










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488On-line version ISSN 2665-4385
Rev. Colomb. Entomol. vol.36 no.2 Bogotá July/Dec. 2010
Ensamblaje de escarabajos coprófagos (Scarabaeidae: Scarabaeinae) del Bosque Tropical Seco, Bahía Concha, Santa Marta (Colombia)
Assemblage of Coprophagous Beetles (Scarabaeidae: Scarabaeinae) of Tropical Dry Forest in Bahia Concha, Santa Marta (Colombia)
Jeison Barraza M.1, Jenilee Montes F.1, Neis Martínez H.2 y Cuauhtémoc Deloya3
1 Est. Biología, Semillero Investigación Insectos (NEOPTERA) del Caribe colombiano. Grupo Biodiversidad del Caribe colombiano. Programa de Biología, Universidad del Atlántico. Ciudadela Universitaria, km 7, Vía Puerto Colombia. Barranquilla, Colombia. jeisonbarraza@gmail.com
2 Docente investigador. Grupo Biodiversidad Del Caribe. Programa de Biología, Universidad del Atlántico, Ciudadela Universitaria, km 7, Vía Puerto Colombia. Barranquilla - Colombia. neisjosemartinez@yahoo.es,neismartinez@email.uniatlantico.edu.co
3 Ph.D., Instituto de Ecología, A.C. (Red Interacciones Multitróficas). Carretera antigua a Coatepec 351, El Haya, 91070 Xalapa, Veracruz, México. cuauhtemoc.deloya@inecol.edu.mx. Autor para correspondencia.
Recibido: 23-mar-2010 • Aceptado: 10-nov-2010
Resumen: En un paisaje de bosque tropical seco, en la ensenada de Bahía Concha, Santa Marta, Colombia, se evaluó la variación espacio-temporal del ensamblaje de coleópteros coprófagos asociados a tres zonas con diferente grado de intervención antrópica. Se realizaron muestreos mensuales desde marzo hasta octubre del 2008, utilizando trampas de caída, cebadas con excremento humano. Se calculó la riqueza, abundancia, diversidad y la variación espacio-temporal de la estructura de la comunidad. Se capturaron 24.571 individuos pertenecientes a 22 especies. La especie dominante fue Canthidium sp. con 7.860 individuos. El bosque tropical seco antropizado y el pastizal presentaron la mayor riqueza (20) y abundancia (9.644 y 11.330 respectivamente); mientras que en la zona de bosque conservado se reportan 3.567 individuos agrupados en 17 especies. Temporalmente, mayo presentó un mayor número de capturas (5.139) y marzo la menor (151). El ANOSIM (p < 0,05), demostró que existe un patrón temporal en la comunidad debido a la precipitación, lo que está asociado a los cambios que se producen en el suelo y en la mayor disponibilidad y composición del recurso alimenticio. Los resultados demuestran la sensibilidad del gremio ante distintos niveles de intervención antrópica y la relación directa de la precipitación con el ensamblaje de coleópteros coprófagos en el área de estudio.
Palabras clave: Canthidium . Intervención antrópica. Patrón de nidificación.
Abstract: In a tropical dry forest landscape, in the inlet of Bahia Concha, Santa Marta, Colombia, we evaluated the spatio-temporal variation in the assemblage of coprophagous coleopterans associated with three zones with different degrees of anthropogenic disturbance. Samples were collected monthly from March to October 2008 using pitfall traps baited with human excrement. We calculated the richness, abundance and spatio-temporal variation of the structure of the community. We captured a total of 24,571 individuals belonging to 22 species. The dominant species was Canthidium sp, with 7,860 individuals. The disturbed tropical dry forest and the grassland had the highest richness (20) and abundance (9,644 and 11,330); while in the conserved forest area 3,567 individuals grouped in 17 species were reported. Temporally, May presented the greatest number of captures (5,139) and March the least (151). The ANOSIM (p < 0.05%) demonstrated that there is a temporal pattern in the community owing to precipitation, which is associated with changes that are produced in the soil and in the greater availability and composition of the food resource. The results demonstrate the sensitivity of the guild to different levels of anthropogenic disturbance and the direct relationship of precipitation with the assemblage of coprophagous coleopterans in the study area.
Key words: Canthidium . Antropogenic disturbance. Nesting pattern.
