










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488On-line version ISSN 2665-4385
Rev. Colomb. Entomol. vol.36 no.2 Bogotá July/Dec. 2010
Evaluación de nematodos entomopatógenos para el manejo de Phyllophaga bicolor (Coleoptera: Melolonthidae)
Evaluation of entomopathogenic nematodes for the management of Phyllophaga bicolor (Coleoptera: Melolonthidae)
ELSA LILIANA MELO M.1, CARLOS ALBERTO ORTEGA O.2, ANDREAS GAIGL3 y ANTHONY BELLOTTI4
1 M. Sc. Centro Internacional de Agricultura Tropical (CIAT), Cali, Colombia. A.A. 6713, tel.: 445 0000, fax: 445 00 73. e.l.melo@cgiar.org. Autora para correspondencia
2 M. Sc. Laboratorios de Biotecnología LABIOTSA, Universidad Central del Ecuador. Quito-Ecuador, caortegao@gmail.com.
3 Ph. D. Universidad Nacional de Colombia, Facultad de Agronomía, Sede Bogotá, agaigl@unal.edu.co.
4 Ph. D. Centro Internacional de Agricultura Tropical - CIAT. A. A. 6713, Cali, a.bellotti@cgiar.org.
Recibido: 25-jun-2009 • Aceptado: 24-jul-2010
Resumen: Las larvas de escarabajos (Coleoptera: Melolonthidae) son plagas muy importantes en diversidad de cultivos y climas debido a las pérdidas que producen en la calidad y rendimiento de los productos agrícolas. Una alternativa para su manejo biológico son los nematodos entomopatógenos, parásitos generalistas de éstas, y controladores efectivos en pruebas de laboratorio y campo. Se evaluaron aislamientos de nematodos entomopatógenos nativos e introducidos sobre larvas de tercer estadio de Phyllophaga bicolor. Se determinaron la infectividad y mortalidad de aislamientos de Steinernema(S. riobravis-Sr, S. carpocapsae all Strain-Sc, Steinernemasp.-SNI, S. arenarium -Sa y S. feltiae-Sf1 y Sf2) y Heterorhabditis (H. bacteriophora-Hb1, Hb2 y Hb3) con una concentración de 10.000 JIs/ml, a 23°C y 70 ± 5% H.R. La evaluación se hizo en larvas de uno, dos y tres meses. Los aislamientos más infectivos fueron los de Heterorhabditis (93,75%-Hb2, 89,58%-Hb1 y 64,58%-Hb3), comparados con los de Steinernema(10,42%-SNI; 12,50%-Sr; 16,67%-Sa; 22,92%-Sc; 33,34%-Sf2; 66,67%-Sf1). Las menores mortalidades se produjeron con los aislamientos de Steinernema(0%-Sc y SNI; 2,08%-Sr; 6,25%-Sa; 6,25-Sf2 y 47,92%-Sf1) mientras que las mayores con los de Heterorhabditis (52,09%-Hb3; 72,92%-Hb1 y 91,67%-Hb2). Dependiendo de la madurez de la larva se presentaron diferencias en infección (1 = 65,64%; 2 = 42,50%; 3 = 29,17%) y mortalidad (1 = 53,33%; 2 y 3 promediaron 31,46%), notándose una disminución en la magnitud de estas variables al acercarse a prepupa. El género Heterorhabditis es el más promisorio para el manejo de chisas especialmente aplicado a las larvas jóvenes.
Palabras clave: Steinernema. Heterorhabditis. Control biológico. Chisa.
Abstract: Scarab larvae (Coleoptera: Melolonthidae) are important pests in a diversity of crops and climates owing to their damage to the quality and yield of agricultural products. An alternative for their biological control are the entomopathogenic nematodes, generalist parasites and effective control agents in laboratory and field trials. Isolates of introduced and native entomopathogenic nematodes were evaluated against third instar larvae of Phyllophaga bicolor. We determined the infectivity and mortality of isolates of Steinernema(S. riobravis-Sr, S. carpocapsae Strain all-Sc, Steinernemasp.-SNI, S. arenarium-Sa and S. feltiae-Sf1 and Sf2) and Heterorhabditis (H. bacteriophora-Hb1, Hb2 and Hb3) at a concentration of 10,000 IJs/ml at 23°C and 70 ± 5% R.H. The evaluation was done on larvae that were one, two and three months old. The most ineffective isolates were those of Heterorhabditis (93.75%-Hb2; 89.58% and 64.58%-Hb1-Hb3), compared to those of Steinernema(10.42%-SIN; 12.50%-Sr; 16.67%-Sa, 22.92%-Sc; 33.34%- Sf2; 66.67%-Sf1). The lowest mortalities resulted from Steinernemaisolates (0%-Sc and SNI; 2.08%-Sr, 6.25%-Sa; 6.25%-Sf2 and 47.92%-Sf1) while the highest were from Heterorhabditis (52.09%-Hb3; 72.92% and 91.67%-Hb1- Hb2). Differences were noted in infection (1 = 65.64%, 2 = 42.50%, 3 = 29.17%) and mortality (1 = 53.33%, 2 and 3 averaged 31.46%), depending on larval maturity, with a noticeable reduction in these variables when approaching the prepupa. The genus Heterorhabditis is the most promising for the white grub management, especially when applied to young larvae.
