (M), II.1965, S. Restrepo (MUJ - 5918); San Juan de Río Seco, 4°50´54”N; 74°37´35”W, 1798 m, 1 (M), manual, cultivo de café-guayaba, 19.V.2003, A Mendoza y J. Cárdenas (UNAB); Cáqueza, 4°24´30”N; 73°56´58”W, 1746 m, 1 (M), 18.III.1995, Ivan P. (UNAB); Pacho, Pan de Azúcar, 5°07´57”N; 74°09´42”W, 1798 m, 1 (H), sobre un tronco, 12.IV.2004, N. Robayo y J. Girón (UNAB); Guayabal de Síquima, 4°52´54”N; 74°28´59”W, 1200 m, 1 (H), manual en café, 25.IV.2004, J. Escalante y M. Y. Romero (UNAB); Tena, Pedro-Palo, 1200-2400 m, 1 (M), sin fecha de colección, grupo #1 (MUJ - 5926); Cachipay, Vereda Naranjal, Finca Microguaca, 1800 m, 1 (M), jama en cítricos, 10.V.2003, C. Jerez (UNAB); Tudela, 1 (M), 1.IV.1996, C. Díaz (FAUNIC 06, UNAB); El Trigo, 1 (H), 12.IV.1997, Paola Serna (FAUNIC 05, UNAB); La Vega, 4°59´57”N; 74°20´23”W, 1230 m, 1 (H), 18.VI.1996, A. Mejía (FAUNIC 08,UNAB); San Antonio, Vereda Chicaque, 4°29´06”N; 73°53´24”W, 1867 m, 1 (H), 6. XII.1997, G. Sánchez (FAUNIC 02, UNAB); Cachipay, 1600 m, 1 (M), bajo tronco, 5.XII.1996, C. Martínez (ICN-MHN-CO 7558, ICN); Albán, 2200 m, 2 (M), Granjas del Padre Luna, 9. II.2000 y 24.II.2000, G. Amat (ICN-MHN-CO 7561, 7556, ICN). Magdalena. Santa Marta, Minca, 1 (M), 9.IV.1977, grupo 2 (ICN-MHN-CO 2331, ICN); Santa Marta, Sierra Nevada de Santa Marta, 1 (M), 15.II.1995, C. Sarmiento (ICNMHN- CO 7553, ICN). Meta. Macarena, Caño Cabra, Río Guayabero, 1 (H), 22.II.1969, P. Pinto (ICN-MHN-CO 2332, ICN); El Dorado, Vereda Santa Rosa, Caño Paujil, 550 m, 1 (M), sin fecha de colecta ni colector (ICN-MHN-CO 4638, ICN); San Juan de Arama, PNN La Macarena, La Curia, Caño La Curia, 580 m, 1 (M) y 1 (H), manual, II.1989, F. Fernández (IAvH - 01279, 01287); Macarena, PNN Tinigua, Pto. Chamuza, Rio Duda, CIEM, 2°40´N; 74°10´W, 350 m, 1 (M), 1994, M. Álvarez y A. Calixto (IAvH - 01367). Nariño. Barbacoas, Corregimiento Altaquer, Reserva Natural privada Río Ñambí, 1 (H), VII.1995, sin colector (MUJ - 5922). Norte de Santander. Pamplona, 1 (H), 1972, Hno. Ramón Guevara (MSL - 1854). Quindío. Filandia, Bremen C.R.Q., 1 (M), recolección directa, 21.IV.1998, V. Cruz et.al. (MUJ - 5838); Filandia, Estación Bremen C.R.Q., 1800-1900 m, 3 (H) y 1 (M), luz blanca en bosque de pino, en camino, manual, bosque nativo, 14-20.IV.1998, P. Maldonado, Martha Fonseca, Berrío et al . (MUJ CP 0159, 0158, 0328, 0162, MUJ); Circacia, 4°37´21”N; 75°38´14”W, 1805 m, 1 (H), al vuelo, 29.V.1975, J. Carvajal (FAUNIC 01, UNAB). Risaralda. Pereira, 1 (H), en suelo, 18.IX.1996, Escobar (LEUC); Ucumarí, La Suiza, 1900 m, 1 (H), 13.V.1991, Germán Amat (MUJ - 5927); Ucumarí, La Suiza, 1995 m, 1 (H), VIII.1992, G. Amat (MUJ - 5920). Santander. Charalá, Virolín, 1 (H), 7.III.1981, sin colector (ICN-MHN-CO 5853, ICN); Virolín, carretera Duitama- Charalá, 1750 m, 1 (H), 9.III.1981, sin colector (ICNMHN- CO 2828, ICN); Vélez, Ciénaga. Oph., 1 (M), L. Richter (ICN-MHN-CO 2893, ICN); S. Claver, 1 (M), 25.X.1963, R. Cortés (MUJ - 5915); Enciso, Vereda Tierra Amarilla, 6°40´16”N; 72°42´08”W, 1580 m, 1 (H), 22.III.1997, Gutiérrez (FAUNIC 09, UNAB); La Belleza, 2240 m, 1 (H), alrededores del casco urbano, VI.2004, Estudiantes Colegio Juan Bosco (ICN-MHN-CO 7559, ICN); Tona, Sector el Brasil, Quebrada Brasil, 1600 m, 1 (H), 20.VI.2004, G. Amat y Vélez (ICN-MHN-CO 7554, ICN). Tolima. Villahermosa, 1 (M), XII.1941, F. L. Gallego (No. 8771, MFLG); Villa Rica, 1 (M), 22.IX.1997, Bustos-Mesa (UNAB). Valle del Cauca. Cali, 1 (H), en arvenses, 13.II.1996, Villa (LEUC); Cali, Carretera al mar, 1 (H), VI.1970, L. Ángel (ICN-MHN-CO 2333, ICN); Cali, Mares, Cordillera Occidental, 3 (M) y 2 (H), 3.VI.1968 y VII.1968, Humberto Salazar (MSL - 607, 608, 609, 1862, 1856); Palmira, Amaime, arenillo, 1100 m, 1 (M), III.1990, J. Carana (MUSENUV - 6394); Cali, 1000 m, 1 (H), IV.1980 (MUSENUV - 6399); PNN Farallones, El Topacio, 1800 m, 1 (H), sin fecha de colección ni colector (MUSENUV - 6406); Alto Anchicayá, 500 m, 1 (M), III.1983, R. Astaiza (MUSENUV - 6395); Cali, Peñas Blancas, 1500-1900 m, 1 (M) y 2 (H), III.1987 y 9.III.1990, J. Montealegre, L. F. Fory y H. Rubio (MUSENUV); Dagua, Queremal, 1 (H), X.1985, Martha L. Baena (MUSENUV - 21973); Buenaventura, Central Hidroeléctrica Anchicayá, 400 m, 1 (H), fuente de luz, 6.I.1989, Claudia A. Medina (MUSENUV); Cali, San Antonio, 2200 m, 1 (H), Trampa de luz, 10.VI.1991, Ximena Cáceres (MUSENUV); Alto Anchicayá, 680 m, 1 (H), vegetación, 17. IX.1997, Raúl Gallego (MUSENUV); Tuluá, Mateguadua, 1200 m, 1 (H), trampa de luz, 22.X, M. P. Aguirre (MUSENUV); Central Hidroeléctrica Anchicayá, 300 m, 2 (M), trampa de luz, 10.XI.1991, E. Reyes (MUSENUV); Chicoral, 1700-1800 m, 2 (M), maleza y tronco en descomposición, 26. II.1992 y 17.XI.1992, G. Villegas y F. Lozano (MUSENUV); Bajo Anchicayá, 270 m, 1 (M), trampa de luz, 3.IV.1992, Vicky Vélez (MUSENUV); San Antonio, 2600 m, 1 (M), 2.X.1984, Nancy Carrejo (MUSENUV); San Antonio, Km 18, 1870 m, 1 (M), tronco en descomposición, 1.VI.1995, A. Montoya, (MUSENUV); M. Calima, Cristalina c/Alegre, 1 (M), luz, X.1990, C. Escobar y otros (CFPL); Málaga, B. Málaga, 1 (M), 1991, H. A. Rodríguez (CFPL); B. Dagua, Escalarete, 1 (M) y 3 (H), luz, 25.XII.1990 a 15.III.1991, L. Rivas (CFPL); M. Calima, La Palmera, 2 (M) y 1 (H), Colecta diurna en follaje, 10-12. XI.1990, L. C. Reyes (CFPL); M. Calima, Lago Calima, 1 (H), luz, 6.III.1987, L. C. Pardo Locarno (CFPL); M. Calima, Q. Cristalina Alta, 1 (M), Colecta diurna en follaje, 21-28.VII.1992, varios colectores (CFPL); R. Bravo, La Palmera Var. Just, 1 (M), 15.X.1990 a 7.XI.1990, A. Papamija (CFPL); Saladito, C. San Antonio, 1 (M), Colecta diurna en follaje, VI.1985, L. C. Pardo Locarno leg. (CFPL). Localidades inciertas. sin datos de localidad, 1 (H), X.1947, F. L. Gallego (No. 8771, MFLG); Rio Negrito, 2000 m, 1 (M), VI.1951, F. L. Gallego (No. 8771, MFLG).
Parandra (Parandra) humboldti (Santos-Silva, 2003)
Hesperandra (Zikandra) humboldti Santos-Silva, 2003a: 123; Monné & Hovore 2006: 5 (cat.); Monné, 2006: 14 (cat.); Santos-Silva, 2007: 5, 9 (clave). Localidad-tipo: Colombia, Risaralda: Santuarío Campamento (2200 m). (IAvH).
Parandra (Parandra) humboldti; Santos-Silva & Shute, 2009: 31 (comb. nov.).
Diagnosis. Aspecto brillante (Fig. 3A); puntuación imperceptible. Cabeza grande (tan larga como el pronoto en especímenes de tamaño promedio) (Figs. 3B y 3C). Submento con puntuación muy fina y dispersa (Fig. 3D); pilosidad escasa en la región centro-anterior, microscópica en las regiones laterales; borde anterior elevado, precedido por un surco poco profundo. Mandíbulas fuertemente falciformes y prominentes en los machos, subtriangulares en las hembras (Figs. 3B y 3C). Antenas alcanzando el borde posterior del protórax (Fig. 3E); antenómeros III-XI con el área sensorial dividida por una carena completa y alta. Pronoto liso (Fig. 3F). Metepisternos con puntuación inconspicua, cubiertos completamente de pelos evidentes, largos, y abundantes (Fig. 3G). Metaesterno liso en la región central; con puntos muy pequeños y pelos claramente visibles en las áreas adyacentes a las mesocoxas y a los metepisternos (Fig. 3H). Élitros lisos (Fig. 3I).

Distribución. Colombia (Fig. 10B) y Ecuador.
Comentarios. Esta especie se diferencia de P. glabra porque los metepisternos están cubiertos de pelos largos y abundantes. En general las hembras no presentan ninguna puntuación en el submento, aunque sí algunos pelos dorados y largos. Se ha colectado en seis departamentos en Colombia, en las tres cordilleras a alturas por encima de los 500 m. Adicionalmente, Santos-Silva (2003a) registró un espécimen del Meta, depositado en el Museo de Historia Natural de la Universidad Nacional de Colombia (Bogotá). Al igual que otras especies que poseen pilosidad abundante en los esternitos, habita principalmente regiones montañosas (Santos-Silva 2003b).