Introducción
El bosque tropical seco (BTs) se considera entre los ecosistemas más degradados, fragmentados y menos conocido en Colombia y actualmente solo representa el 3% de la cobertura original; debido en gran parte a la expansión de las fronteras agrícolas y al sobrepastoreo, disminuyendo la extensión del hábitat original (IAvH 1998).
Este panorama se observó en la ensenada de Bahía Concha, Santa Marta, Colombia, donde se presenta una reducción del hábitat original, debido al aumento de las zonas de pastoreo, agricultura y entresacado de madera, lo que ha generado una alteración del paisaje y un posible cambio en el ensamblaje de grupos faunísticos. Por lo cual, se hace urgente determinar el estado de estos bosques considerando el gremio de los escarabajos coprófagos Scarabaeidae, Scarabaeinae. Este grupo es considerado en la región Neotropical como el principal reciclador del excremento de mamíferos omnívoros y herbívoros, cuyos nutrientes son reincorporados a la cadena alimenticia o al ciclo de nutrientes (Howden y Young 1981; Escobar y Chacón 2000). Además, participan en la aireación del suelo, la dispersión secundaria de semillas y como agentes de control biológico de parásitos presentes en las heces (Halffter 1959; Hanski y Camberfort 1991), debido a que al enterrar el estiércol impiden que los dípteros ovipositen y puedan multiplicarse, contribuyendo de esta forma al control de la diseminación de enfermedades infectocontagiosas (Tapia 2005). Por otro lado, estos insectos son importantes en estudios de biodiversidad por ser sensibles a perturbaciones que alteren tanto la estructura como el microclima del hábitat (Halffter y Mattews 1966; Nealis 1977; Doube 1991).
Los estudios sobre Scarabaeinae en el BTs de la costa Caribe colombiana son escasos. Se tiene el inventario del bosque seco al norte del Tolima (Escobar 1997) y el listado de especies de Santa Marta (Jiménez et al. 2008), y la diversidad de los Scarabaeinae en la Cuenca del Río Gaira (Martínez et al. 2009). Sin embargo, la composición y estructura de los escarabajos coprófagos en las estribaciones de la Sierra Nevada de Santa Marta no ha sido documentada. Por lo anteriormente expuesto, el objetivo del presente estudio fue evaluar la variación espacio-temporal del ensamblaje de escarabajos coprófagos (Scarabaeinae) en un paisaje de bosque tropical seco en la ensenada de Bahía Concha, Santa Marta, Colombia.
Materiales y Métodos
El estudio se realizó en la finca Villa Concha (11º18’120’’N, 74º08’824’’O), donde se ubican fragmentos típicos del bosque tropical seco, en la formación costera de la SNSM, departamento del Magdalena, norte de Colombia (Fig. 1). En el BTs predominan en su composición las familias Bignoniaceae, Bombacaceae, Cactaceae, Capparidaceae, Fabaceae y Rubiaceae (Holdridge et al. 1971). Las especies características son Prosopis juliflora (Sw.) DC; Ficus sp., Spondias monbin L. (1753); Crataeva tapia L., Capparis sp. y Crescentia cujete L.
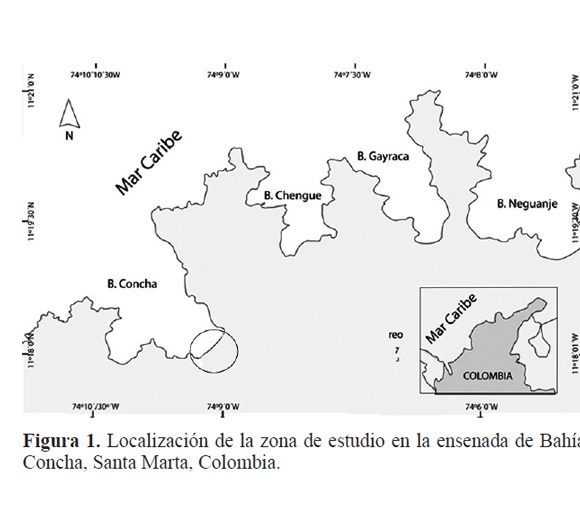
Los muestreos se realizaron de marzo a octubre del 2008, considerando la época seca, primeras lluvias y lluvias. Durante el estudio no se completó el ciclo anual de muestreo por el difícil acceso a la zona y la inseguridad en el área de estudio debido a la extracción ilegal de madera.