Key words: Steinernema. Heterorhabditis. Biological control. White grub
Introducción
Las larvas de la familia Melolonthidae (Coleoptera) son muy importantes desde el punto de vista ecológico por participar en el reciclaje de material orgánico y en el enriquecimiento del suelo. Sin embargo, varias especies se han convertido en plagas de importancia económica, entre las que se destacan especies de las subfamilias Melolonthinae, Dynastinae, Rutelinae y Cetoniinae; éstas representan gran parte de las pérdidas en zonas agrícolas debido a la gravedad del daño que producen y a su permanencia en los sitios donde se cultiva; este hecho se observa en el Oriente Antioqueño (Noroccidente colombiano), donde estos insectos se han incrementado desde 1988, igualmente el daño que ocasionan en los cultivos (Restrepo-Giraldo y López-Ávila 2000; Pardo-Locarno 2002; Londoño 2001).
Las larvas de Melolonthidae son plagas de importancia a nivel del suelo en diferentes cultivos del mundo; solamente las larvas de tercer instar causan daños significativos actuando como tierreros o trozadores al roer y consumir raíces; en las etapas avanzadas de los cultivos causan amarillamiento del área foliar, pasando por plasmólisis hasta la marchitez en plantas jóvenes, debido al consumo de raíces pequeñas completas y hasta corteza de raíces grandes lo que involucra la zona cambial; este daño depende igualmente de la población del rizófago, edad del cultivo y la cobertura vegetal que lo rodea, así como de los macro y micro organismos entomopatógenos y entomófagos cohabitantes en el sustrato (Posada 1993; Ortega 2005); este estado, además de ser el estado más voraz, es también el de mayor duración (171 días) comparado con el primer estadio larval que se desarrolla en 24 días y el segundo con un promedio de 29 días, lo que posibilita su daño por un tiempo mayor (CIAT 2004). El adulto no es tan dañino como la larva, pero ocasionalmente afecta las inflorescencias de cultivos como el maíz o el follaje de arbustos árboles y ciertas plantas anuales entre las que se destacan las familias Papilionaceae, Euphorbiaceae, Solanaceae, Cucurbitaceae, Poaceae, entre otras (Ortega 2005).
En Colombia estas larvas se conocen comúnmente como chisas, cuyos adultos son escarabajos o cucarrones que varían en tamaño, coloración, forma y hábitos de acuerdo con la especie. Varias especies de estos melolóntidos constituyen un complejo de plagas que afectan en forma severa una gran diversidad de cultivos, como plagas rizófagas o plagas de follaje y fruto (Restrepo-Giraldo y López-Ávila 2000).
Los mayores daños económicos que ocasionan estos rizófagos están relacionados con los cultivos de yuca, fríjol, papa, cereales, maíz y hortalizas, donde las pérdidas alcanzan entre el 10 y el 95% de las cosechas (Restrepo-Giraldo y López-Ávila 2000).
Experimentos realizados en campos de yuca en el Norte del Departamento del Cauca, sur occidente colombiano, con el complejo de larva de tercer instar de Phyllophaga spp. (Melolonthidae) mostraron pérdidas de 26% en material fresco y 30% en almidón; además en condiciones controladas se observó que densidades entre cinco a siete larvas por estaca consumen más del 95% de corteza y 35% de médula, en un período de dos meses, y que el daño realizado por una chisa resulta en el 53% de pérdidas de su potencial productivo por el consumo de la corteza, médula y raíces de las estacas; esto muestra la severidad del problema, especialmente si la siembra se da en el momento de la aparición del estado más agresivo de la plaga (Ortega 2005).
