










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488On-line version ISSN 2665-4385
Rev. Colomb. Entomol. vol.36 no.2 Bogotá July/Dec. 2010
Soil spiders in differing environments: Eucalyptus plantations and grasslands in the Pampa biome, southern Brazil
Arañas del suelo en ambientes diferentes: Plantaciones de Eucalyptus y pasturas en el bioma de Pampa, sur de Brasil
EVERTON NEI LOPES RODRIGUES1,2, MILTON DE S. MENDONÇA, JR.1, JOÃO L. O. ROSADO3 and ALCI E. LOECK3
1 Programa de Pós-Graduação em Biologia Animal, Departamento de Zoologia, Instituto de Biociências, Universidade Federal do Rio Grande do Sul. Av. Bento Gonçalves, 9500, Bloco IV, Prédio 43435, 91501-970 Porto Alegre, RS, Brazil. enlrodrigues@yahoo.com.br Corresponding author.
2 Museu de Ciências Naturais, Fundação Zoobotânica do Rio Grande do Sul. Rua Dr. Salvador França, 1427, 90690-000 Porto Alegre, RS, Brazil.
3 Programa de Pós-Graduaçao em Fitossanidade, Departamento de Fitossanidade, Faculdade de Agronomia Eliseu Maciel, Universidade Federal de Pelotas. Campus Universitário, s/nº, caixa postal 354, 96010-900 Pelotas, RS, Brazil.
Recibido: 20-mar-2010 • Aceptado: 6-nov-2010
Abstract: The Pampa biome in southern Brazil has grassland areas with high biodiversity. Given the fast advance of Eucalyptus silviculture over grassland areas in this biome, and the scant knowledge on the soil spider fauna in these landscapes, we aimed to provide a first view of spiders occurring there and to quantitatively evaluate differences in the spider fauna between the two environments. Study areas included five farms each with the two environments, native grassland and Eucalyptus plantation. Spider densities were 6.53 (±1.01s.e.) individuals/m-2 in silviculture and 3.88 individuals/m-2 (±0.73) in grassland. This could be due to spiders finding more shelter sites underneath a denser silviculture litter than in the grassland where they could be more exposed, for example, to their own predators. Twentyfour spider families were captured; the most abundant and diverse were Salticidae and Linyphiidae. Nineteen families occurred in the silviculture and 21 in the grassland. For adult spiders, 51 morphospecies were determined, the most abundant being Guaraniella mahnerti. Adult abundance was marginally significant for environment, with silviculture areas having more spiders. Species density did not differ between environments or sites, but evenness was significantly higher for the grassland. This better balance in species abundances for spider assemblages in grasslands suggests a healthier environment compared to a monoculture. The most abundant guild was that of the running hunters. Even as a rapid spider diversity inventory, the information gathered here adds considerably to our knowledge on how this new economic upsurge in silviculture affects native environments.
Key words: Araneae. Monoculture. Richness. Rio Grande do Sul.
Resumen: El bioma de la Pampa en el sur de Brasil tiene pasturas con alta biodiversidad. Dado el rápido crecimiento del cultivo de Eucalyptus sobre estas áreas de pasturas y el poco conocimiento de la fauna de arañas de suelo, se planteó hacer un reconocimiento de las arañas que ocurren allí y una evaluación cuantitativa de las diferencias en esta fauna entre los dos ambientes. Las áreas de estudio incluyeron cinco fincas cada una con los dos ambientes, pasturas nativas y plantaciones de Eucalyptus. La densidad de arañas fue de 6,53 (±1,01d.e.) individuos/m-2 en el silvicultivo y de 3,88 individuos/m-2 (±0,73) en las pasturas. Este resultado podría deberse a que las arañas encontraron más refugios bajo una capa de hojarasca más densa en el silvicultivo que en las pasturas donde pueden estar más expuestas entre otras cosas a sus depredadores. Se capturaron 24 familias de arañas, las más abundantes y diversas fueron Salticidae y Linyphiidae. Diecinueve familias se encontraron en el silvicultivo y 21 en las pasturas. Se identificaron 51 morfoespecies de arañas adultos, la más abundante es Guaraniella mahnerti. La abundancia de adultos fue marginalmente mayor en las áreas de silvicultivo. La densidad de especies no fue diferente entre ambientes o sitios, pero la equitatividad fue significativamente mayor para las pasturas. Este mayor balance en las abundancias de especies de los ensamblajes de arañas en las pasturas sugiere un ambiente más sano comparado con el de un monocultivo. El gremio más abundante fue el de las cazadoras cursoriales. Aun siendo un inventario rápido de la diversidad de arañas, la información obtenida aquí es un aporte significativo a nuestro conocimiento de cómo esta nueva tendencia económica en silvicultura afecta los ambientes nativos.