Material examinado. COLOMBIA. Antioquia. Andes, Corregimiento Santa Rita, Vereda La Soledad, Finca La Reina, 05°35´17,6”N; 075°55´46,0”W, 2010 m, 1 (M), red de niebla, bosque, 2.X.2001, J. Cardona-D. (CEUA -27114); Caldas, 1750 m, 1 (M), manual, en suelo, IX.1973, R. Vélez (No. 4508, MFLG); San Jerónimo, 1 (M), manual en maleza, 30.IV.1986, Patricia Duque V. (No. 4508, MFLG); Caicedo, Vereda La Noque, 2200 m, 1 (M), manual, V.2009, L. M. Urrea (CEUA - 44577); Medellín, Corregimiento de San Cristóbal, Vereda La Palma, 3 (M), manual, III.2008, G. Marín (CEUA - 44564, 44565, 44566). Boyacá. Bravita, 2 (M), 25.V.1972, Hno. Antonio Camilo (MSL - 1953, 1864). Caldas: Palestina, Vereda Santágueda, 1 (M), en arvenses, 18.XII.1978, Bolaños (LEUC). Huila: Gigante, 4 (M) y 1 (H), directa, 16.X.1993, Héctor Bonilla (CFPL). Valle del Cauca. Corregimiento La Horqueta, 2000 m, 1 (H), suelo, 14.III.1994, F. López (MUSENUV - 6396); Km 18 al mar, 1800 m, 1 (H), tronco de árbol, 24.I, J. Castillo (MUSENUV - 6397); Peñas Blancas, 1500 m, 1 (M), III.1993, M. R. Manzano (MUSENUV - 6398); Antena T.V., 2030 m, 1 (H), IV.1978, Alberto Gómez (MUSENUV); La Cumbre, Dapa Chicoral Alto, 1970 m, 2 (H), tronco en descomposición, 6.V.1989, Yolanda Gutiérrez (MUSENUV); San Antonio, Antena, 2100 m, 1 (H), 27.III.1998, F. Molano (MUSENUV - 21971); Cali, Pance, El Topacio, 2000 m, 1 (M), barrido con jama, IV.1990, F. Escobar I. (MUSENUV - 21969); La Cumbre, Dapa Chicoral Alto, 1970 m, 1 (M), tronco en descomposición, 6.V.1989, A. M. Astudillo (MUSENUV - 21972); Ca Saladito, 1800 m, 1 (M), V.1976, sin colector (MUSENUV); Ca Cali, PNN Farallones, 2200 m, 1 (M), tronco en descomposición, 17.IV.1990, Pablo E. Ruiz (MUSENUV - 21970); San Antonio, Jamundí, 1 (H), 7.III.1983, L. C. Pardo Locarno (CFPL); Saladito, C. San Antonio, 3 (M) y 1 (H), Colecta diurna en follaje, 1990 y V.1985, L. C. Pardo Locarno (CFPL); M. Calima, La Palmera, 1 (M) y 2 (H), Colecta diurna en follaje, 10-12.XI.1990, L. C. Reyes (CFPL). Risaralda. Santuario, Campamento, 2200 m, HOLOTIPO 1 (M), 1.III.1990, F. Fernández (IAvH - 01282).
Parandra (Tavandra) Santos-Silva, 2003
Hesperandra (Tavandra) Santos-Silva, 2003a: 125; Monné & Hovore, 2006: 4 (cat.); Monné, 2006: 10 (cat.).
Parandra (Tavandra); Santos-Silva & Shute, 2009: 31.
Especie-tipo. Parandra longicollis Thomson, 1861 (designación original).
Proyección del borde ocular posterior sobresaliente. Submento deprimido principalmente en los machos, con pelos conspicuos que pueden ser abundantes, puntuación gruesa (puede estar anastomosada), margen anterior no elevado en los machos, delgado y elevado en las hembras. Carena ventral de las genas delgada y elevada, generalmente como consecuencia de la depresión del submento. Área sensorial de los antenómeros III-XI, dividida por una carena.
Clave para las especies de Parandra (Tavandra) presentes en Colombia
1. Metepisternos con pelos evidentes y concentrados (Fig. 5G) ...............................................................................2
- Metepisternos glabros o con pelos dispersos (Fig. 6D) 4 2(1). Metaesternos con pilosidad menos concentrada que en los metepisternos (pelos un poco más abundantes en la parte anterior, cerca a las mesocoxas y metepisternos). Ápice del labro agudo................................................P. (Tavandra) villei Lameere, 1885
- Metaesternos con pilosidad igual a los metepisternos.
Ápice del labro redondeado...................................................3 3(2). Puntuación elitral gruesa (Fig. 4H). Diente del margen interno de las mandíbulas de los machos con el ápice redondeado, localizado en la mitad apical...................P. (Tavandra) colombica White, 1853
- Puntuación elitral fina (Fig. 5I). Diente del margen interno de las mandíbulas de los machos con el ápice agudo, localizado en el medio......P. (Tavandra) scaritoides Thomson, 1861
4(1). Dientes del margen interno de las mandíbulas de los machos, localizados en el medio o un poco hacia la base (Fig.6A); margen interno de las mandíbulas de las hembras cóncavo............................................ P. (Tavandra) polita Say, 1835
- Dientes del margen interno de las mandíbulas de los machos, localizados en el tercio apical; margen interno de las mandíbulas de las hembras, con una concavidad en el centro .............................. P. (Tavandra) longicollis Thomson, 1861
Parandra (Tavandra) villei Lameere, 1885
Parandra villei Lameere, 1885a: xi. Localidad-tipo: Ecuador (ISNB, SMTD, BMNH).
Parandra (Tavandra) villei; Santos-Silva & Shute, 2009: 31 (comb. nov.).
Hesperandra (Tavandra) villei; Monné, 2006: 12 (cat.); Monné & Hovore, 2006: 4 (cat.).
Diagnosis. Color del tegumento castaño o castaño claro. Cabeza grande (tan larga como el pronoto en machos grandes, un poco más corta en machos pequeños y hembras). Submento con puntos gruesos y anastomosados; margen anterior no elevado. Mandíbulas claramente aguzadas hacia el ápice; diente del margen interno localizado en el medio. Protórax del macho levemente transversal, a veces suavemente subcuadrado; en la hembra más transversal. Metaesterno glabro y liso. Metepisternos con pelos dispersos. Élitros finamente punteados.
Distribución. Colombia (Fig. 10C), Ecuador, Perú y Venezuela.
Comentarios. De acuerdo con Arigony (1970) existen 18 síntipos: 13 (M) y 5 (H), y de esos, 11 machos y 5 hembras estarían depositados en el ISNB, un macho en el SMTD y un macho en el BMNH. Adicionalmente Arigony (1970) afirmó que designó el lectótipo y paralectótipos de la série sintípica, de ejemplares rotulados con “Type” o “Ex-Typis”, algunos con etiquetas manuscritas de Lameere. Escogió como lectotipo el macho en mejor estado de preservación, con el rótulo manuscrito que lo designa como “Type” y los demás quedaron rotulados como paralectotipos (Arigony 1970). Aunque Arigony (1970) no mencionó la instutuición depositária del lectotipo, la designación es conforme a la segunda versión del ICZN (1964) válido en la época.
Sobre la instituición todavía caben algunos comentarios: Lameere (1885a) escribió: “Typi in Mus. reg. Belg. atque nostro, (M) (H)”. No sabemos cuántos ejemplares pertenecían originalmente a la coleción Lameere, ni dónde están realmente depositados. Aunque Arigony (1970) señaló que había sintipos del SMTD y del BMNH, no mencionó que revisó material de éstas instituciones: “Este trabalho tem por objetivo ampliar a descrição original de Parandra (Archandra) villei Lameere, 1885, com base na série sintípica, recebida por deferência de R. Damoiseau do Institut Royal des Sciences Naturelles de Belgique”. Damoiseau y Cools (1987) en su lista de tipos del ISNB escribieron: “Lectotype & 14 paralectotypes, (6), Equateur: E. de Ville, 1871; designes par T. Arigony, 1970, Rev. Bras. Ent. 14(8): 58”. Aunque especifican dónde está depositado el “lectótipo”, las cantidades de “paralectótipos” no concuerdan: de acuerdo con Arigony (1970), el total depositado en el ISNB es 16 tipos, por lo tanto serían 15 “paralectótipos”; adicionalmente The Natural History Museum de Londres, corrobora que hay un tipo depositado en dicha institución (Pitkin 2003).
Por otro lado, Rosales (1966) estudió dos machos que consideró erróneamente Parandra (Archandra) villei. A pesar de la descripción detallada y la fotografia que presentó, no es posible determinar la especie a la cual se refiere, que difere de P. (T.) villei porque el diente del margen interno de la mandíbula es pequeño y los metepisternos son glabros [aunque Rosales (1966) no afirmó que eran glabros, inferimos que los son dado que sus descripciones eran muy detalladas y en este caso no especificó algo diferente]. En Parandra villei el diente del margen interno es bien desarrollado y los metepisternos tienen pilosidad dispersa y evidente.
Arigony (1970) escribió que el margen interno de las mandíbulas tenía pelos en la base hasta el primer diente, que es pequeño y se encuentra más allá del medio. Sin embargo, el macho que examinamos y las fotografias de uno de los síntipos machos presentadas por Arigony (1970), muestran claramente que el diente es bien desarrollado. Posteriormente, Arigony (1978) afirmó haber examinado un macho proveniente de Venezuela y dos machos de Colombia, citando: “COLÔMBIA, Antioquia: Medellín. 1 (M) (MNHN). Caldas: Manizales. 1 (M) (MNHN).”. Es probable que la especie realmente esté presente en Colombia.
Material examinado. ECUADOR. Napo. Baeza (2000m), 1 (M), III.1979, M. Kaulbars col. (MZSP); Província (?): Santo Domingo [existen varias localidades con ese nombre en Ecuador (Falling Rain Genomics 1996-2006)], 1 (H), II.1965, Peña col. (MZSP).
Parandra (Tavandra) colombica White, 1853
Parandra colombica White, 1853: 3. Localidad-tipo: Colombia. (BMNH).
Parandra (Tavandra) colombica; Santos-Silva & Shute, 2009: 31 (comb. nov.).
Hesperandra (Tavandra) colombica; Martínez, 2000: 83 (distr.); Monné, 2006: 11 (cat.); Monné & Hovore, 2006: 4 (cat.).
Parandra Gyllenhalii Thomson, 1867: 110. Localidad-tipo: Venezuela. (MNHN).
Diagnosis. Cabeza mediana (más corta que el pronoto en especímenes de tamaño promedio), con puntos relativamente finos y poco concentrados (Figs. 4A y 4B). Submento con puntos gruesos y anastomosados (en las hembras pueden ser un poco más dispersos), con pelos abundantes, más concentrados en dirección a las genas (Figs. 4C y 4D). Mandíbulas de los machos con diente del margen interno grande, de ápice redondeado y localizado en el tercio basal (aunque puede estar cerca de la región central) (Fig. 4A); en las hembras (Fig. 4B). Protórax subcuadrado (Fig. 4E) (en la parte anterior, puede ser un poco más ancho que el largo total, y un poco estrechado hacia la parte posterior, dando la apariencia de ser trapezoidal). Metepisternos con pilosidad abundante y conspicua (Fig. 4F). Metaesterno con pilosidad similar al metepisterno en las regiones laterales, desprovisto de pelos en la región central (Fig. 4G). Élitros con puntuación muy gruesa (Fig. 4H).

Distribución. Colombia (Fig. 10B), Ecuador y Venezuela.
Comentarios. Esta especie se distingue de Parandra scaritoides Thomson, 1861 por la puntuación elitral más gruesa, la forma de las mandíbulas que no se angostan abruptamente hacia el ápice, el diente medio de las mandíbulas redondeado y la forma del pronoto en los machos, subcuadrado en P. colombica (aproximadamente tan ancho como largo) y transversal en P. scaritoides (un poco más ancho que largo).
Aunque las hembras de P. colombica y P. scaritoides no son fácilmente distinguibles, la puntuación elitral en las hembras de P. colombica es tan gruesa y abundante como en los machos, mientras que en P. scaritoides la puntuación es dispersa, más fina y rasa. Además en P. scaritoides los márgenes laterales del pronoto son paralelas en toda su extensión, mientras que en P. colombica son suavemente divergentes hacia la región anterior. Por otro lado, en P. colombica algunas hembras pueden presentar pelos cortos restringidos a la región lateral del submento (contiguos a la carena ventral de las genas).