Colecciones de larvas realizadas en este sitio han mostrado una multiplicidad de especies, donde las más abundantes son Plectris fassli (Moser, 1919), P. pavida (Burmeister, 1855), Anomala cincta Say, 1835, Ceraspis innotata (Blanchard, 1850) y un complejo de especies del género Phyllophaga, género más importante entre las más dañinas por su carácter rizófago estricto, ciclo anual y larvas de mayor porte, destacándose P. menetriesi Blanchard, 1850 y P. bicolor (Moser, 1921) (Pardo-Locarno 2002; CIAT 2004). Esta última no reportada antes para Colombia, ni mencionada por su importancia agrícola (J. Neita, com. pers.), hasta que en los estudios del Proyecto Insectos de Suelo del Centro Internacional de Agricultura Tropical (CIAT; municipio de Palmira - suroccidente colombiano), se la encontró en gran abundancia alimentándose de raíces de diversos cultivos en la región.
Esta plaga subterránea se maneja mediante métodos tradicionales; el agricultor recurre al control químico, sin obtener resultados positivos, lo cual lo lleva a la sobredosis y el uso irracional de los insecticidas, generando en su lugar el exterminio de controladores naturales además del incremento de la contaminación de suelos y aguas subterráneas así como la pérdida de la calidad del suelo (Restrepo-Giraldo y LópezÁvila 2000).
Manejar la fecha de siembra distante de la aparición de la larva de tercer instar podría convertirse en una medida de control cultural económico y de fácil manejo para los agricultores (Ortega 2005). Otra forma de manejo es el control biológico con microorganismos y los agentes más usados en la actualidad son los bacterios, hongos, virus y nematodos entomopatógenos, éstos son los más exitosos por su alta especificidad (Jackson 1993). Un ejemplo del uso de nematodos entomopatógenos, se dio con el aislamiento de Steinernemaglaseri (Steiner, 1929) encontrado en la chisa Popillia japonica Newman, 1841 en Nueva Jersey, es el primer reporte de uso de entomopatógenos para el control de plagas (Glaser 1932); sin embargo, el desconocimiento de la interacción de este nematodo con su bacteria simbionte produjo resultados desalentadores, resultando en la pérdida de interés. Pese a esto, los estudios se retomaron hacia los años ochentas cuando los aislamientos de Heterorhabditis spp. y S. glaseri se comportaban mejor que otros (S. feltiae Filipjev, 1934 y S. carpocapsae (Weiser, 1955)). Los noventas se caracterizaron por estudios más profundos, donde se investigaba la causa de la resistencia de las chisas frente a los nematodos, además de hallar avances en producción y tecnología de estos últimos. Todo esto hizo más fácil su implementación dentro de programas de control biológico para diferentes especies de chisas (Klein 1990; Georgis y Gaugler 1991; Gaugler et al. 1992; Klein 1993).
Estudios realizados por diferentes autores como Villani y Wright (1988), Forschler y Gardner (1991), Klein (1992), Lacey et al. (1993), Gaugler et al. (1994) con diversas especies de chisas controladas con diferentes especies de NEPs (nematodos entomopatógenos), descubrieron que su uso es promisorio y su efectividad es mayor cuando las dosis de NEPs usadas son altas; además por la capacidad de reproducción y supervivencia de los nematodos es posible reducir el número de aplicaciones y eliminar el uso de algunos insecticidas.
La presente investigación, se estableció para buscar una alternativa viable para el control de la chisa rizófaga, P. bicolor, evaluando la infectividad y mortalidad de esta plaga con diferentes aislamientos nativos e introducidos de nematodos entomopatógenos.
Materiales y Métodos
Este experimento se llevó a cabo en condiciones de laboratorio en el campo experimental del CIAT, Valle del Cauca, Colombia, en un ambiente con 23 ± 2°C y 70 ± 5% de H.R.
Especie objetivo. Se trabajó con la chisa plaga P. bicolor, obtenida a partir de capturas en la región de Caldono del departamento del Cauca (1570 msnm; 18°C). Se seleccionó para este experimento el tercer instar. El efecto de los NEPs sobre este estadio en sus diferentes grados de madurez se evaluó con un mes de diferencia (tercer estadio inicial, intermedio, y final cercano a prepupa). Previamente estas larvas se mantuvieron en cuarentena en el laboratorio para descartar cualquier enfermedad adquirida en campo. La cría en laboratorio se mantuvo en un cuarto de crecimiento acondicionado (19°C y 70 ± 5% de H.R.), dentro de vasos plásticos con tapa (volumen de 100 ml), con arena y suelo orgánico estéril (1:3) alimentadas con trozos de zanahoria (3,5 g).
