










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkRevista Colombiana de Entomología
versão impressa ISSN 0120-0488versão On-line ISSN 2665-4385
Rev. Colomb. Entomol. v.36 n.1 Bogotá jan./jun. 2010
Desarrollo, longevidad y oviposición de Heliconius charithonia (Lepidoptera: Nymphalidae) en dos especies de Passiflora
Development, longevity, and ovipositión of Heliconius charithonia (Lepidoptera: Nymphalidae) on two species of Passiflora
Carolina Millán J.1, Patricia Chacón C.2 y Germán Corredor3
1 Bióloga con énfasis en Entomología. Departamento de Biología, Universidad del Valle. A.A 25360 Cali. lepidoptera.azul@gmail.com. Autora para correspondencia.
2 Bióloga, Ph. D., Departamento de Biología, Universidad del Valle. A.A 25360 Cali. pachacon@univalle.edu.co.
3 Biólogo, M. Sc. Fundación Zoológica de Cali. Carrera 2 Oeste Calle 14 Santa Teresita, Cali. gcorredor@zoologicodecali.com.co.
Recibido: 29-oct-2009 - Aceptado: 1-may-2010
Resumen: El desarrollo de Heliconius charithonia en dos especies de plantas hospederas se estudió en el mariposario del Zoológico de Cali (Colombia) entre diciembre de 2007 y octubre de 2008. Se siguió el desarrollo de larvas provenientes de 90 y 83 huevos puestos en Passiflora adenopoda y P. rubra respectivamente. Se midió la duración de los cinco instares larvales, así como el peso y longitud pupal. Los adultos emergidos se marcaron, midieron, sexaron y se liberaron en el área de exhibición del mariposarío y se hicieron censos semanales para estimar la longevidad. La sobrevivencia larval fue mayor en P. adenopoda (76,4%) con respecto a P. rubra (33,9%). La mortalidad pupal alcanzó un 3% en P. rubra mientras en P. adenopoda todas las pupas fueron viables. Los resultados idican que P. adenopoda es el hospedero de oviposición más propicio para la cría masiva de H. charithonia, ya que en dicho hospedero se observó un mejor desarrollo larval, pupas más grandes y más pesadas, y los adultos mostraron mayor longitud alar y mayor longevidad (140 días vs 70 días). La preferencia de oviposición mostraron que del total de huevos (N = 357) el 71% fué depositado sobre P. adenopoda, aun en aquellos casos en que las hembras se desarrollaron sobre P. rubra.
Palabras clave: Tablas de vida. Mariposa cebra. Passiflora rubra. Passiflora adenopoda. Colombia.
Abstract: The development of Heliconius charithonia on two host plant species was studied at the butterfly house of the Cali Zoo (Colombia) between December 2007 and October 2008. The development of larvae from 90 and 83 eggs laid on Passiflora adenopoda and P. rubra, respectively, was followed. The duratión of the five larval instars was measured, as well as pupal weight and length. The emerged adults were marked, measured, sexed and released in the exhibitión area of the butterfly house and weekly censuses were conducted to estimate longevity. Larval survival was higher in P. adenopoda (76.4%) than in P. rubra (33.9%). Pupal mortality reached 3% in P. rubra while in P. adenopoda all pupae were viable. The results indicate that P. adenopoda is the host plant most suitable for mass rearing of H. charithonia, since on this host species we observed better larval development, larger and heavier pupae, and adults showed greater wing length and greater longevity (140 days vs 70 days). Ovipositión preferences showed that among all eggs (N = 357), 71% were laid on P. adenopoda even in those cases where females developed on P. rubra.
Key words: Life tables. Zebra butterfly. Passiflora rubra. Passiflora adenopoda. Colombia.
Introducción
En la búsqueda de alternativas de explotación sostenible de la biodiversidad, el desarrollo de trabajos de investigación es prioritario, de manera que se consiga adoptar estrategias que estimulen la comercialización y el uso de los recursos naturales en planes de educación para la preservación de la fauna (Andrade 2001). Una de las alternativas para la conservación de bosques tropicales es la cría comercial de mariposas ornamentales, un recurso forestal promisorío que reduce la pérdida de diversidad ejercida por la captura indiscriminada de poblaciones naturales de lepidópteros en Colombia (Gómez-S 2006).
