










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkRevista Colombiana de Entomología
versão impressa ISSN 0120-0488versão On-line ISSN 2665-4385
Rev. Colomb. Entomol. v.36 n.1 Bogotá jan./jun. 2010
Nota científica
Registro de Mechanitis polymnia (Lepidoptera: Ithomiinae) en Solanum jamaicense y ciclo de vida en laboratorio
Record of Mechanitis polymnia (Lepidoptera: Ithomiinae) on Solanum jamaicense and life cycle in the laboratory
CARLOS EDUARDO GIRALDO S.1 y SANDRA I URIBE S.2
1 Ingeniero Agrónomo. Grupo de Investigación en Sistemática molecular Universidad Nacional de Colombia sede Medellín. Estudiante Maestría en Biología convenio Universidad de Antioquia-Universidad Nacional. Correspondencia: Calle 59A No 63 - 20. Bloque 18-102 Universidad Nacional de Colombia sede Medellín. cegiral0@unalmed.edu.co. Autor para correspondencia.
2 Ingeniera Agrónoma, M. Sc., Ph. D., Profesora asociada Facultad de Ciencias, Posgrado de Entomología Universidad Nacional de Colombia sede Medellín, Calle 59A No 63-20. Bloque 18-102 Medellín, Colombia.
Recibido: 5-mar-2009 - Aceptado: 20-dic-2009
Resumen: Estados inmaduros de la mariposa Mechanitis polymnia (Lepidoptera: Ithomiinae) fueron recolectados sobre Solanum jamaicense en seis localidades de las cordilleras central y occidental de los Andes colombianos de los departamentos de Antioquia y Caldas. Observaciones de los hábitos de postura de las hembras fueron realizadas en campo. Se hacen anotaciones sobre morfología, desarrollo, comportamiento de los estados inmaduros y se describe el ciclo de vida bajo condiciones de laboratorio.
Palabras clave: Solanaceae. Planta hospedante.
Abstract: Immature stages of the butterfly Mechanitis polymnia (Lepidoptera: Ithomiinae) were collected on Solanum jamaicense in six localities of the western and central ranges of the Colombian Andes, from the departments of Antioquia and Caldas. Observations on the ovipositión habits of the females were made in the field. Notes were made on the morphology, development and behavior of the immatures stages, and the life cycle under laboratory conditions is described.
Key words: Solanaceae. Host plant.
Introducción
Las mariposas Ithomiinae (Lepidoptera: Nymphalidae) agrupan alrededor de 370 especies (Lamas et al . 2004), que pueden ser encontradas en el Neotrópico desde el norte de Argentina hasta México (Willmott y Freitas 2006). Los Ithomiinae se encuentran asociados normalmente a bosques húmedos, sin embargo Brown y Freitas (2003) mencionan que algunas especies pueden permanecer en pequeños fragmentos de bosque y en sistemas urbanos y suburbanos. Las especies del género Mechanitis prosperan en un amplio rango de hábitats agrícolas y bosques secundarios, donde crecen las plantas hospedantes de sus larvas (Young y Moffett 1979; García et al . 2002). La relación de la subfamilia Ithomiinae con las plantas de la familia Solanaceae ha sido bien documentada (Drummond y Brown 1987, 1999; Willmott y Mallet 2004), pero aún existe desconocimiento en cuanto a las plantas hospedantes específicas para las diferentes especies de Ithomiinae (Willmott 2008).