Se seleccionaron tres sitios con base en las características de la vegetación y los niveles de intervención antrópica, como el pastoreo y la producción de carbón: a) Bosque tropical seco conservado (BTsC: 45 msnm), vegetación original con dosel cerrado, abundante hojarasca, suelo arenoso con buen drenaje; b) Bosque tropical seco antropizado (BTsA: 33 msnm), actividad humanada con extracción de madera y dosel semiabierto, poca hojarasca, suelo arenoso con drenaje moderado; c) Pastizal, 16 msnm, totalmente antropizado para fines pecuarios, presencia de arbolado disperso, suelo arenoso con drenaje moderado.
En cada sitio se montó un transecto lineal donde se instalaron 12 trampas de caída (“pitfall traps”) separadas 30m (Escobar 2000b), para un total de 288 muestras. Las trampas de caída fueron modificadas, acondicionando al vaso de plástico (500 ml de volumen) en la parte superior un alambre en forma de “L” invertida, y colocando en el extremo el cebo de excremento humano envuelto en gasa. Estas trampas fueron enterradas a nivel del suelo, adicionando una solución de agua con detergente a un cuarto de su capacidad. Las trampas estuvieron expuestas 24 horas y los especímenes capturados fueron conservados en etanol al 70%. Se determinaron a género siguiendo los criterios propuestos por Medina y Lopera (2000) y Vaz de Melo y Edmonds (2007). Adicionalmente, se consultó la colección del Instituto de Investigación de Recursos Biológicos Alexander von Humboldt (IAvH) y del Instituto de Ciencias Naturales (ICN) de la Universidad Nacional de Colombia. Los ejemplares se encuentran depositados en la Colección Entomológica del IAvH y el Laboratorio de Zoología, Universidad del Atlántico (UARC).
Los datos de precipitación fueron suministrados por el Instituto de Hidrología, Meteorología y Estudios Ambientales de Colombia (IDEAM) y con base en los datos se consideró época seca (marzo-abril), primeras lluvias (mayo-septiembre) y lluvias en octubre.
Se registró el número de especies por sitio (S = diversidad alfa) y el número total de individuos (N). Para evaluar el esfuerzo de muestreo y la efectividad del mismo se realizaron curvas de acumulación de especies, utilizando el programa EstimateS 8.2 (Colwell 2009).
La diversidad de especies, se calculó mediante el índice de riqueza de Margalef (d); la dominancia como el número de especies efectivo por sitio, ponderadas de acuerdo con su abundancia relativa con la serie de Números de Hill (N1) y (N2), y para establecer como está repartido el número de individuos se estimó el índice de equidad de Pielou (J’) por medio del paquete estadístico PRIMER 6.0. Para determinar el recambio de especies entre zonas, se utilizó el índice de complementariedad propuesto por Colwell y Coddington (1994). Las descripciones de estos índices se encuentran en Moreno (2001). Para determinar diferencias en la estructura de las especies de escarabajos coprófagos entre los sitios y entre las épocas de muestreo, se aplicó ANOSIM de una vía. Para identificar las especies que caracterizaron o tipificaron los sitios y las épocas a través de su abundancia, se utilizó la rutina SIMPER. Se consideraron las especies que aportaron más del 10% de la abundancia total.
Las especies fueron separadas con base en los patrones de nidificación del alimento en cavadores, rodadores y residentes (Hanski y Camberfort 1991). Adicionalmente se graficó la variación espacial y temporal de estos patrones con el fin de observar su distribución.
Resultados y Discusión
Se capturaron 24.571 individuos, 10 géneros y 22 especies (Tabla 1) que corresponden al 68,75% de las especies registradas por Escobar (1998) para la región Caribe colombiana. A pesar de la alta transformación del paisaje en la zona, se recolectó el 84,61% de las especies reportadas para la región de Santa Marta, Colombia (Jiménez et al. 2009); lo cual es indicativo que en Bahía Concha, todavía se alberga una fauna representativa de las especies nativas de escarabajos coprófagos. Con base en los resultados obtenidos, se hace necesario conciliar con la comunidad aledaña y autoridades del sistema de parques naturales, la inclusión de estos fragmentos en una figura apropiada de protección.