La cría masiva de mariposas promueve un fuerte lazo entre la conservación y el desarrollo porque disminuye la presión sobre poblaciones naturales y provee individuos de mejor calidad y en mayor cantidad a los mercados locales e internacionales (Fagua et al . 2002). En Colombia, donde se conocen aproximadamente 3.019 especies de mariposas que representan el 61,9% de las especies conocidas para el Neotrópico (Andrade 2002), varias instituciones educativas como la Fundación Zoológica de Cali crían 25 especies de maripo sas, el Jardín Botánico del Quindío cuenta con 36 especies (Gómez et al . 2008) y empresas dedicadas al biocomercio, como Alas de Colombia, producen unas 40 especies (Restrepo y Wilches 2008) ya sea para exhibiciones en vivo o para la comercialización de adultos y artesanías.
Un factor clave a evaluar cuando se quiere mejorar y conocer la productividad de mariposas, es la calidad alimenticia que presentan las plantas hospederas a los estados inmaduros (Catta-Preta y Zucolto 2003). En el caso de la subfamilia Heliconiinae (Lepidoptera: Nymphalidae), ampliamente distribuida en Colombia desde el nivel del mar hasta los 2600 msnm (García et al . 2002), los adultos depositan sus huevos únicamente sobre pasionarias como resultado de una estrecha relación coevolutiva entre ellas (Gilbert 1998). Ejemplos del nivel de especialización de las mariposas del género Heliconius en plantas de la familia Passifloraceae son las especies H. hewitsoni (Hewitson, 1875) y H. cydno (Doubleday, 1847); la primera sólo oviposita en individuos de Passiflora pittieri Masters mientras que la segunda puede depositar huevos sobre dos especies de pasifloras (P. rubra Linnaeus y P. maliformis Linnaeus) (Gilbert 1998). Por su parte, H. erato (Linnaeus, 1758) es una especie oligófaga que puede alimentarse de P. misera Kunth, P. capsularis Linnaeus, P. alata Curtis, P. edulis Sims, P. suberosa Linnaeus, P. caerulea Linnaeus y P. elegans Sims (Menna-Barreto y Araujo 1985). En ese mismo sentido, Bianchi y Moreira (2005) encontraron que larvas de Dione juno (Cramer, 1779) pueden sobrevivir diferencialmente sobre cinco de diez especies de Passiflora. Por su parte, las larvas de la mariposa cebra H. charithonia (Linnaeus, 1767) son capaces de desarrollarse en especies como P. adenopoda en la que otras especies de Heliconius no pueden sobrevivir (Espinoza 2003), debido a la eficiente defensa mecánica que proveen los tricomas en forma de gancho del subgénero Decaloba De Candolle (Género Passiflora) (McDougal 1994; McDougal y Torsten 2004), que perforan el integumento y ocasionan la muerte de las larvas por pérdida de hemolinfa (Gilbert 1971). Para especies de Passiflora con tricomas en forma de gancho se registran dos casos de herbivoría: H. charitonia y Dione moneta [Huber, 1825] (Benson et al . 1975; MacDougal 1994). Cardoso (2008) demostró que larvas de H. charithonia son capaces de evitar el daño ocasionado por los tricomas usando fuerza física y a su vez va diseminando hilos de seda sobre los mismos, retirando las puntas con las mandíbulas. Este fenómeno se corroboró al encontrar puntas de tricomas en las heces de los inmaduros de esta especie.
H. charithonia también se alimenta de Passiflora lobata (Killip) Hutch, P. biflora Lamarck y P. bicornis Houston ex. Miller, (Espinoza 2003; Cardoso 2008), y se han registrado visitas usuales de hembras silvestres en P. rubra (Millán et al . 2009), todas especies del subgénero Decaloba. Teniendo en cuenta que H. charithonia es una de las especies de Heliconius más ampliamente distribuidas en Colombia (0 a 1600 msnm) (García et al . 2002) y su producción es de primordial interés para el mariposarío del Zoológico de Cali, se propuso comparar su desarrollo, longevidad y oviposición en dos plantas hospederas comúnmente usadas para la cría masiva de esta especie.