Nematodos. Se evaluaron nueve aislamientos de NEPs nativos e introducidos (Tabla 1), éstos se multiplicaron para cada experimento en larvas de último ínstar de Galleria mellonella (Linnaeus, 1758) (Lepidoptera: Pyralidae), obteniéndose juveniles infectivos (JI); éstos se mantuvieron en agua destilada estéril (ADE) a 15°C. Este procedimiento se hizo una semana antes de la aplicación.
Diseño Experimental. Se usó un Diseño Completamente al Azar (DCA) con cuatro repeticiones, con arreglo factorial,donde los factores fueron los aislamientos de NEPs y la edad de la larva tres. El testigo absoluto solo recibió agua destilada estéril. Se trabajó con 12 individuos. La dosis empleada fue de 10.000 JI/larva. Las evaluaciones se realizaron 10 días después de la infección. En el primer experimento se consideraron los aislamientos como un factor y en el segundo y tercer experimento los factores fueron los aislamientos de NEPs y la edad del tercer ínstar.
Inicialmente se evaluaron nueve aislamientos. Estos comprendían aislamientos nativos e introducidos de diversas especies. Posteriormente se evaluaron siete de estos y finalmente, se hizo una evaluación con cinco asilamientos, seleccionándolos del segundo y tercer experimento por ocasionar la mayor mortalidad en las pruebas previas. Además, por su amplio uso para el manejo de diversidad de plagas, S. carpocapsae (Sc) se utilizó para el segundo experimento. El aislamiento S. feltiae de Colombia (Sf1) se seleccionó en el segundo y tercer experimento, por ser nativo, por lo cual se considera importante compararlo con las especies de H. bacteriophora Poinar, 1975 (Hb) nativas e introducidas. S. riobravis Cabanillas, Poinar y Raulston, 1994 proveniente de Estados Unidos (Sr) se seleccionó por ser una especie usada comercialmente en el mundo. Además, estas especies presentaron la mejor reproducción in vivo durante este estudio, característica importante para un controlador. La evaluación en diferentes edades se realizó para determinar el efecto de la madurez de las larvas tres sobre la patogenicidad de los NEPs.
Se evaluaron las variables infección y mortalidad, para ambos casos se disectaron larvas vivas y muertas del insecto para verificar la presencia de NEPs en su interior. Se consideró que hubo infección para las larvas vivas o muertas si se encontraron nematodos en su interior; y se consideró mortalidad por nematodos cuando el nematodo se encontraba dentro de la larva muerta. Durante el experimento no se encontraron larvas muertas que no estuvieran parasitadas por nematodos.
La unidad experimental consistió en un vaso plástico de 56 ml con tapa, con suelo-arena estéril en proporción (3:1), donde se aplicó 1 ml de la suspensión de nematodos y 3 ml de agua destilada estéril (ADE) para ajustar la humedad a capacidad de campo. En el experimento se aplicaron los NEPs un día después de colocar la larva con un pedazo de zanahoria (3,5 g) como alimento. Las unidades así constituidas se mantuvieron en un cuarto de crecimiento en condiciones controladas (23 ± 2°C y 70 ± 5% de H.R).
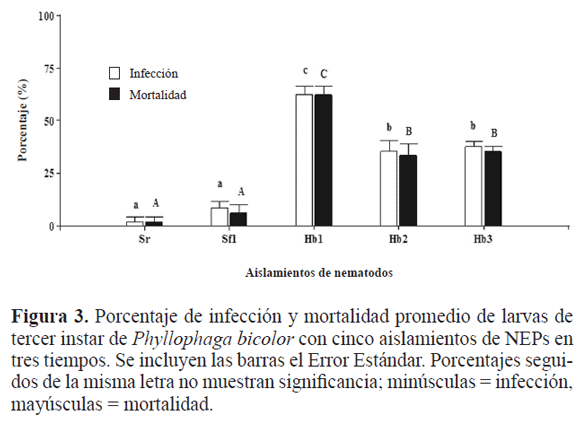
Análisis Estadístico. Se hizo un análisis de varianza con separación de medias para las variables significativas (Tukey P ≤ 0.05) con ayuda del paquete estadístico Infostat (2009), con una transformación previa utilizando la fórmula 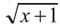 para corregir el efecto de valores cero en los datos y facilitar el análisis estadístico y la expresión del efecto de los tratamientos. Para las gráficas se utilizaron los datos en porcentaje, considerando el promedio de las cuatro repeticiones; la variabilidad de la distribución (como medida de dispersión) respecto de su media, se calculó tomando las desviaciones al cuadrado (varianza).
para corregir el efecto de valores cero en los datos y facilitar el análisis estadístico y la expresión del efecto de los tratamientos. Para las gráficas se utilizaron los datos en porcentaje, considerando el promedio de las cuatro repeticiones; la variabilidad de la distribución (como medida de dispersión) respecto de su media, se calculó tomando las desviaciones al cuadrado (varianza).