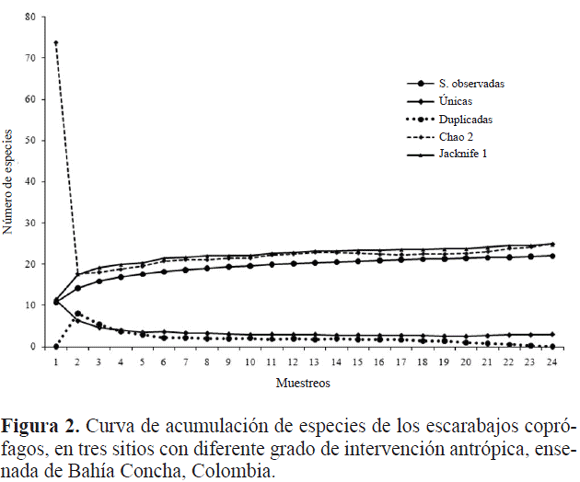
Los estimadores de riqueza esperada (Chao 2 y Jacknife 1) indican que con el esfuerzo de muestreo realizado se obtuvo el 95% de las especies, faltando por capturar una especie (Fig. 2). Lo anterior indica que la técnica de captura, el número de trampas y muestreo, fueron suficientes y eficaces para obtener un número representativo del total de la fauna de escarabajos coprófagos en el área de estudio. Además, se notauna tendencia de la curva de especies únicas y duplicadas a lograr una asíntota, por lo cual se deduce una estabilización de éstas para el muestreo y se concluye que el porcentaje fue óptimo, según lo propuesto por Moreno y Halffter (2000).
Las especies más abundantes son Canthidium sp. (32%), Dichotomius belus (Harold, 1880) (17%), Canthon aberrans Harold, 1868 (15%) y C. mutabilis Lucas, 1857 (14%) (Tabla 1). La abundancia de Canthidium sp. confirma el carácter euritópico del género, además de explotar las heces de diferentes mamíferos, también muestran un comportamiento saprófago secundario, alimentándose de frutos u otras partes de plantas superiores en fermentación (Halffter y Matthews 1966); no fue posible su determinación específica porque la taxonomía del género Canthidium no está resuelta y su ecología es poco conocida en el país, además muy probablemente se trate de un complejo de especies; aunado a su amplia distribución Neotropical (139 especies) y diversidad en Colombia con 27 especies, de las cuales seis están registradas para la Sierra de Santa Marta (Escobar 2000a; Medina et al. 2001; Noriega et al. 2007b).
En el BTsC se registró la menor riqueza (S = 17) y abundancia (14,63%), mientras que el BTsA y Pastizal obtuvieron la mayor riqueza (S = 20) y abundancia (39,24% y 46,11% respectivamente) (Tabla 1). La mayor abundancia en los dos últimos sitios, se debe a la alta disponibilidad del recurso alimenticio, factor determinante en la composición y estructura de la escarabeidofauna coprófaga. Además, se evidenció el aporte de algunas especies (D. belus, C. mutabilis , Canthon septemmaculatum Latreille 1811, Onthophagus clypeatus Blanchard, 1843) que se caracterizan por colonizar hábitats abiertos y alterados donde hay presencia de animales domesticados. Onthophagus acuminatus (Harold, 1880) y Dichotomius sp. se capturaron únicamente en el pastizal, Scybalocanthon sp. fue exclusiva del BTsA. Se catalogan como especies raras por la baja representatividad. Es posible que presenten hábitos especializados, por lo cual no pueden ser detectadas con facilidad con la técnica utilizada. Onthophagus acuminatus Harold, 1880 y Dichotomius sp. capturadas en el pastizal y Scybalocanthon sp. en el BTsA, podrían catalogarse como especies raras por su baja representatividad, tal vez por tener hábitos especializados y no fueron atraídas con la técnica utilizada, o ser consideradas especies turistas que fueron atraídas a las copro-trampas.
En la región, la temperatura media anual es de 27°C con precipitaciones mayores a 233,3 mm, al inicio de la época de lluvias, donde se observa un incremento del 100% en la riqueza específica (Fig. 3). Esto se debe a que en la temporada seca, la humedad del suelo es muy baja, lo cual puede condicionar la emergencia del adulto de la bola-nido, por no estar lo suficientemente ablandada (Bustos-Gómez y Lopera 2003). Además la severidad de la estación seca limita la actividad de estos escarabajos al impedir la construcción de galerías debido a la dureza y compactación del terreno (Janzen 1983). En la época de primeras lluvias y lluvias los valores de riqueza no fluctúan de manera brusca, lo que indica que las especies utilizan diferentes estrategias para evitar la competencia por un mismo recurso, teniendo picos reproductivos alternados (Fig. 4). Resultados similares fueron obtenidos por Vidaurre et al. (2009) en el Jardín Botánico de Santa Cruz de la Sierra (Bolivia) y Oliveira (2009) en la Caatinga, Minas Gerais (Brasil).