Materiales y Métodos
Plantas hospederas y obtención de huevos. En el vivero de la Fundación Zoológico de Cali (970 msnm, 3°26’56,08”N, 76°37’27,15”W), sembramos dos especies de pasionaria (Passiflora adenopoda y P. rubra) en semilleros individuales y se cultivaron sin adición de abonos ni insecticidas comerciales. La escogencia de estas dos especies se hizo con base en su amplia distribución en la geografía colombiana (McDougal 1994) y su fácil manejo en condiciones de invernadero, además de registrarse como hospederas usuales de Heliconius charithonia (Beccaloni et al . 2008).
Posteriormente, las plantas se trasplantaron a materas plásticas y se mantuvieron en invernadero durante dos o tres meses hasta alcanzar robustez foliar suficiente para el desarrollo exitoso de las larvas de H. charithonia durante los experimentos de oviposición. Durante cinco semanas, plantas de ambas especies se llevaron diariamente al recinto de las mariposas parentales y una vez obtenidas las posturas, las plantas se trasladaron al laboratorío para continuar con el seguimiento del desarrollo larval. De esta manera no se manipularon los huevos evitando así alterar su viabilidad.
Desarrollo de estados inmaduros. Se realizó en el laboratorio del mariposarío del Zoológico de Cali a una temperatura de 26 a 31°C y 41 a 54% de humedad relativa. Cada planta albergó entre 10 a 12 larvas asegurando su permanencia mediante adición de vaselina comercial en la parte basal de sus ramas para evitar el acceso a hormigas o chinches depredadores. El seguimiento del desarrollo larval se hizo a diario. Para cada estadio larval se tomaron medidas de duración (días). Todas las pupas se midieron y se pesaron (balanza analítica Ohius ± 0,03 mg) y se registró la duración del estado pupal. Los pedúnculos de las pupas se pegaron a alfileres con silicona líquida, luego se fijaron en pizarras y se depositaron en una cámara de vidrío donde constantemente se humedecieron hasta la emergencia de los adultos.
Longevidad. Todos los individuos se marcaron con números en el lado ventral de las dos alas posteriores, utilizando marcador indeleble no tóxico de punta fina de color negro (Sharpie®) para reconocer las mariposas criadas sobre P. adenopoda y rojo para aquellas provenientes de P. rubra. Para cada individuo se registró el día de emergencia, el sexo y la longitud del ala anterior. Los adultos se alimentaron ad libitum de plantas nectaríferas y productoras de polen (Impatiens balsamina Linnaeus, Lantana camara Linnaeus, Stachytarpheta cayenensis Linnaeus, Ixora coccinea Linnaeus, Hibiscus rosasinensis Linnaeus, Pentas lanceolata Forssk., Senecio confusus Britten, Asclepias curassavica Linnaeus, Hamelia patens Jacquin), adicionalmente se dispuso de comederos con preparaciones de polen artificial y cebos de fruta en descomposición (banano, borojó, guayaba, guanábana, mango y uchuva). Una vez por semana se hicieron lecturas de sobrevivencia por conteo de individuos marcados, aprovechando el comportamiento gregarío de las mariposas Heliconius a la hora habitual de percha (17:00 – 18:00 horas aprox.) (Chacón y Montero 2007), en sitios específicos dentro del mariposario.
Preferencia de oviposición. Para conocer si la selección de la planta de oviposición depende de la alimentación recibida durante el estado larval, se separaron por un período de tres semanas, cinco hembras provenientes de larvas alimentadas con P. rubra y cinco hembras nacidas en P. adenopoda. Se mantuvieron en dos mariposarios auxiliares (3 x 2 x 2m) bajo condiciones ambientales controladas de 22 a 31°C y 58 a 82 % de H.R. Durante la primera semana se dispusieron equidistantes en este lugar dos plantas de cada especie de Passiflora y se cuantificó el número de eventos de postura y el número de huevos depositados en cada hospedero. El mismo protocolo se realizó durante la segunda semana pero se dispusieron cuatro plantas del mismo hospedero (P. rubra), y así mismo durante la tercera semana, cuando la oferta correspondió exclusivamente a P. adenopoda.