Aunque existe un registro de esta especie para Nicaragua (Bates 1872), el estudio de una gran cantidad de parandrinos de Centroamérica por Santos-Silva, no ha revelado la presencia de esta especie en esa área. Aunque Arigony (1978) registró esta especie para los departamentos de Antioquia, Caldas, Cauca, Cundinamarca y Valle del Cauca en Colombia, en 1978 P. scaritoides estaba bajo sinonimia de P. colombica. En las colecciones colombianas sólo se encontraron especímenes de P. colombica recolectados en Boyacá y Huila.
Material examinado. COLOMBIA. Boyacá. Arcabuco, 2600 m, 1 (H), 11.V.1996, J. C. Mendoza (ICN-MHN-CO 4655, ICN); Villa de Leyva, 5°38´19”N; 73°31´42”W, 2200 m, 1 (H), manual en calle del pueblo, 11.II.1999, C. Martínez (IAvH - 01910); Villa de Leyva, 5°38´N; 73°31´W, 2200 m, 1 (M), luz pueblo, 10.X.1996, F. Fernández (IAvH - 01365); Villa de Leyva, 2200 m, 1 (H), XI.1996, M. Baena (IAvH - 01284); SFF Iguaque, vía Carrizal, 2550 m, 1 (M), manual en robledal en camino, sin fecha de colecta, Trujillo D. (ICN-MHN-CO 7562, ICN). Huila. Algeciras, El Paraíso, Termópilos, 2200 m, 1 (H), II.1999, D. Trujillo (ICN-MHN-CO 6298, ICN); Gigante, directa, 16.X.1993, Héctor Bonilla (CFPL).
Parandra (Tavandra) scaritoides Thomson, 1861
Parandra scaritoides Thomson, 1861b: 82. Localidad-tipo: Colombia, Cartagena. (MNHN).
Parandra (Tavandra) scaritoides: Santos-Silva & Shute, 2009: 31 (comb. nov.).
Hesperandra (Tavandra) scaritoides; Monné, 2006: 12 (cat.); Monné & Hovore, 2006: 4 (cat.).
Diagnosis. Cabeza grande (tan larga como el pronoto en especímenes de tamaño promedio) (Figs. 5A-5C). Frente con puntos muy pequeños y dispersos; vértex convexo con puntos pequeños, un poco mayores que los de la frente e igualmente dispersos. Submento con el margen anterior plano, glabro y sin puntuación, o suavemente elevado y con puntos pequeños y dispersos (Fig. 5C). Mandíbulas de los machos con diente del margen interno grande, agudo y localizado aproximadamente en el medio (Fig. 5D); en las hembras con una fuerte invaginación en el margen interno (Fig. 5E). Área sensorial de los antenómeros con carena sobresaliente; antenómeros II-XI con pelos cortos en el área ventro-apical, y V-X con algunos pelos cortos dorsales. Protórax transversal (levemente más ancho que largo) (Fig. 5F). Pronoto con puntos pequeños y dispersos en el área central, más concentrados en las regiones laterales. Metepisternos con pilosidad abundante y conspicua (Fig. 5G). Metaesterno con puntos y pelos similares al metepisterno en las regiones laterales, desprovisto de pelos en la región central (Fig. 5H). Abdomen con tubérculos pequeños de los cuales emerge un pelo. Élitros con puntos pequeños, conspicuos y concentrados (Fig. 5I). Tibias dilatadas apicalmente.
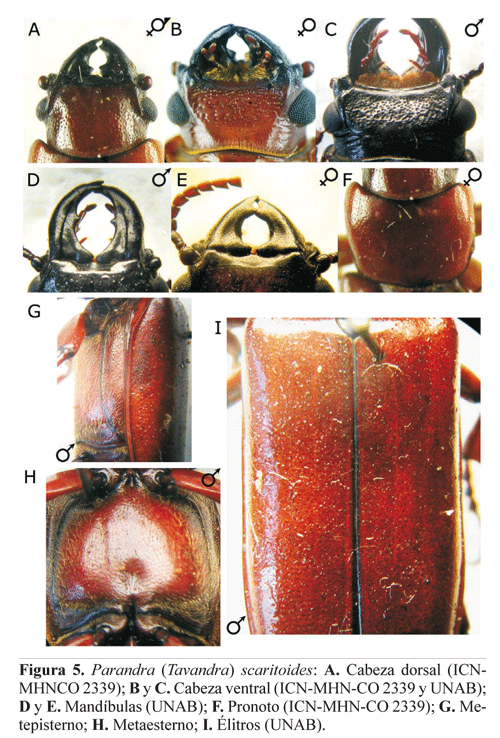
Distribución. Colombia (Fig. 10B).
Comentarios. Esta especie fue colocada bajo sinonimia de P. colombica por Lameere (1902), quien se basó únicamente en la descripción sucinta de Thomson (1861b) y no revisó los tipos que actualmente se encuentran en el MNHN. Santos- Silva (2003b) revalidó la especie presentando una serie de caracteres que la diferencian de P. colombica: Parandra scaritoides posee puntuación elitral más fina, el ápice del diente del margen interno de las mandíbulas agudo, la forma de las tibias un poco dilatada apicalmente y el pronoto transversal con márgenes laterales paralelos. La puntuación del submento varía y generalmente es más fina en las hembras. Igualmente el grado de protuberancia del borde ocular posterior puede variar.
Aunque Bates (1879) registró P. scaritoides para Nicaragua, Santos-Silva, a pesar de haber estudiado una gran cantidad de especímenes de Centroamérica, sólo ha revisado ejemplares provenientes de Colombia.
Biología. Mendoza (com. pers.) colectó uno de los especímenes examinados bajo la corteza de un sauce (Salix sp.), mientras se alimentaba de la madera en un árbol que aún se encontraba en pie (con algunas ramas y hojas verdes).
Material examinado. COLOMBIA. Boyacá. Paipa, La Pradera, 2650 m, 1 (H), 2.XII.1978, Z. Arévalo G. S. (ICN-MHNCO 2986, ICN); Chiquinquirá, 1 (H), 13.II.1969, J. Norato R. (ICN-MHN-CO 2339, ICN); Guacamayas, cerca de la cabecera municipal, 1 (M), en un tronco de Sauce, de una galería, sin fecha de colección, G. Mendoza (GMIC); Otanche, 1 (M), sin fecha de colecta ni colector (MSL - 1858); Chita, 1 (M), 20.II.1993, L. Lizarazo (UNAB). Cundinamarca. Une, Vereda Combura, 2600 m, 1 (H), manual en cultivo de hortalizas, 27.X.2002, sin colector (UNAB). Magdalena. Fundación, Finca “El Paraíso”, 10°31´N; 74°11´W, 45 m, 1 (H), jama en pastos, 14.I.2004, A. Osorío (UNAB). Norte de Santander. Pamplona, 4 (M), Páramo, sin fecha de colecta ni colector (MSL - 1861, 1865, 1863, 1855); Pamplona, 5 (M) y 2 (H), sin fecha de colecta ni colector (MSL - 1848, 1849, 1857, 1859, 1867, 1850, 1852). Valle del Cauca. San Antonio, 2600 m, 1 (M), fuente de luz, 7.IV.1997, T. Lozano (MUSENUV - 7951); Km 12, 1100 m, 1 (H), 1975, sin colector (MUSENUV - 6405); San Antonio, 1800 m, 1 (H), 10.VII.1988, B. Quesada (MUSENUV).
Parandra (Tavandra) polita Say, 1835
Parandra polita Say, 1835: 192; Yanega, 1996: 25; Jeniš, 2001: 19. Localidad-tipo: Estados Unidos, Indiana. (Institución depositaria desconocida).
Parandra (Tavandra) polita; Santos-Silva & Shute, 2009: 31 (comb. nov.)
Hesperandra (Tavandra) polita; Monné, 2006: 11 (cat.); Monné & Hovore, 2006: 4 (cat.).
Parandra cylindrica Thomson, 1861b: 82. Localidad-tipo: México. (MNHN).
Diagnosis. Cuerpo subrectangular (con los lados fuertemente paralelos). Cabeza pequeña (más corta que el pronoto en especímenes de tamaño promedio), con puntuación débil (puntos pequeños y rasos) y homogénea dorsalmente (Fig.6A). Submento con puntuación gruesa (Fig. 6B). Mandíbulas en los machos con puntuación más fina y concentrada que en la cabeza (Fig. 6A); diente del margen interno de las mandíbulas conspicuo, localizado cerca al medio o un poco más hacia la base; margen interno de las mandíbulas de las hembras cóncavo. Protórax sin puntos, se estrecha gradualmente hacia la parte posterior (Fig. 6C). Metepisterno glabro, con puntos finos y dispersos (Fig. 6D). Metaesterno glabro y sin puntuación (Fig. 6E). Élitros con puntuación diminuta (Fig. 6F).

Distribución. Estados Unidos, México, Centroamérica, Guyana, Colombia (Fig. 10C), Ecuador, Perú, Bolivia, e Inglaterra (introducida) (Santos-Silva 2003a).
Comentarios. Parandra polita es una especie de amplia distribución, sin embargo en las colecciones del país sólo se halló un ejemplar colectado en la costa pacífica a una altura inferior a 100 m. Adicionalmente Arigony (1978) registró un ejemplar de Medellín ((H)) que se encuentra depositado en el MNHN.
Material examinado. COLOMBIA. Chocó. Nuquí, Jurubidá, 1 (M), manual intradomicilio, 6.VI.1994, J. Zuluaga (CEUA - 36694).
Acutandra Santos-Silva, 2002
Acutandra Santos-Silva, 2002: 49; Monné, 2006: 15 (cat.); Monné & Hovore, 2006: 8 (cat.)
Especie-tipo - Parandra punctatissima Thomson, 1861 (designación original).
Cabeza mediana, transversal o suavemente longitudinal; borde ocular posterior generalmente sobresaliente. Mandíbulas poco desarrolladas, subtriangulares, túmidas en la cara externa, semejantes en los dos sexos. Ápice del labro delgado y acuminado, semejante en ambos sexos. Área sensorial dorsal del antenómero XI clara o debilmente delimitada. Área sensorial ventral de los antenómeros III-XI entera o dividida por una carena. Pronoto moderada o fuertemente punteado; borde anterior del pronoto, en general, fuertemente sinuoso en ambos sexos. Élitros con puntuación fuerte y concentrada. Metaesterno y metepisternos glabros. Cavidades coxales anteriores abiertas en la parte posterior. Paroniquio desarrollado.
Todas las especies de este género se distribuyen en Suramérica.
Acutandra ubitiara (Santos-Silva y Martins, 2000)
Parandra (Parandra) ubitiara Santos-Silva y Martins, 2000: 167. Localidad-tipo: Colombia, Chocó, Bajo San Juan (CPFL).
Acutandra ubitiara; Monné, 2006: 9 (cat.). Acutandra punctatissima; Santos-Silva, 2002: 51, fig. 52 (error).
Diagnosis. Macho: cabeza mediana (tan larga como la mitad de la longitud del pronoto en especímenes de tamaño promedio), borde anterior oscurecido (Fig. 7A); frente glabra con puntos gruesos. Submento con puntos dispersos, sin puntos en la región posterior (Fig. 7B). Ojos grandes. Antenas con el área sensorial de los antenómeros III a XI dividida por una carena longitudinal baja (Figs. 7C y 7D). Pronoto convexo, levemente transversal (Fig. 7E); borde anterior fuertemente sinuoso; región anterior con círculos irregulares en relieve, cada uno con una depresión central puntiforme (escultura más acentuada en la región latero-anterior), área centro-anterior sin círculos (Fig. 7F); ángulos posteriores obtusos. Proesterno glabro, con puntos mayores en la región lateral; área central cerca del borde anterior ligeramente arrugada (Fig. 7G). Urosternitos I a IV glabros, puntos gruesos más concentrados en las áreas laterales (Fig. 7H). Élitros con puntos gruesos irregularmente distribuidos. Hembra. Similar al macho pero sin la escultura de círculos en relieve en la parte anterior del pronoto (Fig. 7I).

Distribución. Colombia (Fig. 10C).