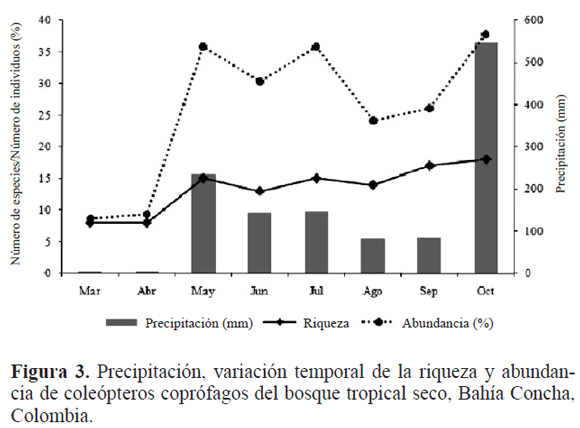
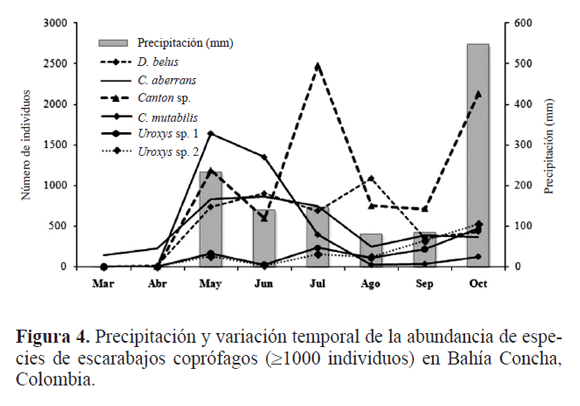
La mayor abundancia de escarabajos coprófagos (21%) se presentó en mayo, que corresponde al inicio del periodo de lluvias (Fig. 3). En esta época puede existir una mayor oferta de recursos tanto a nivel cualitativo como cuantitativo por parte del bosque, lo que representa disponibilidad de recursos como frutos y otros invertebrados para muchos mamíferos. Lo anterior, puede determinar variaciones en la composición y disponibilidad del excremento a lo largo del año (Estrada et al. 1993); generando una mejor y mayor disponibilidad de alimento para los escarabajos coprófagos (Huston 1994). Resultados similares se han obtenido en la región Neotropical, donde las comunidades residentes aumentan sus poblaciones con el inicio y posterior establecimiento del periodo de lluvias. Esto último conlleva a que aumente la cobertura vegetal en los parches de BTs en el área de estudio, contribuyendo a una mayor heterogeneidad de recursos que son aprovechados por estos insectos durante esta época del año (Lauer 1992; Pinheiro et al. 2002; Bustos-Gómez y Lopera 2003; Martínez et al. 2009). Durante agosto y septiembre disminuye la precipitación y la abundancia, aumentando ambas en octubre con el inicio de las lluvias fuertes en la región. Andresen (2005) propone que el aumento y descenso de la abundancia y los cambios en la estructura de la comunidad de escarabajos coprófagos en bosques secos, están ligados a la precipitación, tal como fue obtenido en el presente estudio.
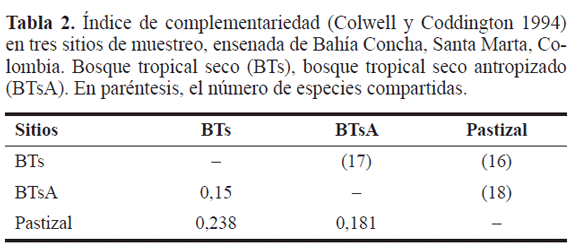
Los índices de diversidad, equitatividad y complementariedad muestran que los valores varían muy poco entre los sitios, debido a que comparten entre el 73% y 82% de las especies. En el BTsC se presentó la mayor diversidad y equitatividad, indicando que las especies se distribuyen de forma más homogénea, al ser un hábitat más complejo y maduro, como ha sido referido por Noss (1983). La mayor complementariedad se observó entre el BTsC y Pastizal, indicando que la tasa de recambio de especies es mayor, a medida que aumenta la degradación del bosque (Tabla 2). Además, tampoco se observaron diferencias significativas en la estructura de la comunidad entre los sitios (R = 0,071, P = 0,09). Esto debido a que los escarabajos coprófagos tienen una alta capacidad para la consecución del recurso alimenticio, y algunas especies penetran al bosque intervenido o potreros donde existen mamíferos domesticados que aporten excremento, como ha sido informado por Escobar y Chacón de Ulloa (2000). Sin embargo, Phanaeus hermes Harold, 1863 y P. prasinus Harold, 1868 fueron capturadas en los fragmentos poco antropizados, lo cual indicaría que son especies umbrófilas, aunque no necesariamente especies indicadoras de la calidad del hábitat, como ha sido referido por Escobar 2000b).