Análisis de datos. Para el ciclo de vida de H. charithonia en cada hospedero, se elaboraron tablas de supervivencia de cohortes (Begon et al . 1999) que incluyen las siguientes variables: ax: Número de individuos al inicio de cada fase. lx: Proporción de la cohorte original que sobrevive al inicio de la fase. dx: Proporción de la cohorte original que muere durante cada fase. qx: Tasa de mortalidad. kx: Fuerza de mortalidad. Se compararon las medias de las variables de respuesta (longitud, peso y duración pupal, longitud alar y duración total del ciclo), entre individuos criados sobre las dos plantas hospederas, mediante prueba de t, previo análisis de normalidad por la prueba de Kolmogorov – Smirnov. Para las variables no normales se aplicó la prueba no para métrica de Mann-Whitney. Se aplicó un análisis de varianza factorial para comparar la duración de los cinco estadios larvales, en las dos plantas hospederas, seguido de una prueba múltiple de Tukey (Statistica versión 7.0) (StatSoft Inc. 2004).
Resultados
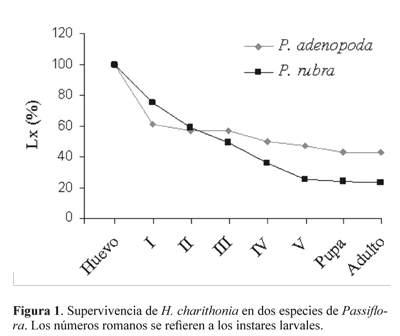
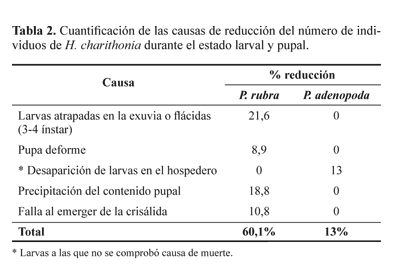
Tablas de vida. El seguimiento de H. charithonia en P. rubra y en P. adenopoda se realizó a partir de 83 y 90 huevos respectivamente (Tabla 1), encontrándose que la mortalidad total (Kx) fue mayor en P. rubra (0,64) comparada con P. adenopoda (0,35). La supervivencia de cada estado en las dos plantas hospederas muestra que en P. adenopoda la mortalidad durante la emergencia del huevo fue mayor que en P. rubra, pero en esta última la mortalidad aumentó a partir del primer ínstar (Fig. 1). Las larvas alimentadas con P. rubra murieron principalmente en el paso de tercer a cuarto ínstar (qx = 0,30), mientras que en P. adenopoda no se pudo comprobar la muerte de ningún individuo y se atribuyó a larvas desaparecidas del hospedero (qx = 0,12) (Tabla 1). La mortalidad en P. rubra se debió a larvas atrapadas en la exuvia, pupas deformes, precipitación del contenido pupal y fallas en la emergencia de la crisálida (Tabla 2). Del total de 19 adultos emergidos en P. rubra, 65,5 % fueron machos y de los 39 correspondientes a P. adenopoda, 63 % fueron machos.
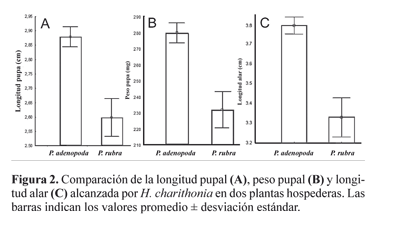
Peso corporal. Las pupas desarrolladas sobre P. adenopoda resultaron significativamente más grandes (U = 1156, N = 39, P << 0.01) (Fig. 2A) y más pesadas (t = 8.06, g.l.= 164, P << 0,01) (Fig. 2B) que aquellas observadas en P. rubra. En concordancia con lo observado para los estados juveniles, los adultos emergidos de P. adenopoda mostraron una longitud alar mayor (U= 834, N= 58,108; P << 0.01) (Fig. 2C).