Resultados y Discusión
En el primer experimento se encontró el mayor rango de infección para H. bacteriophora: Hb2 (CENICAFÉ) y Hb1 (Italia) (F8,24 = 28,16; P ≤ 0,0001) y S. feltiae (Sf1) (Fig. 1). Las infecciones más bajas corresponden a las especies de Steinernema: SN1; Sr; Sa; Sc; Sf2. La mortalidad (F8,24 = 49,45; P ≤ 0,0001) fue mayor con el aislamiento nativo de CENICAFÉ (Hb2) (Fig. 1). Según Klein (1990, 1993) generalmente el complejo de especies de Heterorhabditis resulta más virulento que los de Steinernema; sin embargo, en una evaluación hecha sobre P. bicolor frente a la especie S. scarabaei (Stock y Koppenhöfer, 2003), aplicándole concentraciones menores, se observaron altas mortalidades. Esta nueva especie de Steinernemaha mostrado ser promisoria para el manejo de diversas especies de chisas (Koppenhöfer y Fuzy 2004; Melo et al. 2006).
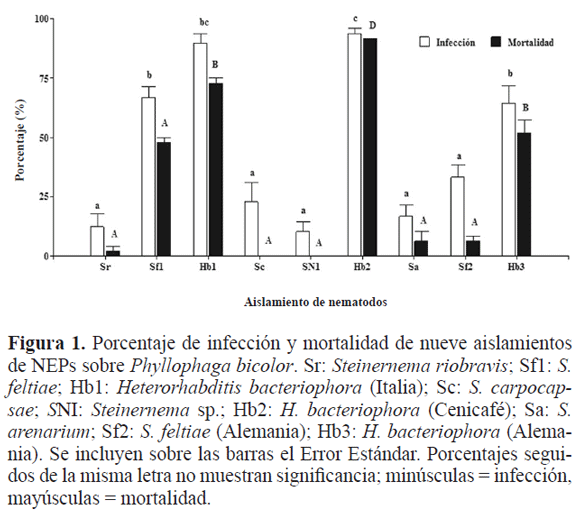
En el segundo experimento donde se evaluaron siete aislamientos, los NEPs del género Heterorhabditis fueron los más infectivos: Hb3; Hb1 y Hb2 (F6,18 = 21,26; P < 0,0001), y los que produjeron mayor mortalidad fueron Hb3; Hb2; Hb1 (F6,18 = 23,01; P < 0,0001) (Fig. 2). Además al comparar el efecto del tiempo sobre estas dos variables, se observa que en la segunda evaluación los valores bajaron significativamente así, la infección pasó de 52,38% a 33,63%; (F1,48 = 13,51; P < 0,0001); la mortalidad pasó de 38,99% a 26,49%, (F6,48= 7,15; P < 0,0102); del mismo modo, el comportamiento de los aislamientos en estos dos tiempos para las chisas de tercerinstar, Hb, tanto en infección (x = 73,95%) (F6,48 = 18,77; P < 0,0001) como en mortalidad (x = 64,58%) (F6,18 = 33,47; P < 0,0001) fueron más patogénicos que los de Steinernema(x Inf = 19,8%; x Mort: 8,85%). Esto es coherente con lo hallado en los estudios de Melo et al. (2007) y Koppenhöfer y Fuzy (2004), donde un incremento en la edad de las chisas P. menetriesi, Anomala inconstans Burmeister, 1844, P. japonica y A. orientalis Waterhouse, 1875, disminuyó la efectividad de H. bacteriophora.
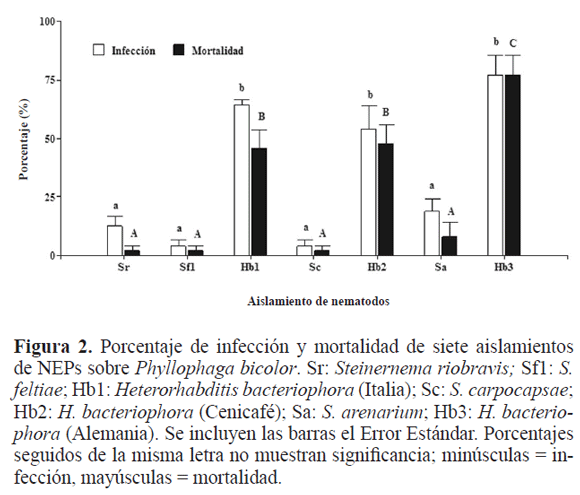
Al analizar con cinco aislamientos de NEPs, frente a la edad de la larva tres se obtuvieron datos constantes a través del tiempo, al presentarse disminución de la infección y mortalidad con la madurez de ésta (Inf.: de 65,42% a 29,17%; Mort.: de 53,33% a 27,92%). Los aislamientos de Heterorhabditis se destacan frente a los otros en infección (F4,12 = 31,26; P ≤ 0,0001). Los resultados fueron similares para la mortalidad (F4,12 = 27,44; P ≤ 0,0001) (Fig. 3).