Palabras clave: Araneae. Monocultivo. Riqueza. Rio Grande do Sul.
Introduction
The Pampa biome has vast areas of grasslands, covering the southern half of Rio Grande do Sul state, already the southernmost state of Brazil, filling approximately 176.496 km2 (IBGE 2004). Southern grasslands include areas with a high diversity of both plants and animals, with a long history of low impact pasture management under extensive cattle farming. However, its conservation has been threatened by the increased degradation caused by the inadequate use of exotic species, especially grasses, and especially by a recent economic interest in agriculture and silviculture (Pillar et al. 2009).
Land use (agriculture, silviculture and pasture) can degrade natural environments, reducing biodiversity, mainly through loss of habitat (Wilcox and Murphy 1985; Primack and Rodrigues 2002). The establishment of exotic species silviculture has been a widely debated subject in southern Brazil, an activity that is known to generate strong negative impact in the original environments (Pillar et al. 2009). Substitution of native areas for tree monocultures may lead to continuous and irreversible biodiversity loss, either directly through species extinction, or through habitat fragmentation. Some studies suggest planted forests have also a lower productivity than most local natural habitats (Lima 1993; Bird et al. 2004).
Many soil organisms, as most invertebrates, are directly affected by land use mode. Arthropods are an important component of the natural diversity in any habitat (May 1986), including native grasslands and silviculture. On the soil ofboth environments a complex system of organic matter cycling is established and leaf litter acts as food for a variety of arthropods, and as shelter for others, composing a particular food chain of detritivores and predators (Höfer et al. 1996). Among soil arthropods, spiders stand out as important predators (Nyffeler et al. 1994; Foelix 1996; Beck et al. 1997), with a role in checking the balance of such edaphic/litter communities (Bultman and Uetz 1982; Toti et al. 2000).
Few literature sources have studied the edaphic fauna of the eucalypt monoculture, comparing it to other environments, and mostly such comparisons address native forests (Ferreira and Marques 1998; Pellens and Garay 2000; Moço et al. 2005; Lo-Man-Hung et al. 2008). Besides, usually species lists of groups as Araneae are not provided, with the exception of Rinaldi (2005) recording the spider fauna of a silviculture area.
Evaluations of diversity, richness and invertebrate species composition, especially arthropods, can help understand the dynamics of these introduced habitats. It is thus fundamental for wide scale planning of environmental management and conservation of all habitats composing a landscape. Given the advance of eucalypt silviculture activities over the grassland ecosystem of the Pampa biome, we aimed to evaluate the abundance, species richness, and foraging guild proportions and species composition for soil spiders comparing both environments.
Material and Methods
Study areas. Sampling took place in three municipalities of the southern region of Rio Grande do Sul state of Brazil, within the Pampa biome. Five farms were sampled in the summer of 2008: in Cerrito municipality, Nossa Senhora do Guadalupe (NG) farm (31°77’56.57”S, 52°64’53.32”W; 208 ha of planted area; 440 ha of total area, sampled in 03/01/2008) and Pitangueiras (PI) farm (31°79’38.57”S, 52°53’50.51”W; 101 ha of planted area; 230 ha of total area, sampled in 24/01/2008); in Capão do Leão municipality, Ouro Verde (OV) farm (31°57’55.40”S, 52°51’42.73”W; 125 ha of planted area; 297 ha of total area, sampled in 15/01/2008); in Piratini municipality, Santa Izabel (SI) farm (31°56’50.95”S, 52°88’20.48”W; 145 ha of planted area; 330 ha of total area, sampled in 14/02/2008) and Santa Maria (SM) farm (31°56’58.01”S, 53°17’06.13”W; sampled in 21/02/2008). Silviculture was based on Eucalyptus saligna Smith planted in 2006. Grassland areas are managed lightly by releasing cattle infrequently for use as pasture.