Las larvas del género Mechanitis, objeto del presente estudio, han sido registradas en diferentes países alimentándose de varias especies del género Solanum (Vasconcellos-Neto 1980, 1986; Acevedo 1992; Constantino 1997; Haber 2001), sin embargo no hay documentación precisa sobre la relación entre especies de Mechanitis y las especies de Solanum (Willmott Com. Personal). Al respecto, Willmott y Mallet (2004) en su recopilación sobre plantas hospedantes de Ithomiinae, registraron 23 plantas hospedantes para M. polymnia (Linnaeus, 1758) en Ecuador, Costa Rica y Brasil. Vale anotar que Beccaloni et al . (2008) realizaron el trabajo recopilatorío más actualizado sobre plantas hospedantes de mariposas neotropicales y señalan a Passiflora edulis Sims. como planta hospedante de M. polymnia para Colombia con base en la referencia de Castellar y Figueroa (1969); sin embargo, al revisar en detalle el trabajo de Castellar y Figueroa se evidencia que los estados inmaduros descritos corresponden a un individuo de la subfamilia Heliconiinae, posiblemente de la especie Eueides isabella Cramer, 1781 cuyo adulto es un comimético de las especies del género Mechanitis. En Colombia, otros registros señalan a Solanum torvum (Sw., 1788) y Solanum quitoense (Lam., 1794), como plantas hospedantes de M. polymnia (Constantino 1997). En el presente trabajo se registra por primera vez para Colombia a S. jamaicense como planta hospedante de Mechanitis polymnia y se realizan anotaciones sobre particularidades de la morfología, el comportamiento de las larvas y el ciclo de vida sobre dicha especie.
Materiales y Métodos
El material biológico fue obtenido a partir de cinco localidades del departamento de Antioquia, Amagá (1480 msnm, 6°01’29,94”N, 75°42’14,41”W ), Hispania (1022 msnm, 5°45’16,84”N, 75°55’04,89”W), Sopetrán (850 msnm, 6°29’58,76”N, 75°44’03,89”W), Valparaíso (1000 msnm, 5°41’34”N, 75°38’21”W), Venecia (1493 msnm, 5°57’40,66”N, 75°44’19,18”W) y Anserma en el departamento de Caldas ( 840 msnm, 5°10’31,64”N, 71°40’54,16”W). Los muestreos se realizaron entre los meses de Octubre de 2006 y Octubre de 2008. Las masas de huevos o larvas visualizadas fueron recolectadas directamente de las hojas de la planta hospedante, las cuales se tomaron con sus respectivos esquejes. Se tomaron muestras vegetales para la identificación taxonómica a nivel de especie en el herbarío MEDEL - Universidad Nacional de Colombia sede Medellín y material vegetativo para la reproducción y obtención de alimento. La cría de las larvas se realizó en el insectarío de la Universidad Nacional de Colombia sede Medellín, bajo las siguientes condiciones 1.538 msnm, 27°C de temperatura media y 45% de humedad relativa.
Se realizaron observaciones detalladas de cada estadío larval con un estéreomicroscopio Boeco 3500 a 45X. Diariamente se midió la longitud y el ancho de las larvas y se registró la presencia de las cápsulas cefálicas como indicadores de muda y la coloración de los segmentos abdominales. Los comportamientos de las larvas durante la alimentación fueron también registrados y descritos para cada estadío. Una vez obtenidos los adultos, se sexaron, registraron y liberaron en la jaula para adultos del insectarío para facilitar la cópula y reiniciar el ciclo. La confirmación taxonómica de las especies se realizó por los especialistas en cada uno de los respectivos grupos.
Resultados y Discusión
Planta hospedante. La planta hospedante fue identificada como Solanum jamaicense Mill. 1768. Dos muestras completas fueron depositadas en el herbarío MEDEL de la Universidad Nacional de Colombia sede Medellín y en el Herbarío de la Universidad de Antioquia (HUA).
Solanum jamaicense (Fig. 1F) se distribuye desde el sur de México hasta el Norte de Perú. (NHM 2008) y ha sido previamente registrada como planta hospedante de M. isthmia (M. polymnia isthmia H. W. Bates, 1863) en Costa Rica (Haber 2001). Aunque en Colombia esta especie de Solanum no se había registrado como planta hospedante del género Mechanitis, en este estudio se encontró como planta nutricia de las larvas de M. polymnia. Esta observación fue igual para las seis localidades estudiadas y su condición como planta hospedante se verificó por la alimentación de las larvas en condiciones de laboratorio. Otras hospedantes de esta especie, como S. torvum, previamente registrada como planta hospedante de M. polymnia (Constantino 1997), fue también encontrada con posturas en tres localidades (Sopetrán, Amagá y Valparaíso) pero en menor frecuencia.