Finalmente se comparó la infección (F2,45 = 40,60; P ≤ 0,0001) y mortalidad en las tres edades (F2,45 = 29,26; P = 0,0001) (Fig. 4), los mayores valores se obtuvieron para las larvas más jóvenes siendo ésta la mejor edad para controlar la plaga con los NEPs; en la interacción de estos dos factores en estudio, edad y aislamientos, las cepas más infectivas fueron las de H. bacteriophora presentándose datos tan contrastantes como 93,7% para Hb2 en las larvas más jóvenes, seguidos de Hb1 en la misma edad y por Hb3 en la edad intermedia frente a Sf1 (4,14% y 8,33% en la edad más avanzada y la intermedia, respectivamente) (F8,45 = 7,64; P ≤ 0,0001) al igual que en la infección, la que produjo más mortalidad fue la de Hb2 en las larvas más jóvenes (91,7%) y la de menor mortalidad fue Sf1 (2,1%) en la edad intermedia, compartiendo rango con Sr para las tres edades (F8,45 = 11,13; P ≤ 0,0001) el aislamiento nativo Hb2 mostró ser más efectivo para controlar esta plaga.
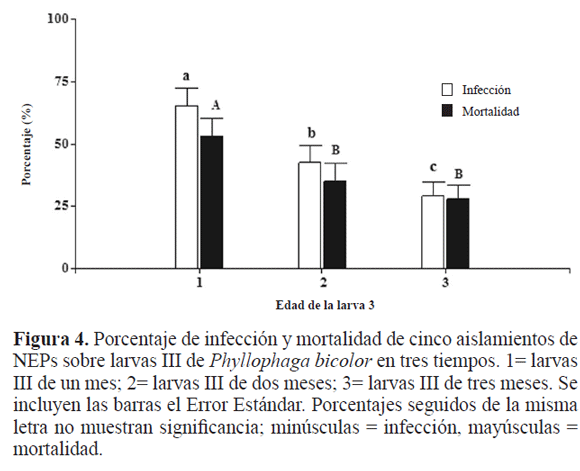
Se observa que a medida que la larva del tercer instar aumenta la edad, la infección y mortalidad disminuyen, sin embargo los nematodos de la especie H. bacteriophora, muestran ser más efectivos lo que puede deberse, entre muchas causas, a que posee un diente que le facilita el ingreso por cualquier parte de la cutícula de la chisa sin depender el ingreso exclusivamente de sus orificios naturales, como espiráculos, boca y ano; no obstante, esta característica podría enfrentarlo eventualmente a dificultades como la reacción inmunológica a los nematodos. Los resultados soportan el supuesto de que las larvas con la edad se hacen más fuertesal ataque de los NEPs. En experimentos hechos por Melo et al. (2006) se observó que las prepupas eran más resistentes al efecto de los nematodos que larvas más jóvenes y Koppenhöfer y Fuzy (2004) encontraron en otros géneros de chisas el mismo comportamiento, aunque resultados específicos sobre el efecto de los nematodos sobre el mismo ínstar a diferentes edades no se han reportado antes. Algunos de los aspectos que pueden estar influyendo en el efecto de los NEPs es el hecho de poseer placas sobre los espiráculos que impiden la penetración del patógeno; la frecuente defecación, en la que se expulsa a los nematodos; el poseer un comportamiento defensivo y evasivo, en los cuales rompen los nematodos con las mandíbulas, además que se desplazan rápidamente cuando se ven atacados; una fuerte respuesta inmune asociada a la edad, donde estados más desarrollados pueden eliminar patógenos invasores, y por último la cutícula en Larva III es muy gruesa lo cual dificulta ser penetrada (Kaya 1985; Jackson y Brooks 1995; Koppenhöfer et al. 2004).