En cambio, si se observaron diferencias en la estructura de la comunidad (R = 0,073, P = 0,01) entre épocas de muestreo, mostrándose un patrón temporal en el área de estudio y una alternancia en los picos de emergencia (> 1000 individuos; Fig. 4). Esto permite la coexistencia de distintas especies, disminuyendo la competencia interespecifica (Hanski y Camberfort 1991). Canthidium sp. y D. belus son especies que presentan comportamiento reproductivo multivoltino con varios picos de reproducción, y posiblemente realicen una mayor explotación y aprovechamiento del recurso alimenticio y de los diferentes requerimientos nutricionales que necesitan (Janzen 1983; Edwards 1991). Lo anterior está relacionado con la composición y disponibilidad del excremento proporcionado por los mamíferos durante la época de lluvias, debido a que muchos de estos se alimentan del follaje, flores, frutos frescos e invertebrados (Estrada et al. 1998).
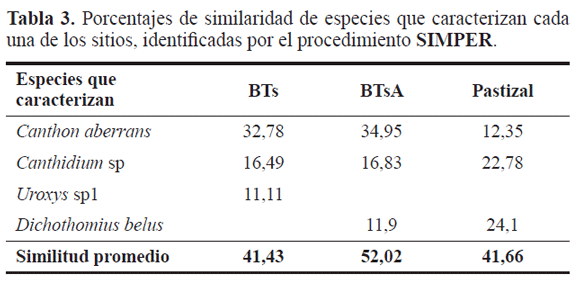
Cada sitio fue caracterizado por la abundancia de dos especies, C. aberrans y Canthidium sp, por el aporte a la abundancia en cada lugar y pueden considerarse especies euritópicas, con amplios rangos de tolerancia. Bustos-Gómez y Lopera (2003) las han citado tanto en hábitats perturbados como no perturbados. Uroxys sp. 1 caracteriza el BTsC, con una tendencia a localizarse en ambientes con poca intervención antrópica. Dichotomius belus caracteriza los sitios antropizados (BTsA y pastizal) Este resultado difiere a lo sugerido por Bohórquez y Montoya (2009), quienes asocian esta especie con áreas menos perturbadas. El porcentaje más bajo de similitud se presentó entre BTs (41,43%) y el más alto en BTsA con 52,02%. Sin embargo, estos valores de similaridad son muy bajos, debido a que el número de especies es muy bajo y los aportes a la abundancia de C. aberrans y Canthidium sp. 2 son muy similares (Tabla 3).
Canthon aberrans puede considerarse generalista y multivoltina, debido a que estuvo presente durante las tres épocas, confirmándose que la estacionalidad no influye de igual forma sobre el comportamiento de las especies presentes de escarabajos coprófagos, y que probablemente esté más relacionado con la eficiencia y aprovechamiento de diversos recursos alimenticios (Halffter y Favila 1993). Similar caso se observó en D. belus, pero en menor proporción en la época seca, tal vez debido a que es una especie umbrófila con requerimientos específicos de humedad. Resultados similares obtuvieron Bohórquez y Montoya (2009), donde observaron un aumento en su abundancia durante el periodo de lluvias en la Reserva Forestal de Colosó, Sucre.
Canthidium sp. mostró una alta relación con los niveles de precipitación, tipificando la época de primeras lluvias y la época de lluvias, donde sus principales picos de emergencia se presentaron en los meses donde ésta fue mayor. Lo anterior puede estar asociado a los cambios que se producen en la humedad del suelo y en la disponibilidad del recurso alimenticio. Canthon mutabilis caracterizó la época de primeras lluvias, teniendo la mayor abundancia en mayo y descendiendo rápidamente en los siguientes meses. Por otra parte, las especies de Uroxys caracterizaron la época de lluvias, lo cual indica una sincronización y estacionalidad con esta época del año (Tabla 3, Fig. 4).