Sampling method and design. For each farm a native grassland and eucalypt plantation area were designated. From each area 25 samples were taken, distributed along a linear transect. To avoid edge effects, samples were at least 50 m far from the limits between the areas, and to guarantee a degree of independence between samples, there were also at least 50 m between consecutive sampling areas. For each sample all leaf litter and a small superficial fraction of the soil was taken from an area of 1 m2. The material was put through a field sieve, with the content transferred to closed nylon bags. These were taken to the lab where bag content was placed in Winkler extractors, where it stayed for 72h to collect the animals. Spiders were identified in the Laboratório de Aracnologia and deposited in the spider collection (curator: E. H. Buckup) of Museu de Ciências Naturais of Fundação Zoobotânica do Rio Grande do Sul, in Porto Alegre, Brazil.
Data analysis. Diversity variables were compared using PASt (Paleontological Statistics 1.97, Hammer and Harper 2009), with environment (eucalypt plantation or native grassland) and site (farms) as factors. Alpha diversity (abundance, richness and equability) was compared at the family and species levels using two-way ANOVAs. To illustrate spider species composition comparisons two ordenations were plotted (Non-metric MultiDimensional Scaling - nMDS) using a qualitative similarity index (Simpson) and a quantitative one (Morisita). To test for statistical differences among spider assemblage composition for the above similarity indexes, we applied two one-way ANOSIM (Analysis of Similarities) with Bonferroni correction, one for each factor. A SIMPER Analysis (Percentage Similarity) was employed to rank species contributing more for dissimilarities among environments and sites (Clarke and Warwick 1994). Species accumulation curves and analytical species richness estimator Chao1 were calculated to verify sampling sufficiency, as suggested by Toti et al. (2000). We used EstimateS 8.0 (Colwell 2005) with 500 randomizations.
Guild classification was based on Uetz et al. (1999), Höfer and Brescovit (2001) and Rodrigues et al. (2009), where all captured spiders, separated by family, were grouped as weavers, divided between orb weavers - ORB (build bidimensional webs) and space web sheet builders - SPW (build tridimensional webs) or as hunters, divided between hunting runners - HRU (search actively for prey) and hunting ambushers/ stalkers - HAS (do not build webs but sit-and-wait for prey). Comparisons of guild proportions among environments and sites used a two-way ANOVA (with data arcsine transformed).
Results
Overall 1.301 spiders were found, mostly immatures (76.4%). Among adults, females were more common (77.5%) than males (22.5%) (a sex ratio of 1:3.5). A higher absolute abundance characterised the eucalypt plantation with 816 individuals, whilst grassland resulted in 485 spiders. Thus, spider densities were 6.53 (± 1.010 s.e.) individuals.m-2 in eucalypt, and 3.88 individuals.m-2 (± 0.731 s.e.) in grassland. Out of 125 samples, seven did not record spiders in eucalypts and 21 in the grassland; the highest number of spiders in a sample was 36 individuals for eucalypt and 27 for grassland.
Twenty-four spider families were captured, of which eleven were represented only by juveniles; the most abundant were: Salticidae (N = 276), Linyphiidae (230), Gnaphosidae (185), Theridiidae (124) and Lycosidae (116); the least abundant were Nemesiidae, Oonopidae, Senoculidae and Sparassidae, all with singletons. Of the five most abundant families, four were more abundant in eucalypt, the exception being Lycosidae, found more in the grasslands (Table 1). Nineteen families occurred in eucalypt and 21 in grassland. Three were exclusive to eucalypt, five to grassland. Six families were recorded from all sites and both environments (Table 1).