Hábitos de postura. Las posturas se observaron siempre en el haz de la hoja, en grupos de 14 a 60 huevos (30,47 promedio huevos/postura +- 10,34; n = 34) (Fig. 1F), preponderantemente en plantas jóvenes o en el tercio inferior de plantas que superan 1 metro de altura (76,47% de las posturas en plantas de menos de 0,5m). De las 166 plantas examinadas 38 presentaron huevos. Solo tres posturas se encontraron en S. torvum y 35 en S. jamaicense. No hubo correlación entre la altura sobre el nivel del mar y el tamaño de la postura de M. polymnia (R = 0.0422; P = 0.86. Datos no mostrados). La mayoría de las plantas con más de un metro de altura se encontraban en estado de fructificación, cuando la planta presenta áreas foliares reducidas en comparación con las hojas bajeras.
Vasconcellos-Neto y Monteiro (1993), observaron un comportamiento similar para M. lysimnia (Fabricius, 1793) en un estudio realizado en el Sureste de Brasil. Estos investigadores encontraron que las hembras preferían ovipositar las hojas jóvenes y sin daños mayores causados por insectos o por eventos mecánicos. Además se demostró experimentalmente que las hembras prefieren plantas sin otros individuos co-específicos. Una situación similar podría estarse presentando en las hembras de M. polymnia dado que solo en dos de las plantas de S. jamaicense se encontró más de un grupo de huevos o grupos de larvas en la misma planta. En cuanto al comportamiento de oviposición se observó la hembra realizando sobrevuelos consecutivos a distancias muy cortas de la planta, tocando suavemente las hojas con sus patas. Posteriormente posándose sobre la hoja, y deslizando el extremo distal de su abdomen sobre la superficie de la hoja en un área reducida para finalmente comenzar la postura.
Estados inmaduros. Los huevos de M. polymnia son blancos recién ovipositados, pero ligeramente translúcidos cuando se examinan con detalle, bajo la luz del estéreomicroscopio. Tienen forma ovalada y 14 estrías longitudinales profundas que van desde el micrópilo hasta la base del huevo. Estrías transversales superficiales alrededor del huevo son también observables (10X) (Fig. 1A). Los huevos miden 1,5 mm de largo y 1 mm de diámetro en su mayor amplitud (tercio medio). La coloración de los huevos se torna gris oscuro aproximadamente dos horas antes de la eclosión.
Las larvas generalmente consumen totalmente el corión luego de su eclosión. Las larvas miden 3,6 mm (+/- 0,37) de largo y 0,5 mm de ancho. La cápsula cefálica es negra recién eclosionada, pero seis horas después se torna marrón. El cuerpo se observa verde oscuro después de comenzar la alimentación y posee un par de tubérculos reducidos en el pronoto y ocho pares de tubérculos laterales poco desarrollados (Fig. 1B). Las larvas son gregarias al comienzo de su ciclo, se alimentan posadas sobre el envés de la hoja y evitan la ingestión de los tricomas estrellados. Este comportamiento fue reportado también por Rathcke y Poole (1975) para M. polymnia. Las larvas pasan al envés de la hoja a través del orificio generado a medida que se alimentan y continúan con el mismo comportamiento de alimentación dejando a un lado los tricomas no ingeridos.