Comentarios. Aunque Acutandra ubitiara sólo se conoce para unas pocas localidades en la región Pacífica de Colombia, es probable que se encuentre en otras regiones del suroccidente del país. En la descripción Santos-Silva y Martins (2000) reportan que el holotipo está depositado en el Museo de Ciencias Naturales de Cali y que fue enviado en 1991 por Luís Carlos Pardo Locarno para su identificación. Sin embargo la institución depositaria del holotipo realmente es CFPL que se encuentra en Palmira, Valle.
Por otro lado la localidad típica registrada en la descripción es: “Colômbia, Valle del Cauca: Bajo Calima, L.C. Pardo Locarno col., ‘S12-083´”. Pardo Locarno (com. pers.) dice que todos los individuos de Acutandra que están en CFPL fueron colectados por él y sus compañeros durante un proyecto en la región de Calima (Valle del Cauca y Chocó). El código que aparecía en la etiqueta del holotipo (S12-083) también estaba en las etiquetas de los otros especímenes de Acutandra, los cuales además tenían dos etiquetas (Fig. 7J) (aunque el holotipo y el paratipo sólo tenían una etiqueta con los datos que constan en la descripción original). Los especímenes con este código se colectaron durante la “salida 12 del proyecto Coleópteros del Calima (COLCIENCIAS-INCIVA) realizada al Bajo San Juan, Chocó” (Pardo Locarno com. pers.). De acuerdo con Pardo Locarno el dato correcto de localidad y en consecuencia la localidad típica es: COLOMBIA, Chocó, Bajo San Juan, Julio de 1990, L.C. Pardo Locarno leg. (CPFL).
Por otro lado, sólo se conocían tres especímenes de A. ubitiara: el holotipo depositado en CFPL, un paratipo depositado en MZSP y un espécimen que Santos-Silva (2002) reportó depositado en el Museo de Historia Natural de la Universidad Nacional de Colombia en Bogotá. Este último en realidad pertenece al IAvH y la localidad registrada en la etiqueta no es “Valle del Cauca” sino: ‘Cauca, I.[Isla] Gorgona, Sendero Yundigua, 40 m´, siendo el primer registro de un Parandrinae para esta isla del Pacífico colombiano.
La hembra registrada por Santos-Silva (2002: Fig. 5) como A. punctatissima realmente corresponde a la hembra de A. ubitiara y difiere de ésta principalmente por la forma de los ángulos posteriores del pronoto. Aunque todos los ejemplares conocidos de A. ubitiara eran machos, en CFPL revisamos tres hembras de Acutandra colectadas junto con los machos de A. ubitiara de esa colección; éstas corresponden a la hembra de esta especie y sólo difieren de los machos por la ausencia de la escultura pronotal característica de éstos. Por otro lado, la hembra listada abajo, procedente de Riosucio (Chocó), posee tegumento claramente más oscuro en comparación con las hembras de la Colección CFLP.
Material examinado. COLOMBIA. Cauca. Guapi, Isla de Gorgona, Sendero Yundigua, 40 m, 1 (M), 17.XI.1990, M. Baena (IAvH). Chocó. Riosucio, Caño Seco, 1 (H), 26.III.1999, Carlos Sierra (ICN-MHN-CO 7555, ICN); B. San Juan (Togoromá), 1 (M), colecta diurna en follaje, 22.VII.1990, L. C. Pardo Locarno (CFPL); B. San Juan (Q. Taparal Var.), 1 (M) y 1 (H), colecta diurna en follaje, 21.VII.1990, varios colectores (CFPL); (Taparal), 1 (H), Colecta diurna en follaje, 23.VII.1990, L. C. Pardo Locarno (CFPL); Bajo San Juan (Docordó), 1 (M) y 1 (H), colecta diurna en follaje, 23.VII.1990, L. C. Pardo Locarno (CFPL); Bajo San Juan, HOLOTIPO 1 (M) VII.1990, L. C. Pardo Locarno leg. (CFPL). Valle del Cauca. Bajo Anchicayá, 400 m, 1 (M), II.1985, M. E. Burbano T. (MUSENUV).
Birandra Santos-Silva, 2002
Parandra (Birandra) Santos-Silva, 2002: 47; Monné, 2006: 5 (cat.).
Parandra; Monné, 2006: 15 (cat.); Bousquet, 2008: 620.
Espécie-tipo. Parandra punctata White, 1853, designación original.
Cuerpo corto, poco convexo. Cabeza grande (Figs. 1D y 1E). Cavidades coxales anteriores abiertas posteriormente (Fig. 1F). Mandíbulas grandes y falciformes en los machos major (Fig. 1D), cortas y no falciformes en los machos minor y en las hembras (Fig. 1E). Ápice del labro truncado en los machos (Fig. 1D), agudo en las hembras (Fig. 1E). Área sensorial dorsal del antenómero XI bien delimitada. Élitros puntuados. Paroniquio tarsal con una seta.
Birandra (Yvesandra) Santos-Silva & Shute, 2009
Parandra (Parandra); Monné, 2006: 16 (cat.).
Parandra (Yvesandra) Santos-Silva & Shute, 2009: 24, 31.
Área sensorial del antenómero III-XI dividida por una carena que va desde la base hasta el ápice [a excepción de Birandra (Yvesandra) tavakiliani (Santos-Silva, 2002), donde la carena está presente sólo en el antenómero XI, y es incompleta].
Birandra (Yvesandra) antioquensis (Cardona-Duque et al ., 2007)
Parandra (Parandra) antioquensis Cardona-Duque et al ., 2007: 40.
Birandra (Yvesandra) antioquensis; Santos-Silva & Shute, 2009: 32 (comb. nov.).
Diagnosis. Hembra (Fig. 8A): Cabeza pequeña (mas corta que el pronoto) con puntos gruesos y abundantes (Fig. 8B). Submento glabro, con puntos gruesos más concentrados hacia la región anterior (Fig. 8C). Mandíbulas con dientes anchos en la margen interna. Ojos pequeños, un poco sobresalientes. Antenas un poco más largas que la longitud del protórax. Protórax transversal, margen anterior suavemente sinuoso (Fig. 8B); ángulos anteriores agudos, ángulos posteriores obtusos; pronoto con puntos gruesos y abundantes; márgenes laterales del pronoto curvados hacia arriba. Metaesterno con puntos gruesos y dispersos en las áreas laterales, finos en la región central. Metepisterno con puntos gruesos y dispersos. Élitros con puntos gruesos, abundantes y concentrados.

Distribución. Colombia (Fig. 10C).
Comentarios. Es la primera especie del subgénero Birandra (Yvesandra) registrada para Suramérica, las demás se distribuye en Centroamérica y Las Antillas. Ésta se asemeja principalmente a Birandra pinchoni (Villiers, 1979) pero difiriere de ésta porque en B. pinchoni los ángulos posteriores del pronoto sobresalen en ángulo casi recto, y porque la carena del área sensorial de los antenómeros III-IV es ausente en B. pinchoni.
Material examinado. COLOMBIA. Antioquia. Granada, HOLOTIPO 1 (H), III.1998, M. Londoño col. (CEUA 12428).
Birandra (Birandra) Santos-Silva, 2002
Parandra (Birandra) Santos-Silva, 2002: 47; Monné, 2006: 17 (cat.); Monné & Hovore, 2006: 5 (cat.).
Birandra (Birandra); Santos-Silva & Shute, 2009: 24, 31. Especie-tipo - Parandra punctata White, 1853 (designación original).
Área sensorial del antenómero XI completa (no dividida por carena) (Fig. 8D).
Con excepción de Birandra (Birandra) mariahelenae (Santos-Silva, 2002), todas las especies están presentes en Suramérica.
Clave para las especies de Birandra (Birandra) presentes en Colombia
1. Élitros con puntos finos en los dos sexos; ángulos anteriores del pronoto redondeados; margen anterior del pronoto recta o subrecta (Fig. 9A). Colombia y Venezuela.................. ............................B. (Birandra) lucanoides (Thomson, 1861)
- Élitros con puntos gruesos en los dos sexos; ángulos anteriores del pronoto salientes y aguzados; margen anterior del pronoto suavemente cóncava en los machos, variable en las hembras ................................................................................2
2(1). Protórax fuertemente transversal en los dos sexos (Fig.9J); ápice de las mandíbulas de los machos bífido, no alargado; margen interna de la mandíbula de los machos con un diente grande entre la base y el medio y uno pequeño entre el medio y el ápice (Fig. 9I); margen interna de la mandíbula de las hembras con entrada acentuada cerca al ápice. Colombia, Trinadad y Tobago, Guyana Francesa, Brasil (Amapá), Perú y Bolivia..............................................B. (Birandra) silvaini (Tavakilian, 2000).
- Protórax transversal en los dos sexos (Figs. 9E y 9F); ápice de las mandíbulas de los machos trífido, alargado; margen interna de la mandíbula de los machos con un diente pequeño aproximadamente en el medio (Fig. 9C); margen interna de la mandíbula de las hembras sin entrada marcada cerca al ápice. Colombia, Venezuela, Ecuador y Perú ...............B. (Birandra) punctata (White, 1853).
Birandra (Birandra) lucanoides (Thomson, 1861)
Parandra lucanoides Thomson, 1861b: 85. Localidad-tipo: Colombia. (MNHN).
Parandra (Birandra) lucanoides; Monné, 2006: 17 (cat.); Monné & Hovore, 2006: 5 (cat.).
Birandra (Birandra) lucanoides; Santos-Silva & Shute, 2009: 31 (comb. nov.).
Diagnosis. Cabeza mediana (un poco más corta que el pronoto, castaño oscuro en el sintipo macho), puntos nítidos en el área próxima a los ojos, más dispersos en dirección al centro. Submento con puntos gruesos. Protórax transversal, más estrecho en la base (Fig. 9A). Disco del pronoto con puntuación fina y dispersa; borde anterior recto o subrecto; ángulos anteriores redondeados. Metaesterno glabro, con puntuación fina y dispersa en los lados. Metepisternos glabros, con puntuación similar a aquella del metaesterno. Élitros con puntuación muy fina y dispersa.
Distribución. Colombia y Venezuela.
Comentarios. No examinamos ejemplares de esa especie, entonces basamos la diagnosis en las fotografías de tres sintipos (dos machos y una hembra), la descripción original y las redescripciones.
Rosales (1966) afirmó erróneamente que la especie fue descrita de Venezuela y aunque Thomson (1861b) registró: “Nov. Granata”, la única especie formalmente descrita de Venezuela, fue Parandra gyllenhalii Thomson, 1867, que actualmente está en sinonimia de Parandra (Tavandra) colombica. Hasta 1810 el territorío que hoy corresponde a Colombia comprendía Panamá, Ecuador, Venezuela, parte de Colombia, y la costa atlántica de Costa Rica, Honduras y Nicaragua. Entre 1810 y 1816 recibió el nombre de Nueva Granada y comprendía principalmente el occidente Colombiano, el norte (Sucre, Córdoba y Bolívar) y el oriente (Tolima, Huila, Santanderes, llano y Cundinamarca). Entre 1816 y 1819 se da la reconquista española y aparentemente a partir de 1819 comprendió siempre los territorios de Panamá y Colombia, recibiendo diferentes nombres: Colombia (entre 1819 y 1830), República de la Nueva Granada (entre 1830 y 1858), Confederación Granadina (entre 1858 y 1861), Estados Unidos de la Nueva Granada (entre 1861 y 1863), Estados Unidos de Colombia (entre 1863 y 1886), República de Colombia (entre 1886 y 1903) y finalmente se independiza Panamá en 1903 (Montoya-Guzmán com. pers.). Palacios y Safford (2002) afirman que Venezuela se independizó en 1830, entonces Venezuela no hace parte del territorío como mínimo desde 1830. Dado que no se sabe en qué período se colectaron los sintipos, es imposible afirmar de qué país actual proceden.
Birandra (Birandra) punctata (White, 1853)
Parandra punctata White, 1853: 2. Localidad-tipo: Colombia. (BMNH).
Parandra (Birandra) punctata; Monné, 2006: 17 (cat.); Monné & Hovore, 2006: 5 (cat.).
Parandra luciana Thomson, 1861b: 86. Localidad-tipo: Colombia. (MNHN).