Analyses at the familial level (i.e. including young spiders) showed average abundance to differ significantly between environments, with more individuals found in eucalypt plantations (F1,4 = 13.83, P = 0.020). Surprisingly, differences in family density among sites are marginally significant (F4,4 = 6.00, P = 0.055; Fig. 1), indicating a possible founder effect. No differences in evenness among families were detected between environments or among sites.
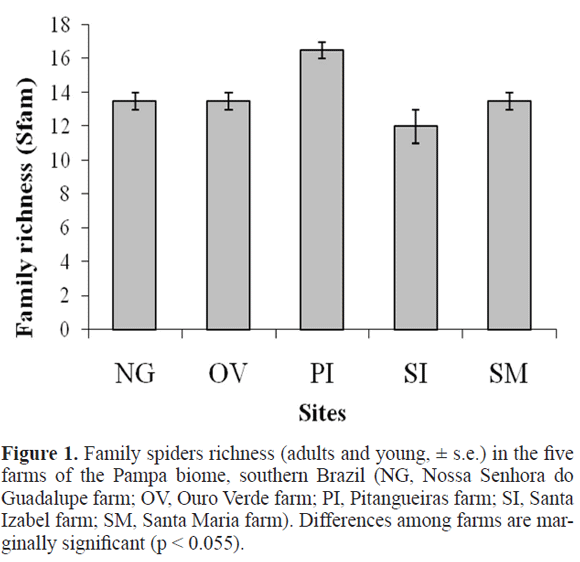
For adult spiders, 51 morphospecies were determined, the most abundant being Guaraniella mahnerti Baert, 1984 (N = 42), Thymoites sp. (34), Smermisia vicosana (Bishop and Crosby, 1938) (24) and Hisukattus tristis (Mello-Leitão 1944) (23) (Table 2). Twenty morphospecies were singletons and eight were doubletons, comprising 55% of the sampled species (Table 2). The richest families in morphospecies were Salticidae (12), Linyphiidae (11) and Theridiidae (8), independently of environment. Thymoites sp. was the only morphospecies recorded from all sampled sites.
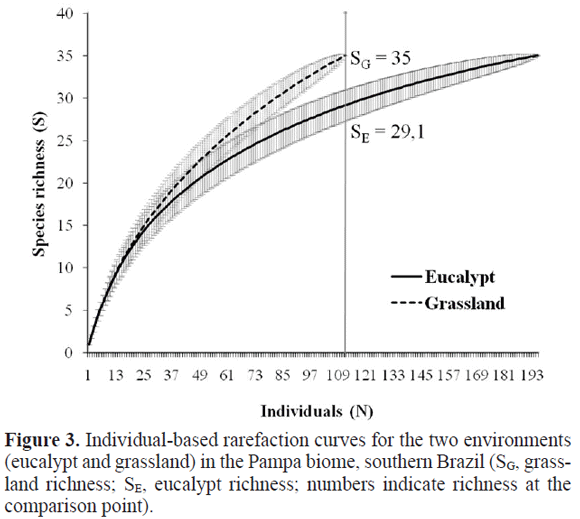
Each environment totalled 35 morphospecies, and thus no sample-based rarefaction was employed to distinguish environments. However, the Chao1 estimator indicated an expected 40.63 species for eucalypt, which would mean 86.1% of the spider fauna have been sampled for that environment. Figures for grassland were somewhat different with 47 species expected, meaning 74.5% of the fauna sampled. However, the estimated values do not differ significantly between environments (considering confidence intervals).
Adult abundance was marginally significant for environment (F1,4 = 7.50; P = 0.052), with eucalypt areas having more spiders (Fig. 2A); no differences were found among sites. Species density (Fig. 2B) did not differ between either environments or sites, but evenness was significant for environment (F1,4 = 14.65; P = 0.019), being higher in grassland (Fig. 2C). Consequently, abundance distribution curves showed higher dominance by a few species in eucalypt with more rare species in grasslands as well. Curves for both environments significantly approximated the log-series model (eucalypt: α = 12.42, x = 0.94, Χ2 = 2.66; P = 1.00; grassland: α =17.46, x = 0.86, Χ2 = 3.68; P = 0.99 Fig. 3). Higher abundance in eucalypt and identical species densities means significantly higher species richness for grasslands under individual-based rarefaction (Fig. 4).