Duración del ciclo de vida. Se encontraron diferencias significativas en la duración de los cinco estadios larvales entre hospederos (F = 20,013, P << 0,01), así como se halló interacción entre los factores hospedero y estadio larval (F = 21,368, P << 0,01). Las diferencias fueron mayores en larvas jóvenes (L2 y L3) ya que se retrasó el desarrollo cuando se alimentaron de P. rubra mientras que el último estadio (L5) tuvo mayor duración en P. adenopoda y así mismo el estado de pupa fue significativamente más largo sobre este hospedero (Tabla 3).
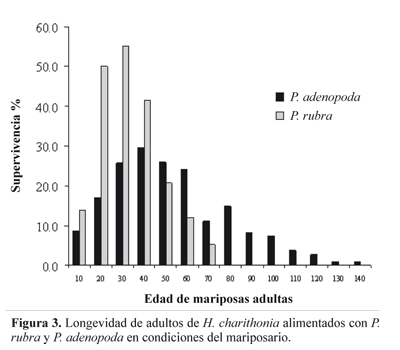
Longevidad. Durante los 4,7 meses de lecturas de longevidad, se identificaron dos sitios de percha: El primero sobre enredaderas situadas en las márgenes de la fuente de agua artificial del mariposarío y el segundo en el envés de hojas de asteráceas; en ambos casos, se encontraron agregaciones de ocho a 32 individuos. La Figura 3 describe la emergencia y sobrevivencia de adultos obtenidos de larvas criadas sobre P. adenopoda y P. rubra. El extremo izquierdo ilustra la etapa inicial donde los individuos eran liberados a medida que emergían de la pupa; el punto más alto corresponde al máximo de individuos obtenidos y el extremo derecho equivale a la desaparición de los imagos con el tiempo. Se obtuvo un máximo de longevidad de 140 días para individuos emergidos de P. adenopoda y de 70 días para mariposas sobrevivientes de la cría sobre P. rubra. Adicionalmente, la muerte de las mariposas obtenidas con P. rubra fue más abrupta, mientras que las mariposas asociadas a P. adenopoda murieron paulatinamente.
Preferencia de oviposición. Hubo menos eventos de oviposición de hembras obtenidas de larvas criadas sobre P. rubra, estas hembras no ovipositaron de manera continua a lo largo de las tres semanas del experimento (Tabla 4). Las mismas hembras produjeron 104 huevos en total y prefirieron depositarlos sobre P. adenopoda (60) que sobre P. rubra (44). Por su parte, las hembras obtenidas de larvas criadas sobre P. adenopoda pusieron continuamente a lo largo de las dos primeras semanas y no lo hicieron en la tercera semana; estas hembras produjeron más del doble de huevos (253) y los depositaron exclusivamente sobre P. adenopoda. El porcentaje de eclosión fue un poco mayor en huevos provenientes de hembras criadas sobre P. adenopoda.
Discusión
En la cría de la mariposa cebra, la productividad fue 2,3 veces mayor sobre el hospedero P. adenopoda ya que la supervivencia, desde el primer ínstar hasta la emergencia de los adultos, alcanzó un valor del 70,9% en comparación con P. rubra (30,6%). La baja mortalidad de larvas alimentadas con P. adenopoda en contraste con P. rubra, sugieren que P. adenopoda es un recurso más apropiado y que la mariposa cebra se ha especializado sobre este hospedero, mientras que con P. rubra se afecta el desarrollo larval con la consecuente producción de pupas no viables.
Las larvas que sobrevivieron sobre P. rubra invirtieron más tiempo en completar su desarrollo y produjeron pupas de menor tamaño y peso; en consecuencia, los adultos fueron menos longevos y fecundos. En cambio, sobre P. adenopoda el estado larval transcurrió más rápido aunque el período de pupa fue significativamente más largo, lo cual podría atribuirse a que crisálidas más grandes y más pesadas requieren más tiempo para completar su desarrollo y producir adultos de mayor tamaño. Los imagos resultantes de la alimentación con P. adenopoda fueron más longevos que los imagos emergidos de P. rubra; lo cual es ventajoso para la cría masiva de H. charithonia cuando se quiere usar para exhibición en insectarios. La proporción de machos fue similar entre plantas hospederas (63% y 65,5%) y se aproxima a los estudios de Fleming et al . (2005) quienes notaron que ese sexo conformó un 68% de las capturas en poblaciones naturales de la mariposa cebra, en Miami, Florida, EEUU.