Parandra fabricii Thomson, 1867: 117. Localidad-tipo: Colombia (“Nov. Granata”). (MNHN).
Birandra (Birandra) punctata; Santos-Silva & Shute, 2009: 31 (comb. nov.).
Diagnosis. Cabeza mediana (un poco más corta que el pronoto) (Fig. 9B). Mandíbulas de los machos con el diente del margen interno de la mandíbula muy pequeño (generalmente más pequeño que el diente subapical), localizado aproximadamente en el medio (Fig. 9C); mandíbulas de las hembras con el margen externo redondeado (Fig. 9D). Protórax subcuadrado, más angosto que los élitros, más ancho que largo en la parte anterior (Figs. 9E y 9F); ángulos anteriores agudos y proyectados, ángulos posteriores con puntos gruesos. Élitros brillantes con puntos relativamente gruesos y rasos (Fig. 9G).
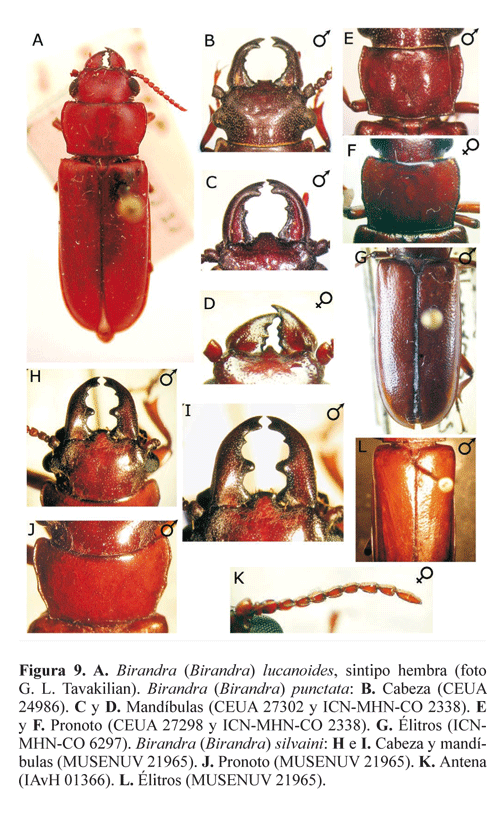
Distribución. Colombia (Fig. 10D), Venezuela, Ecuador y Perú.
Comentarios. En los machos minor el diente subapical de las mandíbulas generalmente está un poco desplazado hacia el centro del margen interno y el diente medio puede ser más grande en relación al tamaño del cuerpo. Por otro lado, las hembras de B. punctata y B. silvaini (Tavakilian, 2000) son muy semejantes, de difícil separación y aunque las hembras de B. punctata son proporcionalmente más delgadas, esa diferencia sólo puede ser bien interpretada cuando se examinan ejemplares de las dos especies. Con 66 especímenes en las colecciones colombianas, fue la segunda especie más abundante; se encuentra en las tres cordilleras a alturas superiores a los 1000 m.
Material examinado. COLOMBIA. Antioquia. Medellín, Corregimiento San Antonio de Prado, Vereda El Astillero, 2400 m, 6 (M), manual en tronco en descomposición, 18.V.2002, M. Castaño (CEUA - 27297, 27298, 27299, 27300, 27301, 27302); Medellín, Corregimiento San Antonio de Prado, Vereda Yarumalito, 2 (M), manual, 1-3.XI.2008, C. Bota y N. Uribe (CEUA - 44567, 44568); Medellín, San Antonio de Prado, 1 (M), en suelo, I.1984, C. U. G. (No. 8773, MFLG); Caldas, Vereda La Clara, Alto de San Miguel, 1 (M), manual, suelo, VI.1996, Grupo de Entomología (CEUA - 40585); Caldas, Vereda La Clara, Alto de San Miguel, 2100 m, 1 (M), manual en vuelo, sin fecha de colección, William Ramírez (CEUA - 24985); Caldas, 1 (M), en M.O., X.1984, Andrés García (No. 8772, MFLG); Caldas, 1 (M), en tronco, IX.1989, A. Zapata (No. 8772, MFLG); Caldas, Reposo, 1 (M), 19.VIII.1989, Saldarriaga (No. 8772, MFLG); Caldas, 1 (H), en piso, IX.1989, Carlos Henao (No. 8772, MFLG); Cocorná, 1 (M), en tronco viejo, IX.1989, A. Zapata (No. 8772, MFLG); Gómez Plata, Hacienda Normandia, 1000 m, 1 (M), bosque, interior de bosque, 16.X.2000, J. D. Guzmán (No. 8772, MFLG); Urrao, 1 (M), en líquenes, 3.V.1972, G. Lozano (ICN-MHN-CO 4639, ICN); Amalfi, Vereda Guayabito, Finca Costa Rica, Finca Caracolí, Quebrada La Hueso, 1840 m, 1 (M) y 7 (H), manual debajo de tronco, 19.II.2008, Vélez, Díaz y Rivera (CEUA - 44569, 44570, 44571, 44572, 44573, 44574, 44575, 44576). Boyacá. Pajarito, Cosinto, 1600 m, 1 (H), 27.X.1979, C. Hernández (ICN-MHN-CO 2338, ICN). Caldas. Samaná, 1 (M), manual en árbol, VIII.1994, R. García E. (CEUA - 24986); Manizales, 2200 m, 1 (M), en suelo, 24.II.2006, Aguirre - Benitez (LEUC); Manizales, Reserva Rio Blanco, 2650 m, 1 (M), 8.X.2005, C. Santamaria (MUSENUV); Manizales, Vereda Las Palomas, Reserva Río Blanco, 5°7,2´58”N; 75°43,7´58”W, 2592-2690 m, 1 (M), Trampa de luz negra, 3.IV.2005, A. Vélez (CEUA - 38252). Cauca. Estación Termal, INDERENA, 2800 m, 1 (M), 29.X.1980, I. de Arévalo (ICN-MHN-CO 4623, ICN); Inzá, Vereda Tierras Blancas, Km 28, 2630 m, 1 (M), 25.III.1982, Coler. Gnipo (ICN-MHN-CO 2336, ICN); El Tambo, PNN Munchique, La Gallera, Km 75, 1800 m, 4 (M) y 1 (H), 10.VI.1990 y 15.IX.1990, A. J. Negret (EN 000231 R, 000232 R, 000230 R, 000228 R, 000229 R, MHN-UC). Cundinamarca. Quétame, 1 (M), en maleza, XI.1946, F. L. Gallego (No. 8773, MFLG); La Mesa, Laguna Pedro-Palo, 4°38´05”N; 74°27´57”W, 1 (H), 16.XI.1991, Barbosa y Garcés (FAUNIC 10, UNAB). Huila. Algeciras, El Paraíso, Termópilos, 2200 m, 3 (M), IX.1998 y II.1999, D. Trujillo (ICN-MHN-CO 6297, 7560, 7565, ICN); Meremberg, 2400 m, 1 (M), X.1981 y 2.X.1981, Jorge García e Inge Armbrecht (MUSENUV - 6407, 6403); Gigante, 1 (M), directa, 25.XI.1993, Héctor Bonilla (CFPL). Nariño. Ricaute, La Planada, 1 (M), bosque, VII.1994, F. J. Serna (No. 8772, MFLG); La Planada, 1850 m, 1 (M), tronco, 20.III.1993, G. Celorío (MUSENUV - 21967). Risaralda. Santuario, Vereda Los Planes, Finca Las Delicias, 2200 m, 1 (M), XI.1991, F. Fernández (ICN-MHN-CO - 4074, ICN); Pueblo Rico, 1200 m, 1 (M), VII.1994, G. Andrade (ICNMHN- CO 2340, ICN); Ucumarí, La Suiza, 1900 m, 2 (M) y 4 (H), 19-21.VIII.1992, Germán Amat, Matuk-Ochoa y Lorma (MUJ - 5908, 5912, 5914, 5916, 5905, 5909); Ucumarí, La Suiza, 1900 m, 1 (M) y 1 (H), 21-22.III.1992, Amat G. y Durán y Sierra (MUJ - 5910, 5911). Tolima. Espinal, 1 (H), 16.IV.1986, D. N. Padilla (ICN-MHN-CO 2335, ICN). Valle del Cauca. Dagua, Queremal, Antena T.V., 2030 m, 1 (M), 1978, sin colector (MUSENUV - 6404); Palmira, La Buitrera, 1500 m, 1 (M), domicilio, 24.III.1994, F. Montealegre (MUSENUV - 6402); Peñas Blancas, 1500 m, 1 (M) y 1 (H), II.1986, G. Bermeo (MUSENUV - 21968); Corea, 2500 m, 1 (M), II.1984, CCR (MUSENUV); M. Calima, Q. Cristalina Alta, 1 (M), Colecta diurna en follaje, 21-28.VII.1992, varios colectores (CFPL).
Birandra (Birandra) silvaini (Tavakilian, 2000)
Parandra silvaini Tavakilian, 2000: 150. Localidad-tipo: Guyana Francesa, Matoury. (MNHN).
Parandra (Birandra) silvaini; Monné, 2006: 18 (cat.).
Birandra (Birandra) silvaini; Santos-Silva & Shute, 2009: 31 (comb. nov.).
Diagnosis. Cabeza grande (tan larga como el pronoto en especímenes de tamaño promedio) (Fig. 9H), puntuación más fuerte que en los élitros e incluso que en el tórax. Mandíbulas de los machos con el diente interno grande (generalmente más grande que el diente subapical), localizado en el tercio basal (Fig. 9I). Antenómeros III-XI con una sola foseta sensorial (Fig. 9K). Protórax transversal, ligeramente emarginado en la parte posterior (Fig. 9J); ángulos anteriores agudos y proyectados; ángulos posteriores agudos, proyectados, moderadamente redondeados; margen anterior del pronoto cóncava. Élitros con puntos relativamente gruesos, concentrados y rasos (Fig. 9L).
Distribución. Bolivia, Brasil, Colombia (Fig. 10C), Guyana Francesa, Perú y Trinidad y Tobago.
Comentarios. De los ejemplares seleccionados por Tavakilian (2000) como paratipos, siete tenían etiquetas que indicaban que procedían de la colección de Thomson, de los cuales un macho portaba la siguiente etiqueta: “Parandra punctata White Vol. 2 Nouvelle-Grenade” (Tavakilian 2000). Sin embargo Tavakilian (2000) anotó: “Je ne connais que des spécimens de Guyane, mis à part l´exemplaire de la collection Thomson qui porte una étiquette bleue indiquant comme provenance la Nouvelle-Grenade (Colombie). Très certainement, cette indicatión fausse est due à una mauvaise interprétation de l´espèce de White et au fait que le matériel acquis par Thomson était dépourvu de mentión de localité”; así, afirmó que los especímenes de B. punctata de la colección de Thomson correspondían a B. silvaini y dudó que hubiesen sido colectados en Colombia (Tavakilian 2000). En consecuencia Monné y Hovore (2006) y Monné (2006) registraron esta especie sólo en la Guyana Francesa, aún cuando Santos- Silva (2002a) ya había registrado B. silvaini para Colombia, Trinidad y Tobago, Guyana Francesa, Brasil, Perú y Bolivia. En las colecciones colombianas se encontraron dos especímenes: una hembra colectada en un Parque Nacional Natural de la Cordillera Oriental, probablemente en una zona boscosa y un macho colectado en una hidroeléctrica en el Valle del Cauca.
Por otro lado, Lameere (1902) sin consultar la colección de Thomson, sinonimizó a Parandra punctata White, 1853 sensu Thomson con Parandra punctatissima Thomson, 1861, sensu Lameere. Posteriormente Tavakilian (2000) sinonimizó ambos conceptos con Birandra silvaini.
Birandra silvaini difiere de B. punctata porque el diente del margen interno de la mandíbula de los machos es muy desarrollado, y porque la relación largo-ancho del cuerpo en ambos sexos es mayor en B. silvaini. Adicionalmente las fosetas antenales son más grandes en B. silvaini y las antenas son un poco más glabras (Tavakilian 2000) (Figs. 8D y 8E).