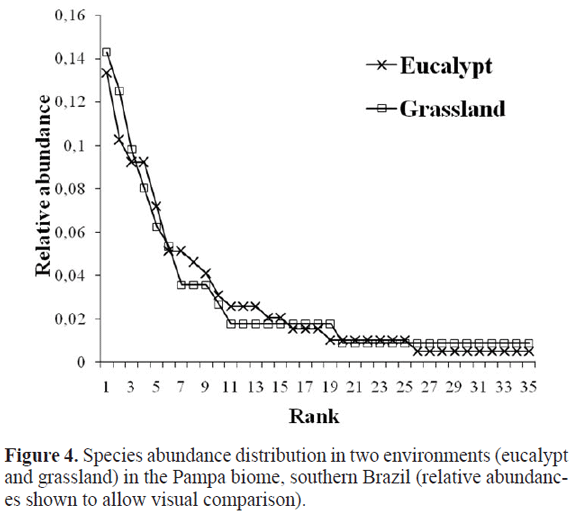
More than 37% of the species were common to both environments and the two most common species on each environment were the same. Sixteen species were exclusive to each environment, probably due to the large number of singletons and doubletons in the sample. This amount of exclusivity sets the two environments apart and thus differences in species composition are significant between the two environments for the qualitative index (Simpson, ANOSIM: R = 0.248; P = 0.049; this significance is just lost when singletons are excluded from the analysis). However, taking species abundance into consideration renders this difference insignificant, as the quantitative index shows (Morisita: R = - 0.116; P = 0.776). There is no apparent founder effect in terms of species composition, since no difference among sites was foundfor any index. Figure 5 illustrates species composition differences among environments and sites. The contribution of the most representative species in each environment to dissimilarity (SIMPER) between environments is presented in Table 3. Guaraniella mahnerti (percentage contribution: 7.75%) and Smermisia vicosana (6,85%) were the most important ones.
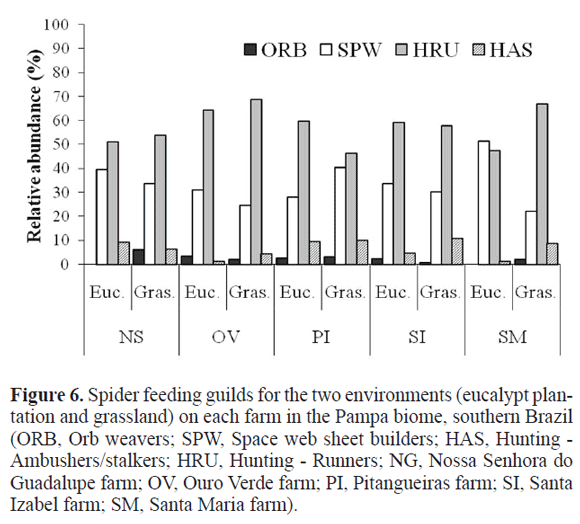
The most abundant guild was the running hunters (N = 743), followed by irregular web builders (454) ambush hunters (77) and especially orbicular web builders (27) were relatively rare (Fig. 6). No differences were found in the proportion represented by each guild, either between environments (two-way ANOVA, interaction term: F3,32 = 1.547 p = 0.221) or sites (two-way ANOVA, interaction term: F12,20 = 0.789 p = 0.656).
Discussion
This is the first record of the spider fauna comparing eucalypt silviculture and grasslands in the Pampa biome of Brazil. Most of the studies compare eucalypt plantation to native forests; among such comparisons, generalizing for soil arthropod fauna, a lower species richness and/or abundance is found in silviculture (Ferreira and Marques 1998; Pellens and Garay 2000; Moço et al. 2005; Lo-Man-Hung et al. 2008).