Larvas bien nutridas pueden originar adultos con mayor capacidad de reproducción (Rodrigues y Moreira 2002, 2004). La oviposición diaria promedio de H. charithonia es directamente proporcional al tamaño del adulto, reflejada en la longitud de las alas anteriores (Dunlap-Pianka et al . 1979); en el presente estudio la oviposición de hembras de H. charithonia criadas con P. adenopoda fue 2,4 veces mayor a la observada en las hembras emergidas de P. rubra. Patrones similares han sido reportados por autores como Rodrigues y Moreira (2002, 2004), quienes atribuyeron una mayor calidad nutricional a P. misera en la cría de la mariposa H. erato, basados en que las hembras obtenidas presentaron mayor tamaño y capacidad reproductiva con respecto a las hembras desarrolladas en otras especies de Passiflora.
La selección de la planta para oviposición es una de las actividades más importantes para las mariposas, ya que afecta la viabilidad de la progenie determinando la probabilidad de éxito de la oruga al estado adulto (Singer y Ehrlich 1991). Diversas especies de mariposas pueden discriminar colores y formas cuando forrajean néctar o cuando buscan una planta hospedera para oviposición (Crane 1955; Rausher 1978; Traynier 1986; Weiss 1995; Kelber 1999). Además, algunos estudios muestran la asociación entre la forma de las hojas con el número de huevos depositados (Rausher 1978; Traynier 1986; Roessingh y Stadler 1990), auxiliando el encuentro de las plantas hospederas que las mariposas consideran más adecuadas para el desarrollo de su prole. En los experimentos de preferencia de oviposición, se observó que las hembras de H. charithonia depositaron el 71% de sus huevos sobre plantas de P. adenopoda y que incluso prefieren ovipositar sobre esta especie aunque hayan pasado todo su estado larval sobre P. rubra. Resultados similares fueron obtenidos por Menna- Barreto y Araujo (1985) quienes cuantificaron la oviposición de H. erato sobre nueve especies de Passiflora, encontrando preferencia por P. misera con una tasa de postura de 2,5 y P. suberosa (1,7), hospederos con los cuales las larvas se desarrollaron normalmente y generaron alta productividad de adultos. Además, observaciones ocasionales P. misera fue la única especie que presentó en campo huevos de la mariposa, pese a que las ocho pasionarias restantes también mostraron una amplia distribución en la zona de estudio.
Con relación al porcentaje de eclosión de huevos según la hembra de origen, se obtuvo un valor un poco más bajo en posturas de hembras alimentadas con P. rubra (70,2%) que en posturas de hembras alimentadas con P. adenopoda ( 76,7%), sugiriendo que este último hospedero podría estar presentado mejores cualidades para las hembras reproductivas, lo cual se vislumbra desde el proceso de escogencia preferencial de la hembra de H. charithonia por P. adenopoda.
La mariposa cebra puede criarse sobre las dos especies de Passiflora, pero P. adenopoda es un recurso de mayor calidad ya que se observa un mejor desarrollo larval, se obtienen pupas sanas y adultos más grandes y longevos; los cuales, en condiciones experimentales, prefieren ovipositar sobre este mismo hospedero pareciendo ignorar la disponibilidad de P. rubra, la cual parece ser un hospedero alternativo para la mariposa. A futuro se hace necesarío realizar estudios que contemplen la distribución y abundancia de ambas especies de Passiflora en zonas verdes de la ciudad de Cali, para estimar la disponibilidad de una especie con respecto a la otra; y determinar cuál de ellas es más utilizada y si realmente P. rubra es un hospedero alternativo ante la escaces de P. adenopoda en la naturaleza.
Agradecimientos
A la Fundación Zoológico de Cali por su apoyo económico. A los técnicos del mariposarío José Alejandro Perdomo, Diego Villaquirán y Juan David Ayala por su acompañamiento permanente y su colaboración en el cuidado de las mariposas. Al profesor Gilson Moreira por su valiosa ayuda en la consecución de literatura. A Carmen Elisa Posso y el laboratorío de Entomología de la Universidad del Valle por el apoyo logístico. A Jimmy Cabra por su ayuda en el análisis estadístico. A Mauricio Ramírez, a los revisores y al editor por sus sugerencias y acertados aportes al manuscrito.