Las larvas de segundo instar alcanzan 5,93 mm de largo (+- 0,68) y 1 mm de ancho. Su cápsula cefálica es color marrón; los dos tubérculos blancos del pronoto se hacen más evidentes. Los tubérculos laterales son verde claro y miden 0,5 mm de largo. Las patas torácicas son negras y las pseudopatas son blancas. Las larvas de tercer instar alcanzan 10,17 mm de largo (+- 0,62) y 1,2 mm de ancho. Su cápsula cefálica es marrón y el abdomen verde oscuro. Los tubérculos laterales son blancos y amarillos en la base, con una mancha negra de forma circular; miden 0,8 mm de longitud. En el tercer estadío las larvas mantienen el comportamiento gregario, pero se ven más distanciadas entre sí. Su cápsula cefálica es marrón y el abdomen es gris oscuro sin líneas longitudinales en el dorso. Los tubérculos laterales son blancos y amarillos en la base, Las larvas de cuarto instar miden 18,37 mm de largo (+/- 0,87) y 1,8 mm de ancho. La cápsula cefálica es de color crema, los tubérculos son blancos y amarillos en la base, miden 1,8 mm de longitud; en la base se observa una mancha circular negra de 0,4 mm de diámetro. El abdomen es gris claro y se hace visible una línea amarilla longitudinal a lo largo del noto. En este instar las larvas se alimentan tanto en el haz como en el envés de la hoja y lo hacen de forma solitaria o en grupos de máximo dos o tres larvas. Las larvas de quinto instar miden 24,39 mm de largo (+/- 0,83) y 33 mm de ancho; conservan las características de coloración del cuarto instar, pero su cápsula cefálica es gris claro (Fig. 1C). Se alimentan en el haz y el envés de la hoja de manera solitaria o gregaria. La prepupa adquiere una coloración amarilla clara y detiene su alimentación. En condiciones de laboratorio, la prepupa se mantiene sobre la planta hospedante pero en condiciones naturales las prepupas nunca fueron observadas sobre las plantas hospedantes. Las pupas recién formadas son amarillas, con los bordes negros. 10 horas más tarde tienen una coloración dorada, la cual permanece hasta el momento de la emergencia de los adultos. Las pupas miden 18 mm de largo y 7 mm de ancho en vista lateral y 5 mm en vista ventral (Fig. 1D). Cuando el adulto está próximo a emerger la pupa se torna negra y se hacen visibles las manchas amarillas y naranja de las alas. Una vez el adulto emerge se tarda en promedio ocho horas en extender y endurecer completamente sus alas (Fig. 1E). Dado que no se encontraron diferencias significativas en la duración promedio de individuos procedentes de diferentes localidades (p=0.84. Datos no mostrados), el promedio de duración del ciclo de vida fue tomado de los datos aleatorizados. El ciclo de vida se inició con un total de 134 huevos, de los cuales 82 llegaron a estado adulto (61,19%); el 54% de estos fueron machos. El ciclo desde huevo hasta adulto tardó en promedio 32,6 días (+/- 1,80) (Tabla 1).
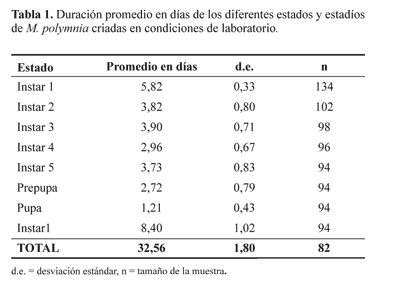
El estado con mayor mortalidad fue el de huevo. Hubo emergencia de microhymenópteros parasitoides en 32 de ellos (23,88%). El estado de pupa también presentó una alta mortalidad ocasionada por una enfermedad no identificada, pero los síntomas visibles se asemejan a aquellos que se presentan enfermedades bacterianas. En el ciclo de vida publicado por Constantino en 1997, cada instar larval tuvo una duración de cuatro días. La diferencia principal con el ciclo registrado en este estudio, es la duración de los instares tres y cinco que en promedio duraron un día menos.
Agradecimientos
Sinceros agradecimientos al Doctor Michael Nee curador Sistemática Botánica del New York Botanical Garden por su ayuda y entrenamiento en la identificación de las plantas hospedantes. Al Doctor Keith Willmott curador del Lepidoptera McGuire Center for Lepidoptera and Biodiversity Florida Museum of Natural History University of Florida por su ayuda en la confirmación de las especies y entrenamiento en el conocimiento de la subfamilia Ithomiinae. Al personal y propietarios de las Haciendas Canoas (Caldas), La Fe (Antioquia) y Finca Taboga (Antioquia) por su hospitalidad y por proporcionar el espacio para el desarrollo de esta investigación. Finalmente a la Facultad de Ciencias, la DIME (20101007738) y el Posgrado de Entomología de la Universidad Nacional de Colombia sede Medellín, por proporcionar los recursos y el insectarío para la realización de este trabajo.