Material examinado. COLOMBIA. Meta. Macarena, PNN Tinigua, Pto. Chamuza, Rio Duda, 2°40´N; 74°10´W, 350 m, 1 (H), 1994, M. Alvarez y A. Calixto (IAvH - 01366). Valle del Cauca. Central Hidroeléctrica Anchicayá, 400 m, 1 (M), XI.1970, sin colector (MUSENUV - 21965).
Registro dudoso para la fauna de Colombia
Parandra (Tavandra) longicollis Thomson, 1861
Parandra longicollis Thomson, 1861b: 83. Localidad-tipo: Brasil. (MNHN).
Hesperandra (Tavandra) longicollis; Monné, 2006: 11 (cat.).
Parandra gracillima Bates, 1869: 38. Localidad-tipo: Brasil, Amazonas: Ega [Tefé]. (MNHN).
Parandra (Tavandra) longicollis; Santos-Silva & Shute, 2009: 31 (comb. nov.).
Distribución. México hasta Argentina.
Comentarios. Arigony (1978) registró Parandra longicollis para los departamentos de Antioquia, Caldas, Cudinamarca y Tolima. Además anotó que se distribuye desde la Mata Atlántica, pasando por las fajas de transición y el Cerrado y alcanzando la Hiléia (límite occidental del estado de Amazonas con los países andinos) hasta las regiones boscosas de las vertientes orientales de los Andes, los bosques pluviales del Pacífico y el Caribe, las zonas altas de los Andes y los valles secos interandinos. Además mencionó que se presentaba en México y Estados Unidos (Arigony 1978).
Chemsak (1996) excluyó esa especie de la fauna de Estados Unidos anotando que aunque Arigony (1978) la registró para Texas con base en dos especímenes del U.S. Museum of Natural History, P. longicollis no se ha encontrado en esta área a pesar de grandes esfuerzos de muestreo.
Es muy probable que Parandra (Tavandra) longicollis esté restringida al oriente brasilero y no esté en la mayoría de las regiones señaladas por Arigony (1978), principalmente en México y Centroamérica. El examen de algunos especímenes de regiones amazónicas sugiere que los ejemplares identificados para esta zona por Arigony (1978) como P. longicollis, corresponden a la forma de Parandra gracillima Bates, 1869. Esta forma concuerda con Parandra lalannecassouorum (Tavakilian, 2000), y por esta razón P. gracillima debería ser revalidada y P. lalannecassouorum sinonimizada bajo ésta. Sin embargo, es necesarío examinar una gran cantidad de especímenes y los tipos para confirmarlo.
Especies excluidas de la fauna de Colombia
Acutandra punctatissima (Thomson, 1861)
Parandra punctatissima Thomson, 1861b: 84. Localidadtipo: Guyana Francesa. (MNHN).
Distribuición: Guyana Francesa
Acutandra punctatissima; Monné, 2006: 9 (cat.).
La distribución conocida de A. punctatissima (Thomson, 1861) es Guyana Francesa y Colombia. Santos-Silva (2000a) registró esta especie para Colombia citando como material examinado lo siguiente: “COLÔMBIA, Valle del Cauca: Bajo Calima, fêmea, sem data, L. C. Pardo Locardo col. (MZSP)”. El ejemplar fue donado al MZSP por Luís Carlos Pardo y procedía de su colección. Al examinar todos los ejemplares de Parandrinae depositados en CFPL, encontramos varios machos de Acutandra ubitiara y varias hembras que se identificaron como A. ubitiara, que corresponden a la hembra fotografiada por Santos-Silva (2002: fig. 52). Adicionalmente se examinó otra hembra en ICN del norte del departamento de Chocó, que tampoco corresponde a A. punctatissima. Todos estos especímenes presentan los ángulos posteriores del pronoto obtusos, como no ocurre en A. punctatissima. Thomson (1861b) en su descripción de A. punctatissima indicó: “...angulis lateralibus anticis et positcis productis,...”; igualmente, Tavakilian (2000) al redescribir la especie señaló en relación a estos ángulos: “il est rétréci vers l´arrière, avec les angles postérieurs vifs”. Con base en esto excluimos formalmente a Acutandra punctatissima de la fauna de Colombia.
Parandra (Parandra) ubirajarai (Santos-Silva, 2001) / Acutandra degeerii (Thomson, 1867)
Hesperandra ubirajarai Santos-Silva, 2001b: 216.
Hesperandra (Zikandra) ubirajarai; Monné, 2006: 15 (cat.); Santos-Silva, 2007: 8, 9 (clave).
Parandra (Parandra) ubirajarai; Santos-Silva & Shute, 2009: 31 (comb. nov.).
Parandra degeerii Thomson, 1867: 111.
Acutandra degeeri; Monné, 2006: 8 (cat.).
Parandra (Parandra) ubirajarai (Santos-Silva, 2001) fue registrada en Colombia por Arigony (1978) y Martínez (2000) como «Parandra (Hesperandra) degeerii Thomson, 1867». Igualmente fue citada en la lista de chequeo de Monné y Giesbert (1994). Adicionalmente Monné y Hovore (2006) mencionaron que Acutandra degeerii (Thomson, 1867) se distribuía en “México?, Panama-c SAmer”. Sin embargo, los registros fuera de Brasil son exclusivamente de Arigony (1978) que en realidad estaba hablando de P. (P.) ubirajarai, confundida por ella y autores anteriores con A. degeerii (= Parandra degeerii). La descripción de A. degeerii sólo señala como localidad Brasil; de esta especie solamente se conocen los sintipos (un macho y una hembra), depositados en el MNHN cuya etiqueta únicamente decía “Brasil” (Gérard Tavakilian com. pers.).
Arigony (1978) registró Acutandra degeeri en Colombia (Manizales, departamento de Caldas), a partir del examen de dos machos provenientes del MNHN, afirmando: “P. degeeri é simpátrica com P. glabra na região da mata atlántica e na costa do Pacífico desde Loja, Equador, até Medellín, Colômbia, na zona equatorial dos Andes rumo ao interior do continente até Tiquié, Amazonas, Brasil.”. También afirmó que la marginación lateral del pronoto es completa en P. (P.) ubirajarai [mencionada como Parandra (Hesperandra) degeeri], lo que es un error. Los especímenes identificados por Arigony como ‘Parandra degeerii´ pertencientes al MZSP, exhiben la marginación lateral del protórax incompleta. Además las figuras y la descripción presentadas por Arigony (1978), y la inclusión en el entonces subgénero P. (Hesperandra), no dejan dudas de que la autora se estaba refiriendo a P. ubirajarai y no a Acutandra degeerii.
Consecuentemente la citación de Monné y Hovore (2006) de Acutandra degeerii para México, Centro América y áreas de Sur América es errónea. La distribución conocida de A. degeerii es Brasil (sin localidad específica) y P. ubirajarai se distribuye en el oriente de Brasil (Monné 2006). De esta forma excluimos formalmente las dos especies de la fauna de Colombia.
Consideraciones Finales
Varias de las especies de Parandrinae presentan una variación relativamente grande a lo largo de su ámbito y los caracteres que delimitan estas especies muchas veces no consiguen abarcar la variación. Por esto se requiere explorar otros caracteres que permitan separar las especies de Parandrinae y aunque algunos autores han propuesto que los órganos genitales son una fuente confiable de caracteres para la separación de especies (e.g. Arnqvist 1998; Córdoba-Aguilar 2000), en los Parandrinae no han sido arduamente explorados como recurso taxonómico. Los pocos trabajos que han considerado estos caracteres, no han realizado análisis suficientemente exhaustivos como para llegar a conclusiones definitivas. Así por ejemplo, Arigony (1977, 1978) afirmó que no existían características muy divergentes en los órganos genitales de los Parandrinae; sin embargo, en sus ilustraciones publicadas se evidencia una variación interespecífica que tal vez permitiría separar las especies.
Por otro lado, algunos autores indicaron que la ausencia de pelos o puntos en las antenas es una característica de Parandrinae (e.g. Linsley 1962; Villiers 1980; Napp 1994; Chemsak 1996). Sin embargo, algunas especies de parandrinos poseen puntuación en las antenas (que puede ser gruesa en el escapo) y la mayoría presentan pilosidad en los antenómeros, la cual es variable pero siempre visible; ésta ha sido utilizada como una herramienta taxonómica que puede ser útil en la separación de especies similares (e.g. Tavakilian 2000).
Finalmente, las especies de Parandrinae son raramente colectadas, por lo que los datos de historia natural son extremadamente pobres y se desconocen los estadíos inmaduros de la gran mayoría de las especies. En Colombia la escasez de registros en regiones como la Orinoquía y Amazonía, son el reflejo de la falta de muestreo. Al igual que para otros grupos, las localidades con algún registro están concentradas principalmente en las regiones cercanas a los grandes centros urbanos, donde existen universidades y centros de investigación.
Agradecimientos
El primer autor agradece al Dr. Ricardo Callejas, Dr. Frank T. Hovore (QEPD), Andrés H. Vélez, Carolina Rivera y María C. Vélez, por su apoyo durante el desarrollo del proyecto de Parandrinae de Colombia. A Natalia Carrillo, Javier Martínez, Dr. Julio Betancur, Dr. Eurípedes Menezes, Claudia Hussak y Gustavo Mesa (QEPD), por su hospitalidad durante el tiempo de visita a las colecciones. A Sandra Enriques por compartir su literatura. A la Dra. Dilma Solange Napp, al Dr. Miguel A. Monné y al Dr. Steven Lingafelter por facilitarme sus trabajos en Parandrinae. A Camilo Flórez por sus ilustraciones y a Juan David Montoya Guzmán por la información sobre la historia de Colombia. A los directores y curadores de todas las colecciones visitadas, por facilitarnos la revisión de las mismas. A Álvaro Idárraga por su ayuda con la información de los hospederos y a Carmen Elisa Posso por su ayuda con algunas localidades del Valle del Cauca. A Claudia Martínez por el préstamo de parte del material de ICN. Al Dr. Daniel R. Miranda-Esquivel y María C. Vélez por aclarar algunas dudas con algunos de los trabajos en sistemática de Cerambycidae. A Cornelio A. Bota-Sierra y Jennifer C. Girón por revisar el manuscrito. Finalmente a GEUA por la financiación parcial del proyecto.