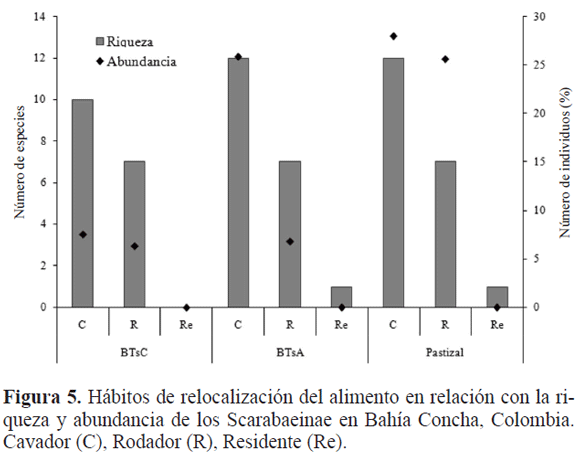
El gremio de los cavadores predominó en los tres sitios (64,67%), seguido por los rodadores (35,19%) y por último los residentes (0,02%). Este último gremio está representado por Eurysternus plebejus Harold, 1880, especie coprófaga con siete especímenes. Su escasa representatividad en las muestras se debe al método de captura, como ha sido informado por Halffter y Edmonds (1982). El predominio del gremio de los cavadores (Fig. 5) se debe a la textura arenosa de los sitios, lo que favorece la habilidad competitiva, aumentando la rapidez con la que estas especies entierran el alimento y la construcción de las bolas-nido (Hanski y Camberfort 1991). Estos datos son similares a los obtenidos por Noriega et al. (2007) en un bosque con tres grados de alteración, en los Llanos Orientales (Meta, Colombia); donde se observó una proporción de 1,1: 1 entre cavadores-rodadores y una baja proporción (1,19%) de residentes.
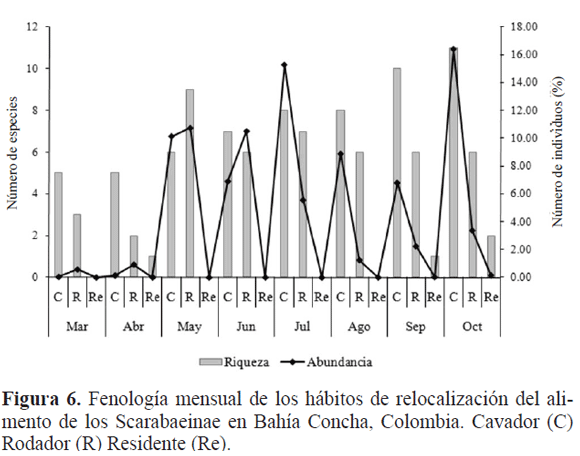
L a variación temporal de las especies muestra una dominancia en riqueza de cavadores sobre rodadores y residentes durante el periodo de muestreo. En cambio, la abundancia mostró un patrón bimodal con un mayor número de rodadores durante la época seca y primeras lluvias y una mayor proporción de especies cavadoras durante el periodo de primeras lluvias y lluvias altas en la zona (Fig. 6). Esto debido a que las condiciones físicas del terreno cambian de una estructura rígida y compacta a una blanda, lo cual beneficia a este gremio trófico. Resultados similares son reportados por Martínez et al. (2009) en la vertiente Noroccidental de la Sierra Nevada de Santa Marta, Colombia.
Agradecimientos
A José Dávila propietario de la Finca “Villa Concha”, por permitir la realización del proyecto en su propiedad. A la comunidad de esta finca por su amabilidad y colaboración. Héctor García gentilmente realizó la identificación de los escarabajos. A Claudia Medina (Instituto Alexander von Humboldt) y Germán Amat (ICN, Universidad Nacional de Colombia) por su colaboración en la identificación y su gran aporte bibliográfico.