Como otros insectos del suelo, las chisas están expuestas a ser atacadas por los nematodos entomopatógenos y muchos de éstos han sido aislados naturalmente de estas larvas. Sin embargo al coevolucionar con estos patógenos, las chisas han desarrollado varios mecanismos de resistencia en su comportamiento y una fuerte respuesta inmune (Klein 1990, 1993; Peters 1996; Shapiro-Ilan et al. 2002). Adicionalmente, la efectividad de los entomopatógenos sobre los insectos está sujeta a su interacción con el ambiente y su variación, lo que también entra a jugar un papel importante en esta relación (Shannon 1994; Shannon y Carballo 1996).
Agradecimientos
Este trabajo fue financiado por el Ministerio de Cooperación Económica (BMZ) de Alemania y el Centro Internacional de Agricultura Tropical (CIAT). Los autores agradecen a Germán Andrés Calberto, Rodrigo Zúñiga, Oscar Yela y Rómulo Riascos por el suministro y mantenimiento del material insectil.
Literatura citada
CIAT. 2004. Soil pests-Cassava and other crops, pp. 116-165. En: Centro Internacional de Agricultura Tropical (CIAT). Annual Report. Integrated pest and disease management in major agroecosystems. Cali, Colombia. 417 p. [ Links ]
FORSCHLER, B. T.; GARDNER, W. A. 1991. Concentration-mortality response of Phyllophaga hirticula (Coleoptera: Scarabaeidae) to three entomogenous nematodes. Journal of Economic Entomology 84 (3): 841-843. [ Links ]
GAUGLER, R.; CAMPBELL, J. F.; SELVAN, S.; LEWIS, E. E. 1992. Large-scale inoculative releases of entomopathogenic nematode Steinernema glaseri: assessment 50 years later. Biological Control 2: 181-187. [ Links ]
GAUGLER, R.; WANG, Y.; CAMPBELL, J. F. 1994. Aggressive and evasive behaviours in Popillia japonica (Coleoptera: Scarabaeidae) larvae: defences against entomopathogenic nematode attack. Journal of Invertebrate Pathology 64: 193-199. [ Links ]
GEORGIS, R.; GAUGLER, R. 1991. Predictability in biological control using entomopathogenic nematodes. Journal of Economic Entomology 84: 713-720. [ Links ]
GLASER, R. 1932. Studies on Neoaplectana glaseri, a nematode parasite of the Japanese beetle (Popillia japonica). New Jersey Department of Agriculture. Circular 211. Trenton, NJ. 34 p. [ Links ]
INFOSTAT. 2009. Manual del usuario. Grupo INFOSTAT, FCA., Universidad Nacional de Córdoba. 1 Ed. Editorial Brujas, Argentina. 318 p. [ Links ]
JACKSON, T. A. 1993. Developing microbial controls for scarab pest. En: Diversidad y manejo de plagas subterráneas. Memorias de la IV mesa redonda sobre plagas subterráneas 14 y 15 de Octubre, Instituto de Ecología, AC. Xalapa, Veracruz, México. Sociedad Mexicana de Entomología, AC. Instituto de Ecología. AC. 261 p. [ Links ]
JACKSON, J. J.; BROOKS, M. A. 1995. Parasitism of western corn rootworm larvae and pupae by Steinernema carpocapsae. Journal of Nematology 27 (1): 15-20. [ Links ]
KAYA, H. K. 1985. Susceptibility of early larval stages of Pseudalletia unipuncia and Spodoptera exigua (Lepidoptera: Noctuidae) to the entomogenous nematode Steinernema feltiae (Rhabditida: Steinernematidae). Journal of Invertebrate Pathology 46: 58-62. [ Links ]
KLEIN, M. G. 1990. Efficacy against soil-inhabiting insects pests, pp. 195-214. En: Gaugler, R.; Kaya, H. K. (Eds.), Entomopathogenic nematodes in biological control. CRC. Press, Boca Raton, Florida. 365 p. [ Links ]
KLEIN, M. G. 1992. Use of Bacillus popilliae in Japanese beetle control, pp. 179-190. En: Jackson, T. A.; Glare, T. R. (Eds.). Use of pathogens in scarab pest management. Intercept Andover. Hampshire, England. 298 p. [ Links ]
KLEIN, M. G. 1993. Biological control of scarabs with entomopathogenic nematodes, pp. 49-57. En: Bedding, R.; Akhurst, R.; Kaya, H. (Eds.). Nematodes and the biological control of insects. CSRIO, East Melbourn. [ Links ]