Literatura citada
ARIGONY, T. H. A. 1970. Notas sôbre Parandrinae (Col., Cerambycidae) II. Redescrição de Parandra villei Lameere, 1885. Revista Brasileira de Entomología 14 (8): 57-62. [ Links ]
ARIGONY, T. H. A. 1977. Notas sobre Parandrinae (Coleoptera, Cerambycidae) V. Redescrição de Parandra (Archandra) caspia Ménétriès, 1832. Revista Brasileira de Entomología 20 (3/4): 159-164. [ Links ]
ARIGONY, T. H. A. 1978. Revisão do gênero Parandra (Coleoptera, Cerambycidae). 1. O Subgênero Hesperandra Arigony, 1977. Revista Brasileira de Entomología 22 (3/4): 119-159. [ Links ]
ARIGONY, T. H. A. 1984. O subgênero Parandra Latreille, 1804 (Coleoptera, Cerambycidae, Parandrinae): Estudo fenético e cladístico de 12 espécies. Iheringia, Série Zoologia 64: 87-125. [ Links ]
ARNQVIST, G. 1998. Comparative evidence for the evolution of genitalia by sexual selection. Nature 393: 784-786. [ Links ]
BATES, H. W. 1869. IX. Contributions to an Insect Fauna of the Amazon Valley (Coleoptera, Prionides). The Transactions of the Entomological Society of London 1869: 37-58. [ Links ]
BATES, H. W. 1872. On the longicorn Coleoptera of Chontales, Nicaragua. The Transactions of the Entomological Society of London 1872: 163-238. [ Links ]
BATES, H. W. 1879. Coleoptera. pp 1-16. En: Goldman, F. D. C.; Salvin, O. (eds.). Biología Centrali-Americana, Insecta. London, British Museum (Natural History) Vol V: 526p + 26 planchas. [ Links ]
BLACKBURN, B. A. 1895. Further notes on Australian Coleoptera, with descriptions of new genera and species. Transactions of the Royal Society of South Australia 19: 27-60. [ Links ]
BLACKWELDER, R. E. 1946. Checklist of the coleopterous insects of México, Central America, the West Indies and South America. Part 4. Bulletin of the United States National Museum 185: 551-763. [ Links ]
BLANCHARD, C. E. 1845. Histoire des Insectes, traitant de leurs moeurs et de leurs métamorphoses en général, et comprenant une nouvelle classificatión fondée sur leurs rapports naturels. Paris, Didot 2: 1-524. [ Links ]
BOUSQUET, Y. 2008. Nomenclatural and bibliographic notes on Cerambycidae (Coleoptera). The Coleopterists Bulletin 61 (4): 616-631. [ Links ]
BÖVING, A. G.; CRAIGHEAD, F. C. 1931. An illustrated synopsis of the principal larval forms of the order Coleoptera. Entomologica Americana 9: 1-35. [ Links ]
CARDONA-DUQUE, J.; SANTOS-SILVA, A.; WOLFF, M. 2007. A new species of Parandrinae from the Central Andes of Colombia (Coleoptera: Cerambycidae). Zootaxa 1661: 39-45. [ Links ]
CASEY, T. L. 1912. III - Studies in the Longicornia of North América. Memoirs on the Coleoptera 3: 215-376. [ Links ]
CHALUMEAU, F.; TOUROULT, J. 2005. Les longicornes des petites Antilles (Coleoptera, Cerambycidae) Taxonomie, éthologie, biogéographie. Sofia, Pensoft. 241 p. [ Links ]
CHEMSAK, J. A. 1996. Illustrated revisión of the Cerambycidae of North America. I. Parandrinae, Spondylidinae, Aseminae, Prioninae. Burbank, Wolfsgarden. x + 149 p. [ Links ]
CÓRDOBA-AGUILAR, A. 2000. Evolución y diversidad de la morfología de los genitales masculinos en insectos. Folia Entomológica Mexicana 110: 95-111. [ Links ]
COSTA, C.; VANIN, S. A.; CASAI-CHEN, S. A. 1988. Larvas de Coleoptera do Brasil. São Paulo, Museu de Zoologia. 282 p. + 165 est. [ Links ]
COWAN, C. F. 1971. On Guérin´s Iconographie: particularly the insects. Journal of the Society for the bibliography of Natural History 6 (1): 18-29. [ Links ]
CRAIGHEAD, F. C. 1915. Contributions toward a classificatión and biology of the north american Cerambycidae. Larvae of the Prioninae.USDA Official Secretary Report 107: 1-24. [ Links ]
CROWSON, R. A. 1955. The natural classification of the families of Coleoptera. London, Nathaniel Lloyd. 187 p. [ Links ]
CROWSON, R. A. 1960. The phylogeny of Coleoptera. Annual Review of Entomology 5: 111-134. [ Links ]
DAMOISEAU R.; COOLS, J. 1987. Liste du materiel typique dans les collections entomologiques de l'Institut royal des Sciences naturelles de Belgique. Coleoptera Cerambycoidea Cerambycidae: Aseminae, Cerambycinae, Disteniinae, Lepturinae, Parandrinae, Prioninae et Spondylinae. Documents de Travail 42: 1-39. [ Links ]
DEGEER, C. 1774. Memoires pour servir a l'histoire des insectes. Stockholm, Imp. Pierre Hesselberg. 4: xii + 456 p. [ Links ]
DIAS, M. M. 1981. Analise cladistica e classificacao de Anoplodermatinae (Coleoptera, Cerambycidae). Tese de doutoramento, Instituto de Biociencias, Universidade de Sao Paulo. v + 144 p. [ Links ]
FABRICIUS, J. C. 1794. Entomología systematica emendata et aucta. Secundum Classes, Ordines, Genera, Species adjectis synonymis, locis, observationibus, descriptionibus. Hafniae. C. G. Proft & fils 4: 1-472. [ Links ]
FAIRMAIRE, L. 1881. Essai sur les Coleopteres des iles Viti (Fidgi). Annales de la Societe Entomologique de France (6) 1: 461- 492. [ Links ]
FALLING RAIN GENOMICS, INC. 1996-2006. Global gazetteer versión 2.1. Calle.com. Disponible en: http://www.fallingrain.com/world/ fecha de revision: abril 2007 [ Links ]
FARRELL, B. D.; SEQUEIRA, A. S. 2004. Evolutionary rates in the adaptive radiatión of beetles on plants. Evolutión 58 (9): 1984-2001. [ Links ]
FONSECA-GESSNER, A. A. 1990. Revisao taxonomica do genero Paramallocera Aurivillius, 1912 (Coleoptera, Cerambycidae, Cerambycinae, Sphaerionini). Revista Brasileira de Entomología 34 (4): 817-856. [ Links ]
FORBES, W. T. M. 1922. The wing-venation of the Coleoptera. Annals of the Entomological Society of America 15 (4): 327-352. [ Links ]
GAHAN, C. J. 1895. On the Longicorn Coleoptera of the West Indian Islands. The Transactions of the Entomological Society of London 1895: 79-140. [ Links ]
GAHAN, C. J. 1906. The Fauna of British India including Ceylon and Burma. Vol. I (Cerambycidae). Londres, C. T. Bingham xviii + 329 p. [ Links ]
GEMMINGER, M.; HAROLD, E. 1872. Catalogus Coleopterorum hucusque descriptorum synonymicus et systematicus. Monachii 9: 2669-2988. [ Links ]
GERMAR, E. F. 1846. En: Herausgeber, J. S. Ersch & J. G. Gruber. Allgemeine Encyclopadie der Wissenschaften und Kunste in alphabetischer Folge von genannten Schriftstellern bearbeitet und herausgegeben. Vol. 25: 513 p. [ Links ]
GIRARD, M. J. A. 1873. Traite elementaire d´Entomologie. Paris, J. Baillere & fils. viii + 840 p. [ Links ]
GORY, H. L. 1831. Iconographie du Regne Animal de Georges Cuvier ou representatión d'apres nature de l'une des especes le plus remarquables et souvent non figurees de chaque genre d'animaux. En: F. E. Guerin-Meneville. Insectes. Cerambycidae. Paris 7: pls 42-44. [ Links ]
GORY, H. L. 1844. Iconographie du Regne Animal de Georges Cuvier ou representatión d'apres nature de l'une des especes le plus remarquables et souvent non figurees de chaque genre d'animaux. En: Guerin-Meneville, F. E. Insectes. Cerambycidae. Paris. 7: iv + 5-576, 110 pls. [ Links ]
HORN, G. H. 1875. Synonymical notes and descriptions of new species of North American Coleoptera. Transactions of the American Entomological Society, Philadelphia 5: 126-156, 8 figs. [ Links ]
HORN, W.; KAHLE, I. 1935-1937. Uber entomologische Sammlungen, Entomologen & Entomo-Musologie. Entomologische Beihefte aus Berlin-Dahlem 2/4: I + 536 p. [ Links ]
IGAC. 1996. Diccionario Geográfico de Colombia. Instituto Geográfico Agustin Codazi. Multimedia. [ Links ]
IGAC. 2002. Atlas de Colombia. Quinta Edicion. Bogota, Imprenta Nacional de Colombia, 342 p. [ Links ]
INTERNATIONAL COMMISSION ON ZOOLOGICAL NOMENCLATURE (ICZN). 1964. International Code of Zoológical Nomenclature adopted by the XV Congress of Zoology. The International Trust for Zoológical Nomenclature, London, United Kingdon. 176 p. [ Links ]
INTERNATIONAL COMMISSION ON ZOOLOGICAL NOMENCLATURE (ICZN). 1999. International Code of Zoológical Nomenclature adopted by the International Unión of Biological Sciences. The International Trust for Zoológical Nomenclature, London, United Kingdon. 306 p. [ Links ]
JENI., I. 2001. Tesa.ici / Long-horned beetles. Distenidae, Oxypeltidae, Vesperidae, Anoplodermatidae & Cerambycidae I. Vesperidae & Cerambycidae Evropy / of Europe I. Zlin, Atelier Regulus, 333 p. [ Links ]
KIRBY, W. 1837. The Insects. Coleoptera. En: Richardson J. Fauna Boreali Americana; or the zoology of the northern parts of British America. Norwich, Josiah Fletcher, 4: xxxix + 335 p. [ Links ]
KIRSCH, T. 1889. Coleoptera gesammelt in den Jahren 1868-1877 auf einer Reise durch Sud Amerika von Alphons Stubel. Abhandlungen und Berichte des koniglichen zoologischen und anthropologisch- ethnographischen Museums zu Dresden 1888/1889 (4): 1-58. [ Links ]
KOLBE, H. J. 1893. Beitrage zur Kenntniss der Longicornier (Coleoptera). Stettiner Entomologische Zeitung 54: 241-290. [ Links ]
LAMEERE, A. A. 1885a. Notes sur quelques Prionidae. Comptes- Rendus des Seances de la Societe Entomologique de Belgique 29: ix-xiii. [ Links ]
LAMEERE, A. A. 1885b. Contributión a l'histoire des metamorphoses des longicornes de la famille des Prionidae. Memoires de la Societe Royale des Sciences de Liege (2)11 (11): 1-13. [ Links ]
LAMEERE, A. A. 1901. Etude sur la Phylogenie des Longicornes. Premiere communication preliminaire. Annales de la Societe Entomologique de Belgique 45: 314-323. [ Links ]
LAMEERE, A. A. 1902. Revision des prionides (Premier memoire - Parandrides). Annales de la Societe Entomologique de Belgique 46: 59-111. [ Links ]
LAMEERE, A. A. 1912. Revision des Prionides. Addenda et corrigenda. Memoires de la Societe Entomologique de Belgique 21: 113-188. [ Links ]
LAMEERE, A. A. 1913. Coleopterorum Catalogus, pars 52, Cerambycidae. Prioninae. Berlin, W. Junk. 108 p. [ Links ]
LAMEERE, A. A. 1919. Genera Insectorum. Coleoptera, Fam. Cerambycidae, subfam. Prioninae. Bruxelles, P. Wytsman. v. 172, 189 p. [ Links ]
LANSBERG, J. W. 1884. Catalogue des Prionides de l'Archipel Indo- Neerlandais, avec descriptions des especes nouvelles. Notes from the Leyden Museum 6: 135-160. [ Links ]
LAPORTE, F. L. N. 1840. Histoire Naturelle des Insectes Coleopteres. Paris, P. Dumenil 2: 1-563. [ Links ]
LATREILLE, P. A. 1802. Histoire Naturelle, generale et particuliere des Crustaces et des Insectes. Imprimerie F. Dufart, Paris, 3: i-xii, 1-468. [ Links ]
LATREILLE, P. A. 1804. Histoire naturelle, generale et particuliere des Crustaces et des Insectes. Paris, F. Dufart, 11: iv + 1-424. [ Links ]
LATREILLE, P. A. 1807. Genera Crustaceorum et Insectorum secundum ordinem naturalem in familias disposita, iconibus exemplisque plurimis explicata. Paris & Argentorat, Armand Koenig, 3: 1-258. [ Links ]