The above sources differ somewhat from what we found in southern Rio Grande do Sul; a higher abundance was found in a monoculture (eucalypt) and species density was identical between eucalypt and grassland, although richness per se was higher in the latter. These two environments differ in other aspects as well, with grasslands showing higher heterogeneity in their spider assemblages and species composition quantitatively distinct. Higher structural vegetation complexity is known to lead to higher spider diversity, thanks to a larger number of microhabitats for web building and shelter (Souza 2007; Rypstra et al. 1999). The only sense in which native grassland have higher heterogeneity compared to eucalypt silviculture is in terms of plant species richness, but its effect seem too indirect to explain patterns in spider distribution.
In the eucalypt plantation the amount of litter can be higher due to a larger plant biomass. Litter can soften extreme abiotic factors (Uetz 1979; Höfer et al. 1996), and in silviculture spiders could more easily find shelter and a substrate, whereas in the grassland they would be more exposed to predators. A low orgaismal diversity due to lower nutrient amount and cycling in the litter has been described for eucalypt plantations (Majer and Recher 1999), however, these factors do not seem to influence spider abundance given the versatility of such opportunistic and generalist organisms (Foelix 1996). Because the plantation areas studied here were recent, with time it may be expected that abundance and species density could even increase more, depending on litter dynamics.
At the litter level, the grassland environment also suffers stronger disturbances, as higher light intensity, higher wind speeds and direct rainfall, among others. Some or all of these abiotic factors could lead to a lower abundance in the grassland, either through stronger hygrothermal stress to spiders, damages to webs, or both. These effects in turn can influence not only spider survival and reproduction but also important behaviors as those involved in environment choice (Wise 1993).
Family composition showed dissimilarity with higher richness and more exclusive families recorded in the grassland.Rinaldi (2005) studied a eucalypt plantation in Botucatu, São Paulo state, Brazil, recording 17 families at different strata, finding only seven for soil; here we report 19. Six families were shared between theirs and our inventory, Caponiidae was exclusive to the Botucatu study and six were exclusive to southern Rio Grande do Sul. No Mygalomorphae was recorded for eucalypt silviculture in either inventory, but we recorded family Nemesiidae for grassland.
Comparing the araneofauna in this study to other work within the Pampa biome is all but impossible given the absence of publications with the same aim. In Rodrigues et al. (2009) we compared the araneofauna of a grassland environment similar to the one reported here against a rice agroecosystem, but at a smaller scale, using a fairly isolated area and a different method (sweeping net): 12 families were found in that case. We report Salticidae as the most abundant family, Rodrigues et al. (2009) recorded Oxyopidae as the most abundant in grasslands near rice.
Some species are worth mentioning for their habitat use. Teminius insularis (Lucas, 1857) was found here from both environments; Lo-Man-Hung et al. (2008) sampled it from pitfall-traps in eucalypt plantations from Amazonia, not finding it either in primary or secondary forests. Rinaldi (2005) recorded this species in edges and interior of eucalypt plantation using sweeping nets. Cheiracanthium inclusum Hentz, 1847 is a species commonly found in agroecosystems (Young and Edwards 1990), and here indeed it occurred only in the monoculture; Rinaldi (2005) recorded it from the edge between eucalypt plantation and in the grassland. Rodrigues et al. (2009) sampled Oxyopes salticus more from the grassland than in a rice agroecosystem, in here it occurred only in the grassland as well. These spiders could act in the biological control of pest insects in the monoculture as suggested by Rinaldi (2005), however, they may need the grassland as a refuge because of eucalypt silvicultural management practices acting as infrequent but strong environmental disturbances.
Spiders are usually among the first organisms to occupy altered or recently formed habitats, actively participating in community succession processes (Uetz et al. 1999). Grassland, as the natural environment usually surrounding the disturbed area, may serve as the origin for predators (among those the araneofauna) colonizing silviculture, a pattern already suggested by Plagens (1983). However, the low similarity between the faunas of the two environments suggests otherwise. Silviculture selectivity in regard to spider species seems to be high given the low degree of nesting of the two faunas. Possibly, rare spider species inhabiting grassland can readily adapt to silviculture, although it should be noted that plantation areas are not too long, and thus a longer time may influence the fauna in silviculture so as to change the observed patterns in still unpredictable ways. Clearly, long term monitoring of such faunas is necessary to better understand such processes.