Literatura citada
ACEVEDO, E. 1992. Reconocimiento de plagas y benéficos en el cultivo de tomate de árbol (Cyphomandra betacea (Cav.) en la zona cafetera de Colombia. Agronomía (Manizales-Colombia) 5 (1): 33-37. [ Links ]
BECCALONI, G. W.; VILORIA, A. L.; HALL, S. K.; ROBINSON, G. S. 2008. Catálogo de las Plantas Huésped de las Mariposas Neotropicales. m3m-Monografías Tercer Milenio, Volumen 8. Zaragoza -España. 536 p. [ Links ]
BROWN, K. S. JR.; FREITAS, A. V. 2003. Butterfly communities of urban forest fragments in Campinas, Sao Paulo, Brazil: structure, instability, environmental correlates, and conservation. Journal of Insect Conservatión 6: 217-231. [ Links ]
CASTELLAR, N.; FIGUEROA, A. 1969. Estudio biológico de dos formas de lepidópteros: Agraulis vanillae (Linn.) y Mechanitis veritabilis (Butler) en el maracuyá (Passiflora edulis var. flavicarpa, D.). Acta agronómica 19 (1): 17-30. [ Links ]
CONSTANTINO, L. M. 1997. Conocimiento de los ciclos de vida y plantas hospederas de lepidópteros diurnos de Colombia como estrategia para el manejo, uso y conservación de poblaciones silvestres. Seminarío Aconteceres Entomológicos. Octubre 30 y 31 (Medellín- Colombia). Páginas. [ Links ]
DRUMMOND, B. A. III.; BROWN, K. S. JR. 1987. Ithomiinae (Lepid.: Nymphalidae): summary of know larval foodplants. Annals of the Missouri Botanical Garden 74: 341-358. [ Links ]
DRUMMOND, B. A. III.; BROWN, K. S. JR. 1999. New records of larval host plants for Ithomiinae butterflies (Nymphalidae). Revista Brasileira de Biología 59 (3): 455-459. [ Links ]
GARCÍA, C. A.; CONSTANTINO, L. M.; HEREDIA, M. D.; KATAN, G. 2002. Mariposas comunes de la cordillera central de Colombia, Guía de campo. Editorial Feriva. Cali - Colombia. [ Links ]
HABER, W. A. 2001. Clearwing butterflies of Costa Rica (Ithomiinae). Disponible en: http://www.cs.umb.edu/~whaber/Monte/Ithomid/Ithomid-fram.html. Fecha de revisión: 25 Enero 2008. [ Links ]
LAMAS, G.; CALLAGHAN, C. J.; CASAGRANDE, M. M.; MIELKE, T. H; PYREZ, W.; ROBBINS, R. K.; VILORIA, A. L. 2004. Atlas of Neotropical Lepidoptera - Checklist: part 4ª Hesperoidea-Papilionoidea. Scientific Publications: Florida, Gainesville, Estados Unidos. 439 p. [ Links ]
NATURAL HISTORY MUSEUM. 2008. Solanaceae Source web site. Disponible en: http://www.nhm.ac.uk/research-curation/projects/solanaceaesource/. Fecha revision: 4 Febrero 2008. [ Links ]
RATHCKE, B. J.; POOLE, R. W. 1975. Coevolutionary race continues: butterfly larval adaptatión to plant trichomes. Science 187 (4172): 175-176. [ Links ] [ Links ]
VASCONCELLOS-NETO, J. 1986. Interactions between Ithomiinae (Lep., Nymphalidae) and Solanaceae. In: D´Arcy, W. G. (Ed.). Solanaceae: biology and systematics. Columbia University Press, New York, pp. 364-377. [ Links ]
VASCONCELLOS-NETO, J.; MONTEIRO, R. F. 1993. Inspection and evaluation of host plant by the butterfly Mechanitis lysimnia (Nymph., Ithomiinae) before laying a mechanism to reduce intraspecific competition. Oecología 95: 431-438. [ Links ] [ Links ]
WILLMOTT, K. R.; MALLET, J. 2004. Correlations between adult mimicry and larval host plants in Ithomiine butterflies. Proceedings of the Royal Society B. (Suppl.) 271: S266-S269. [ Links ] [ Links ]
YOUNG, A.; MOFFETT, M. 1979. Studies on the population biology of the tropical butterfly Mechanitis isthmia in Costa Rica. American Midland Naturalist 101 (2): 309-319. [ Links ]