KOPPENHÖFER, A. M.; FUZY, E. M. 2004. Effect of white grubs developmental stage on susceptibility to entomopathogenic nematodes. Journal of the Entomological Society of America 97 (6): 1842-1849. [ Links ]
KOPPENHÖFFER, A. M.; FUZY, E. M.; CROCKER, R. L.; GELERNTER, W. D.; POLAVAPARU, S. 2004. Pathogenicity of Heterorhabditis bacteriophora, Steinernema glaseri, and S. scarabaei (Rhabditida: Heterorhabditidae, Steinernematidae) against 12 white grubs species (Coleoptera: Scarabaeidae). Biocontrol Science and Technology 14 (1): 87-92. [ Links ]
LACEY, L. A.; BETTENCOURT, R.; GARRETT, F. J.; SIMÕES, N. J.; GAUGLER, R. H. 1993. Factors influencing parasitism of adult Japanese beetles, Popillia japonica (Col.: Scarabaeidae) by entomopathogenic nematodes. Entomophaga 38 (4): 501- 509. [ Links ]
LONDOÑO, M. 2001. Las chizas y su manejo, hortalizas, plagas y enfermedades. Corpoica-Sociedad Colombiana de Entomología. Rionegro. p. 36-47. [ Links ]
MELO, E. L.; ORTEGA, C. A.; GAIGL, A.; KOPPENHÖFER, A.; BELLOTTI, A. C. 2006. Evaluación de patogenicidad e infección de la nueva especie Steinernema scarabaei Stock & Koppenhöfer (Rhabditida: Steinernematidae), sobre la chisa rizófaga Phyllophaga pos. Bicolor, pp. 127-136. En: Parada, S. J.C.; Luque, J. E.; Piedrahita, C. W. de J. (Eds.). Nematodos entomoparásitos: experiencias y perspectivas. Editorial Unibiblos. 193 p. [ Links ]
MELO, E. L.; ORTEGA-OJEDA, C. A.; GAIGL, A. 2007. Efecto de nematodos sobre larvas de Phyllophaga menetriesi y Anomala inconstans (Coleoptera: Melolonthidae). Revista Colombiana de Entomología 33(1): 21-26. [ Links ]
ORTEGA, C. A. 2005. Estudios metodológicos para evaluar el impacto económico de escarabajos Melolonthidae (Insecta: Coleoptera) en tres cultivos tropicales. Trabajo de grado de Master en Ciencias en Fitoprotección. Escuela Politécnica del Ejército. Quito, Ecuador. 109 p. [ Links ]
PARDO-LOCARNO, L. C. 2002. Aspectos sistemáticos y bioecológicos del complejo chisa (Col, Melolonthidae) de Caldono, Norte del Cauca, Colombia. Trabajo de grado de Maestría en Biología. Universidad del Valle, Facultad de Ciencias, Santiago de Cali. 145 p. Nematodos para control de chisa 212 [ Links ]
PETERS, A. 1996. The natural host range of Steinernema and Heterorhabditis spp. and their impact on insects populations. Biocontrol Science and Technology 6: 389-402. [ Links ]
POSADA, O. L. 1993. Las chisas, sus enemigos naturales y recomendaciones sobre su manejo. Agricultura Tropical 30 (3): 71- 79. [ Links ]
RESTREPO-GIRALDO, H.; LÓPEZ-ÁVILA, A. 2000. Especies de chisas (Coleoptera: Melolonthidae) de importancia agrícola en Colombia. Corporación Colombiana de Investigación Agropecuaria. Programa Nacional de Manejo Integrado de Plagas. 62 p. [ Links ]
SHANNON, P. J. 1994. Control microbiano de Phyllophaga spp. (Col: Melolonthidae),pp. 80-92. En: Memoria Seminario - Taller sobre la biología y control de Phyllophaga spp. Informe Técnico No. 277. CATIE. Turrialba, Costa Rica. [ Links ]
SHANNON, P.; CARBALLO, M. 1996. Biología y control de Phyllophaga spp., pp. 80-93. En: Seminario-Taller Centroamericano sobre la biología y control de Phyllophaga spp. Serie Técnica. Informe Técnico 227. CATIE, Turrialba, Costa Rica. [ Links ]
SHAPIRO-ILAN, D. I.; GOUGE, D. H.; KOPPENHÖFER, A. M. 2002. Factors affecting commercial success: case studies in cotton, turf and citrus, pp. 333-355. En: Gaugler, R. (Ed.). Entomopathogenic nematology. CAB International. Wallingford. [ Links ]
VILLANI, M. G.; WRIGHT, R. J. 1988. Entomogenous nematodes as biological control agents of European chafer and Japanese beetle (Coleoptera: Scarabaeidae) larvae infesting turfgrass. Journal of Economic Entomology 81 (2): 484-487. [ Links ]