Literatura citada
ANDRADE, M. G. 2001. Biodiversidad y conservación de la fauna colombiana, pp. 35-47. En: Muñoz, P. (Ed.). Memorias. Primer Congreso Colombiano de Zoología. Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá, D. C. 340 p. [ Links ]
ANDRADE-C, M. G. 2002. Biodiversidad de las mariposas (Lepidoptera: Rhopalocera) de Colombia, pp. 153-172. En: Costa, C., Vanin, S.A. y Lobo, J. M. (Eds.). Proyecto de Red. Iberoamericana de biogeografía y entomología sistemática PrIBES. Monografías Tercer Milenio. Zaragoza. 328 p. [ Links ]
BECCALONI, G.; VILORIA, A. L.; HALL, S. K.; ROBINSON, G. S. 2008. Catalogue of host plants of the Neotropical butterflies. Monografías tercer milenio. Sociedad Entomológica Aragonesa, Zaragoza (8): 430 p. [ Links ]
BEGON, M.; HARPER, J.L.; TO WNSEND, S.R. 1999. Ecología: Individuos, Poblaciones y comunidades. Ediciones Omega S.A. Barcelona. 1068 p. [ Links ]
BENSON, W.W.; BROWN, K.S.; GILBERT , L.E. 1975. Coevolutión of plants and herbivores: Passión flower butterflies. Evolution 29: 659-680. [ Links ]
BIANCHI, V.; MOREIRA, G. R. P. 2005. Preferência alimentar, efeito da planta hospedeira e da densidade larval na sobrevivencia e desenvolvimento de Dione juno (Cramer) (Lepidoptera, Nymphalidae). Revista Brasileira de Zoologia 22 (1): 43-50. [ Links ]
CARDOSO, M. 2008. Herbivore Handling of a Plant´s Trichome: The Case of Heliconius charithonia (L.) (Lepidoptera: Nymphalidae) and Passiflora lobata (Killip) Hutch. (Passifloraceae). Neotropical Entomology 37 (3): 247-252. [ Links ]
CATTA-PRETA, P. D.; ZUCOLTO , F. S. 2003. Oviposition behavior and performance aspects of Ascia monuste (Godart, 1919) (Lepidoptera, Pieridae) on kale (Brassica oleraceae var. acephala).Revista Brasileira de Entomología 47 (2): 169-174. [ Links ]
CHACÓN, I.; MONTERO, J. 2007. Butterflies and Moths of Costa Rica. INBio. Instituto Nacional de Biodiversidad. Banco Mundial. MINAE. 366 p. [ Links ]
CRANE, J. 1955. Imaginal behavior of a Trinidad Butterfly, Heliconius erato hydara Hewitson, with especial reference to the social use color. Zoológica 40: 167-196. [ Links ]
DUNLAP-PIANKA, H.; BOGGS, C. L.; GILBERT , L. E. 1979. Ovarian Dynamics In Heliconiine Butterflies: Programmed Senescence Versus Eternal Youth. Science 197: 487-490. [ Links ]
ESPINOZA, B. 2003. Available species. Heliconius charithonia, Disponible en: http://darnis.inbio.ac.cr/FMPro?-DB=UBIpub.fp3&-lay=WebAll&-Format=/ubi/detail.html&-Op=bw&id=5059&- Find. Fecha última revisión: 30 Agosto 2009. [ Links ]
FAGUA, G.; GÓMEZ, R.; GÓMEZ-MEJÍA, A. 2002. Estudio de viabilidad para la cría de mariposas y coleópteros como alternativa productiva para la regeneración del bosque en territorios dedicados a la siembra de cultivos ilícitos en San José del Guaviare (Colombia). Boletín Sociedad Entomológica Aragonesa (30): 223-224. [ Links ]
FLEMING, T.; SERRANO, D.; NASSAR, J. 2005. Dynamics of a subtropical populatión of the cebra longwing butterfly Heliconius charithonia (Nymphalidae). Florida Entomologist 88 (2): 169 - 179. [ Links ]
GARCÍA-ROBLEDO, C.; CONSTANTINO, L.M.; HEREDIA, M.D.; KATTAN, G. 2002. Mariposas comunes de la Cordillera Central de Colombia. Wildlife Conservation Society y los Autores. 41- 43. [ Links ]