LATREILLE, P. A. 1810. Considerations generales sur l'ordre naturel des animaux composant les classes des crustaces, des arachnides, et des insectes ; avec un tableau methodique de leurs genres, disposes en familles. Paris. 444 p. [ Links ]
LATREILLE. 1817. En : Cuvier. Le Regne Animal distribue d'apres son organisation, pour servir de base a l'Histoire Naturelle des animaux, et d'introductión a l´Anatomie comparee. Crustaces, Arachnides et Insectes. Paris. Deterville. Vol. 3. 653 p. + 2 est. [ Links ]
LAWRENCE, J. F. 1991. Cerambycidae (Chrysomeloidea) (Including Disteniidae, Hypocephalidae, Oxypeltidae, Parandridae, Spondylidae, Vesperiidae). p. 556-561. En: Stehr, F. W. 1991. Immature Insects. Volume 2. Kendall/Hunt Publishing Company, Iowa. 975 p. [ Links ]
LAWRENCE, J. F.; BRITTON E. B. 1994. Australian Beetles. Carlton, Melburne University Press. 192 p. [ Links ]
LECONTE, J. L.; HORN, G. H. 1883. Classificatión of the Coleoptera of North America. Prepared for the Smithsonian Institution. Smithsonian Miscellaneous Collections 26 (507): i-xxvii + 1-567. [ Links ]
LINGAFELTER, S. W. 1998. Observations of interactive behavior in Parandra glabra (Coleoptera: Cerambycidae). Entomological News 109 (1): 75-80. [ Links ]
LINGAFELTER, S. W.; MICHELI, C. J. 2004. New species of Ceramrycidae (Coleoptera) from Puerto Rico with records and notes for other species. Journal of New York Entomological Society 112 (1): 37-55. [ Links ]
LINSLEY, E. G. 1942. A review of the fossil Cerambycidae of North America. Proceedings of the New England Zoológical Club 21: 17-42. [ Links ]
LINSLEY, E. G. 1959. Ecology of Cerambycidae. Annual Review of Entomology 4: 99-138. [ Links ]
LINSLEY, E. G. 1961. The Cerambycidae of North America. Part I. Introduction. University of California Publications in Entomology 18: 1-97. [ Links ]
LINSLEY, E. G. 1962. The Cerambycidae of North America. Part II. Taxonomy and classificatión of the Parandrinae, Prioninae, Spondylinae, and Aseminae. University of California Publications in Entomology 19:1-103. [ Links ]
MARTÍNEZ, C. 2000. Escarabajos Longicornios (Coleoptera: Cerambycidae) de Colombia. Biota Colombiana 1 (1): 76-105. [ Links ]
MARTYNOV, A. V. 1926. On the identificatión of fossil insects from the Jurassic shales of Turkestan. 5. Some forms of beetles. Yearbook of the Russian Palaeontological Society 5: 1-38. [ Links ]
MARVALDI, A. E.; DUCKETT, C. N.; KJER, K. M.; GILLESPIE, J. J. 2009. Structural alignment of 18S and 28S rDNA sequences provides insights into phylogeny of Phytophaga (Coleoptera: Curculionoidea and Chrysomeloidea). Zoológica Scripta 38 (1): 63-77. [ Links ]
MONNÉ, M. A. 2002. Catalogue of the Neotropical Cerambycidae (Coleoptera) with known host plant - Part V: Subfamilies Prioninae, Parandrinae, Oxypeltinae, Anoplodermatinae, Aseminae and Lepturinae. Publicações Avulsas do Museu Nacional 96: 1-72. [ Links ]
MONNÉ, M. A. 2004. Catalogue of the Neotropical Cerambycidae (Coleoptera) with known host plant - Part VI: host plant index. Publicações Avulsas do Museu Nacional 100: 1-95. [ Links ]
MONNÉ, M. A. 2006. Catalogue of the Cerambycidae (Coleoptera) of the Neotropical Region. Part III. Subfamilies Parandrinae, Prioninae, Anoplodermatinae, Aseminae, Spondylidinae, Lepturinae, Oxypeltinae, and addenda to the Cerambycinae and Lamiinae. Zootaxa 1212: 1-244. [ Links ]
MONNÉ, M. A.; GIESBERT, E. F. 1994. Checklist of the Cerambycidae and Disteniidae (Coleoptera) of the Western Hemisphere. Burbank, Wolfsgarden Books. 410 p. [ Links ]
MONNÉ, M. A.; HOVORE, F. T. 2006. Checklist of the Cerambycidae, or longhorned wood-boring beetles, of the Western Hemisphere. Rancho Dominguez, Bio Quip Publications. 394 p. [ Links ]
NAKAMURA, S. 1981. Morphological and taxonomic studies of the Cerambycid pupae of Japan (Coleoptera: Cerambycidae). Miscellaneous Reports of the Hiwa Museum for Natural History 20: 1-159. [ Links ]
NAPP, D. S. 1994. Phylogenetic relationships among the subfamilies of Cerambycidae (Coleoptera - Chrysomeloidea). Revista Brasileira de Entomología 38 (2): 265-419. [ Links ]
OLIVIER, A. G. 1803. Nouveau dictionnaire d´histoire naturelle, appliquée aux arts, principalement à l´agriculture et à l´économie rurale et domestique: par une société de naturalistes et d´agriculteurs: avec des figures tirées des trois règnes de la nature. Tome XVII. Deterville, Paris. 574 p. [ Links ]
PALACIOS, M.; SAFFORD, F. 2002. Colombia: país fragmentado, sociedad dividida. Editorial Norma, Bogotá. 740 p. [ Links ]
PENTEADO-DIAS, A. M. 1984. Estudo comparativo do cordão nervoso nos Cerambycidae (Coleoptera). Revista Brasileira de Entomología 28 (3): 223-243. [ Links ]
PERTY, J. A. M. 1832. De insectorum in America meridionali habitantium vitae genere, moribus ac distributione geographica observationes nonnullae. En: Delectus animalium articulatorum quae in itinere per Brasiliam annis MDCCCXVII-MDCCCXX jussu et auspiciis Maximiliani Joseph I, Bavariae regis augustissimi peracto collegerunt Dr. J. B. de Spix et Dr. C. F. Ph. de Martius, Monachii. 61-124. [ Links ]
PITKIN, B. (editor). 2003. The Coleoptera Collectión and Card Index. Disponible en: http://www.nhm.ac.uk/research-curation/projects/coleoptera/ fecha de revisión: 14 noviembre 2008. [ Links ]
QUENTIN, R. M. ; VILLIERS, A. 1972. Un nouveau Parandrinae de Madagascar [Col. Cerambycidae]. Bulletin de la Société Entomologique de France 77: 208-209. [ Links ]
ROSALES, C. J. 1966. Contribución al conocimiento de los Prioninae (Coleoptera: Cerambycidae) de Venezuela. Revista de la Facultad de Agronomía de la Universidad Central de Venezuela 11: 1-237. [ Links ]
SAALAS, U. 1936. Über das Flügelgeäder und die phylogenetische Entwicklung der Cerambyciden. Annales zoologici Societatis zoologicae-botanicae fennicae Vanamo 4 (1): 1-198. [ Links ]
SAITO, A. 1990. Female reproductive organs of cerambycid beetles from Japan and the neighbouring areas. I. Philini through Atimini. Elytra 18 (2): 231-260. [ Links ]
SANTOS-SILVA, A. 2001a. Parandrinae Neotropicais (Coleoptera, Cerambycidae) II: espécies de Hesperandra Arigony, 1977 com submento elevado e marginação lateral do protórax dos machos incompleta. Revista Brasileira de Entomología 45 (3): 221-241. [ Links ]
SANTOS-SILVA, A. 2001b. Parandrinae Neotropicais (Coleoptera, Cerambycidae) I: redescrição de Parandra degeeri Thomson, 1867 e descrição de uma nova espécie. Revista Brasileira de Entomología 45 (3): 215-219. [ Links ]
SANTOS-SILVA, A. 2002. Notas e descrições em Parandrini (Coleoptera, Cerambycidae, Parandrinae). Iheringia, Série Zoologia 92 (2): 29-52. [ Links ]
SANTOS-SILVA, A. 2003a. Notas, descrições, sinonímias e revalidação em Hesperandra Arigony, 1977 (Coleoptera, Cerambycidae, Parandrinae). Revista Brasileira de Entomología 47 (1): 119-131. [ Links ]
SANTOS-SILVA, A. 2003b. Espécies de Hesperandra (Tavandra) Santos-Silva, com pilosidade abundante no metasterno (Coleoptera, Cerambycidae, Parandrinae). Revista Brasileira de Zoologia 20 (3): 389-395. [ Links ]
SANTOS-SILVA, A. 2007. Nova espécie de Hesperandra (Zikandra) Santos-Silva e chave para o subgênero (Coleoptera, Cerambycidae, Parandrinae). Les Cahiers Magellanes 66: 11p. [ Links ]
SANTOS-SILVA, A.; MARTIS, U. R. 2000. Nova espécie de Parandrinae (Coleoptera, Cerambycidae) da Região Neotropical. Revista Brasileira de Entomología 44 (3/4): 167-170. [ Links ]
SANTOS-SILVA, A.; SHUTE, S. 2009. The identity of Parandra laevis Latreille, 1804 and nomenclatural changes in the Parandrinae (Coleoptera, Cerambycidae). Zookeys 25: 19-35. [ Links ]
SAY, T. 1835. Descriptions of new North American coleopterous insectes, and Observations on some already described. Boston Journal of Natural History 1 (2): 151-203. [ Links ]
SCHJÖDTE, J. C. 1865. On the classificatión of cerambyces, with particular regard to the Danish Fauna. The Annals and Magazine of Natural History 15 (Serie 3): 182-209. [ Links ]
SHARP, D. 1878. On some longicorn Coleoptera from the Hawaiian Islands. The Transactions of the Entomological Society of London 1878: 201-210. [ Links ]
STURM, J. 1826. Catalog meiner Insecten-Sammlung, Erster Theil, Käfer. Nürnberg 1826: 1-207. [ Links ]
SVÁCHA, P. 1987. En: Svácha, P., y M. L. Danilevsky. 1987. Cerambycoid larvae of Europe and Soviet Unión (Coleoptera, Cerambycoidea). Part. I. Acta Universitatis Carolinae- Biología 30: 1-176. [ Links ]
Svácha, P.; WANG, J.; CHEN, S. 1997. Larval morphology and biology of Philus antennatus and Heterophilus punctulatus, and systematic positión of the Philinae (Coleoptera: Cerambycidae and Vesperidae).Annales de la Société Entomologique de France (N.S.) 33 (3): 323-369. [ Links ]
TAVAKILIAN, G. L. 2000. Les Parandrinae de Guyane (Coleoptera, Cerambycoidea). Coléoptères 6 (11): 147-174. [ Links ]
THOMSON, J. 1860-1861a. Essai d´une classificatión de la famille des cérambycides et matériaux pour servir à une monographie de cette famille. Paris. 404 p. [ Links ]
THOMSON, J. 1861b. Monographie de la famille des parandrides. Musée Scientifique ou Recueil d´Histoire Naturelle 2: 73-87. [ Links ]
THOMSON, J. 1861c. Note rectificative et corrections. Musée Scientifique ou Recueil d´Histoire Naturelle 2: 95-96. [ Links ]
THOMSON, J. 1867. Révision des parandrides (Insectes coléoptères). Physis Recueil d´Histoire Naturelle 1 (2): 106-118. [ Links ]
TRIPLEHORN, C. A.; JOHNSON, N. F. 2005. Borror and Delong´s Introductión to the Study of Insects. 7th Edition. Belmont, Brooks/ Cole. 864 p. [ Links ]
VILLIERS, A. 1980. Coléoptères Cerambycidae des Antilles Françaises I. Parandrinae, Prioninae, Lepturinae. Annales de la Société Entomologique de France (n.s.) 16 (1): 133-157. [ Links ]
VITALI, F. 2006. The real taxonomic positión of Spondylis florissantensis Wickham, 1920 (Coleoptera, Cerambycidae). Entomapeiron (P.S.) 1 (2): 13-28. [ Links ]
WHITE, A. 1853. Catalogue of the coleopterous insects in the collection of the British Museum, pt. VII. Longicornia I. 7:1-174. [ Links ]
WHITE, R. E. 1983. A Field Guide to the Beetles of North America. Peterson Field guides Series. Boston, Houghton Mifflin. 368 p. [ Links ]
YANEGA, D. 1996. Field Guide to Northeastern Longhorned Beetles (Coleoptera: Cerambycidae). Champaign, Illinois Natural History Survey Manual 6: 1-174. [ Links ]
ZIKÁN, J. F. 1948. O gênero Parandra Latr., 1804, com a descrição de 14 espécies novas (Col. Cerambycidae). Revista de la Sociedad Entomológica Argentina 14: 22-50. [ Links ]