Although it is known that distinct environments and microenvironments are selective in terms of spider hunting guilds as well as families and species (Uetz 1979; Uetz et al. 1999; Toti et al. 2000; Whitmore et al. 2002), guild composition too was very similar between environments, with cursorial hunters predominating. Rinaldi (2005) also recorded hunters as the most abundant guild in the eucalypt plantation soil. The nearly absence of a shrub layer in the eucalypt plantation and a predominance of herbs in the grassland means a lack of physical structures for orbicular web building, which would appear as tourists on the soil anyway. However, irregular web builders can be successful even without tridimensional structures appropriate for orbiculars, either in the grassland or plantation. This would explain the marked abundance of hunters and irregular web builders in the two environments.
Even a rapid spider diversity inventory can produce information on the main ecological aspects of such an assemblage. Data presented here can help direct future studies focusing on the changes brought to the grassland environment and the whole Pampa biome given the strong efforts to occupy these areas with exotic silviculture. Long term studies, especially, may permit solving the puzzles found here, to better understand the relationship between native and introduced environments, perhaps to help minimize the impacts brought by human disturbance.
Acknowledgements
To Erica H. Buckup (curator of the Araneae collection) and Maria A. L. Marques for access to spider material and bibliography and the direction of MCN by permitting the first author to develop part of this work at it dependences. This study was supported by CNPq (process 140586/2007-5 for E. N. L. Rodrigues from Programa de Pós-Graduação em Biologia Animal da Universidade Federal do Rio Grande do Sul).
Literatured cited
Beck , L.; Höfer , H.; Martius , C.; Römbke , J.; Ver haagh , M. 1997. Bodenbiologie tropischer Regenwälder. Geogrephische Rundschau 49(1): 24-31. [ Links ]
Bird , S. D.; Coulson , R. N.; Fisher , R. F. 2004. Changes in soil and litter arthropod abundance following tree harvesting and site preparation in a loblolly pine (Pinus taeda L.) plantation. Forest Ecology and Management 202: 195-208. [ Links ]
Bultman , T. L.; Uetz , G. W. 1982. Abundance and community structure of forest floor spider following litter manipulation. Oecologia 55: 34-41. [ Links ]
Colwell , R. K. 2005. EstimateS: statistical estimation of species richness and shared species from samples. Version 7.5.1. University of Connecticut, USA. Available at: <http://viceroy.eeb.uconn.edu/EstimateS>. [ Links ]
Clarke , K. R.; Warwick , R. M. 1994. Change in Marine Communities. UK, National Research Concil. 144 p. [ Links ]
Ferreira , R. L.; Mar ques , M. M. G. S. M. 1998. A fauna de artrópodes de serapilheira de áreas de monocultura com Eucalyptus sp. e mata secundária heterogênea. Anais da Sociedade Entomológica do Brasil 27(3): 359-403. [ Links ]
Foeli x, R. F. 1996. Biology of spiders. New York, Oxford University Press. 336p. [ Links ]
Hammer , O.; Harper , D. A. T. 2009. Past: Paleontological Statistics. Version 1.97. Available at: <http://folk.uio.no/ohammer/past>. [ Links ]
Höfer , H.; Brescovit , A. D. 2001. Species and guild structure of a Neotropical spider assemblage (Araneae) (Reserva Florestal Adolpho Ducke, Manaus, Amazonas, Brazil). Andrias 15: 99-120. [ Links ]
Höfer , H.; Martius , C.; Beck , L. 1996. Decomposition in an Amazonian rainforest after experimental litter addition in small plots. Pedobiologia 40: 70-576. [ Links ]
IBGE, 2004. Mapa de Biomas do Brasil, primeira aproximação. Rio de Janeiro: IBGE. Available at: <http://www.ibge.gov.br>. [ Links ]