GILBERT, L.E. 1971. Butterfly-plant coevolution: Has Passiflora adenopoda won the selectional race with heliconiine butterflies? Science 172: 585-586. [ Links ]
GILBERT , L. 1998. Coevolución de mariposas y enredaderas. Revista Investigación y Ciencia (73): 64-72. [ Links ]
GÓMEZ, A., LONDOÑO, J.; ORTEGA, A. 2008. Mariposarío del jardín Botánico del Quindío, una muestra de lepidópteros de Colombia para el mundo. Cali. Universidad Autónoma de Occidente. Resúmenes XXXV Congreso Sociedad Colombiana de Entomología: 206. [ Links ]
GÓMEZ-S, R. 2006. Plan de manejo propuesto para la cría de mariposas promisorias como alternativa productiva para comunidades indígenas de la Amazonia colombiana. Boletín Sociedad Entomológica Aragonesa 38: 451-460. [ Links ]
KELBER, A. 1999. Ovipositing butterfly use a red receptor to see green. Journal of Experimental Biology 202: 2619-2630. [ Links ]
MACDOUGAL, J. M. 1994. Revisión of Passiflora subgenus Decaloba sectión Pseudodysosmia (Passifloraceae). Systematic Botany Monographs 41: 1-146. [ Links ]
MACDOUGAL, J.M.; TORSTEN, U. 2004. Passiflora: Passionflowers of the world. Timber press Inc. Printed in China. 424 p. [ Links ]
MENNA-BARRETO , V.; ARAUJO, A. 1985. Evidence for host plant preferences in Heliconius erato Phyllis from Southern Brazil (Nymphalidae). Journal of Research on the Lepidoptera 24 (1): 41-46. [ Links ]
MILLÁN, C.; CHACÓN, P.; GIRALDO, A. 2009. Estudio de la comunidad de lepidópteros diurnos en zonas naturales y sistemas productivos del municipio de Caloto (Cauca, Colombia). Boletín científico. Centro de museos. Museo de historia natural, Caldas 13 (1): 185 - 195. [ Links ]
RAUSHER, M. 1978. Search image for leaf shape in a butterfly. Science 200: 1071-1073. [ Links ]
RESTREPO, P.; WILCHES, V. 2008. Biocomercio de Lepidoptera Rhopalocera en Colombia: Realidad demostrada por ALAS DE COLOMBIA, mariposas nativas Ltda. Cali. Universidad Autónoma de Occidente. Resúmenes XXXV Congreso Sociedad Colombiana de Entomología, pp. 206. [ Links ]
RODRIGUES, D.; MOREIRA, G. R. P. 2002. Geographical variation in larval - host plant use by Heliconius erato (Lepidoptera: Nymphalidae) and consequences for adult life history. Brazilian Journal of Biology, Sao Carlos (62): 321-332. [ Links ]
RODRIGUES, D.; MOREIRA, G. R. P. 2004. Seasonal variation in larval - host plant and consequences for Heliconius erato (Lepidoptera: Nymphalidae) adult body size. Austral Ecology Carlton (29): 437-445. [ Links ]
ROESSINGH, P.; STADLER, E. 1990. Foliar form, colour and surface characteristics influence ovipositión behavior in the cabbage root fly Delia radicum. Entomología Experimentalis et Applicata 57: 93-100. [ Links ]
SINGER, M.; P. R. EHRLICH. 1991. Host specializatión of Satyrinae butterflies, and their responses to habitat fragmentatión in Trinidad. Journal of Research on the Lepidoptera 30 (3-4): 248- 256. [ Links ]
TRAYNIER, R. M. M. 1986. Visual learning in assays of sinigrin as an ovipositión releaser for the cabbage butterfly, Pieris rapae. Entomología Experimentalis et Applicata 40: 25-33. [ Links ]
WEISS, W. 1995. Associative colour learning in nymphalid butterfly. Ecological Entomology 20: 298-301. [ Links ]