Lima , W. P. 1993. Impacto ambiental do eucalipto. 2ed. São Paulo, Editora da Universidade de São Paulo. 302 p. [ Links ]
Lo- Man- Hung , N. F.; Gardner , T. A.; Ribeiro- Júnior , M. A.; Barlow , J.; Bonaldo , A. B. 2008. The value of primary, secondary, and plantation forests for Neotropical epigeic arachnids. Journal of Arachnology 36: 394-401. [ Links ]
Majer , J. D. ; Recher , H. F. 1999. Are eucalypts Brazil´s friend or foe? An entomological viewpoint. Anais da Sociedade Entomológica do Brasil 28(2): 185-200. [ Links ]
May , R. M. 1986. The search for patterns in the balance of nature: Advances and retreats. Ecology 67: 1115-1126. [ Links ]
Moço , M. K. S.; Gama- Rodrigues , E. F.; Gama- Rodri gues , A. C.; Correia , M. E. F. 2005. Caracterização da fauna edáfica em diferentes coberturas vegetais na região norte fluminense. Revista Brasileira de Ciência do Solo 29: 555-564. [ Links ]
Nyffeler , M.; Sterling , W. L.; Dean , D. 1994. A Insetivorous activities of spiders in United States field crops. Journal of Applied Entomology 118: 113-128. [ Links ]
Pellens , R.; Garay , I. 2000. Edaphic macroarthropod communities in fast-growing plantations of Eucalyptus grandis Hill ex Maid (Myrtaceae) and Acacia mangium Wild (Leguminosae) in Brazil. European Journal of Soil Biology 35: 77-89. [ Links ]
Pillar , V. P.; Müller , S. C.; Castilhos , Z. M. S.; Jac ques , A. V. A. 2009. Campos Sulinos - conservação e uso sustentável da biodiversidade. Brasília, MMA. 403 p. [ Links ]
Plagens , M. J. 1983. Population of Misumenops (Araneida: Thomisidae) in two Arizona cotton fields. Environmental Entomology 12: 572-575. [ Links ]
Primack , R.; Rodrigues , E. 2002. Biologia da conservação. Londrina, Editora Planta. viii+328p. [ Links ]
Rinaldi , I. M. P. 2005. Spiders of a young plantation of eucalypt: diversity and potential predator of the most frequent arboreal species. Acta Biológica Paranaense 34: 1-13. [ Links ]
Rodrigues , E. N. L.; Mendonça , Jr., M. S.; Ott , R. 2009. Spider diversity in a rice agroecosystem and adjacent areas in southern Brazil. Revista Colombiana de Entomología 35(1): 78- 86. [ Links ]
Rypstra , A. L.; Carter , P. E; Balfourd , R. A.; Mar shall , S. D. 1999. Architectural features of agricultural habitats and their impact on the spiders inhabitants. Journal of Arachnology 27: 371-377. [ Links ]
Souza , A. L. T. 2007. Influência da estrutura do habitat na abundância e diversidade de aranhas. In: Gonzaga , M. O.; Santos , A. J.; Japyass ú, H. F. eds. Ecologia e comportamento de aranhas. Rio de Janeiro, Interciência Editora, p.25-44. [ Links ]
Toti , D. S.; Coyle , F. A.; Miller , J. A. 2000. A structured inventory of Appalachian Grass Bald and Heath Bald spider assemblages and a test of species richness estimator performance. Journal of Arachnology 28: 329-345. [ Links ]
Uetz , G. W. 1979. The influence of variation in litter habitats on spider communities. Oecologia 40: 29-42. [ Links ]
Uetz , G. W.; Halaj , J.; Cady , A. B. 1999. Guild structure of spiders in major crops. Journal of Arachnology 27: 270-280. [ Links ]
Whitmore , C.; Slotow , R.; Crouch , T. E.; Dippernaar- Schoeman , A. S. 2002. Diversity of spiders (Araneae) in a savanna reserve, Northern Province, South Africa. Journal of Arachnology 30: 344-356. [ Links ]
Wilco x, B. A.; Murphy , D. D. 1985. Conservation strategy: the effects of the fragmentation on extinction. The American Naturalist 125: 879-887. [ Links ]
Wise , D. H. 1993. Spiders in ecological webs. Cambridge, Cambridge University Press. xiii+328p. [ Links ]
Young , O. P.; Edwards , G. B. 1990. Spiders in United States field crops and their potential effect on crop pests. Journal of Arachnology 18(1): 1-27. [ Links ]