Literatura citada
ANDRESEN, E. 2005. Effects of season and vegetation type on community organization of dung beetles in a tropical dry forest. Biotropica 37 (2): 291-300. [ Links ]
BOHÓRQUEZ, J.; MONTOYA, J. 2009. Abundancia y preferencia trófica de Dichotomius belus (Coleoptera: Scarabaeidae) en la reserva forestal de Colosó, Sucre. Boletín del Museo de Entomología de la Universidad del Valle 10 (1): 1-7. [ Links ]
BUSTOS-GOMEZ, F.; LOPERA, A. 2003. Preferencia por cebo de los escarabajos coprófagos (Coleoptera: Scarabaeidae: Scarabaeinae) en un remanente de bosque seco tropical al norte del Tolima (Colombia). M3m monografías tercer milenio SEA, Zaragoza. 3 (30): 59-65. [ Links ]
COLWELL, R. K. 2009. ESTIMATES: Statistical Estimation of Species Richness and Shared Species from Samples, Version 8.2. Available at http: http://viceroy.eeb.uconn.edu/estimates [ Links ]
COLWELL, R.; CODDINGTON, J. 1994. Estimating terrestrial biodiversity through extrapolation. Philosophical Transactions of the Royal Society London 345: 101-118. [ Links ]
DOUBE, B.M. 1991. Dung beetles of southern Africa, p. 133-155. En: Hanski, I. & Y. Cambefort (Eds.). Dung beetle ecology. Princeton University, New Jersey [ Links ]
EDWARDS, P. B. 1991. Seasonal variation in the dung of African grazing mammals and consequences for coprophagous insects. Functional Ecology 5: 617-628. [ Links ]
ESCOBAR, F. 1997. Estudio de la comunidad de coleópteros coprófagos (Scarabaeidae: Scarabaeinae) en un remanente de Bosque Seco al norte del Tolima. Caldasia 19 (3): 419-430. [ Links ]
ESCOBAR, F. 1998. Análisis regional de la comunidad de escarabajos coprófagos (Coleóptera: Scarabaeidae: Scarabaeinae) de los Bosques Secos de la región Caribe de Colombia, p. 72-75. En M. E. Chávez & N. Arango (Eds.). Informe Nacional del Estado de la Biodiversidad, Colombia, Tomo I: Diversidad Biológica. Instituto Humboldt, PNUMA y Ministerio del Medio Ambiente. Bogotá. [ Links ]
ESCOBAR, F.; CHACÓN DE ULLOA, P. 2000. Distribución espacial y temporal en un gradiente de sucesión de la fauna de coleópteros coprófagos (Scarabaeidae, Aphodiinae) en un bosque Tropical Montano, Nariño, Colombia. Revista de Biología Tropical 48 (4): 961-975. [ Links ]
ESCOBAR, F. 2000a. Diversidad y Distribución de los Escarabajos del Estiércol (Coleoptera: Scarabaeidae: Scarabaeinae) de Colombia. p. 197-210, En F. Martín-Piera, J. J. Morrone & A. Melic (Eds.). Hacia un proyecto CYTED para el inventario y estimación de la diversidad entomológica en Iberoamérica: PrIBES- 2000, Monografías Tercer Milenio, Zaragoza, España. [ Links ]
ESCOBAR, F. 2000b. Diversidad de coleópteros coprófagos (Scarabaeidae: Scarabaeinae) en un mosaico de hábitat en la Reserva Natural Nukak, Guaviare, Colombia. Acta Zoológica Mexicana (ns) 79: 103-121. [ Links ]
ESTRADA, A.; HALFFTER, G.; COATES-ESTRADA, R.; MERITT, D. A. 1993. Dung beetles attracted to mammalian herbivore (Alouatta palliata) and omnivore (Nasua narica) dung in the tropical rain forest of Los Tuxtlas, Mexico. Journal of Tropical Ecology 9: 45-54. [ Links ]
ESTRADA, A.; R. COSTA-ESTRADA, A. A. DADDA.; P. CAMMARANO. 1998. Dung and carrion beetles in tropical rain forest fragments and agricultural habitats at Los Tuxtlas, Mexico. Journal of Tropical Ecology 14: 577-593. [ Links ]
HALFFTER, G. 1959. Etología y Paleontología de Scarabaeinae (Coleóptera: Scarabaeidae). Ciencia 29: 164-178. [ Links ]
HALFFTER, G.; MATTHEWS, E. G. 1966. Natural history of dung beetles of the subfamily Scarabaeinae (Coleoptera: Scarabaeidae). Folia Entomológica Mexicana 12-14: 1-312. [ Links ]
HALFFTER, G.; EDMONDS, W.D. 1982. The nesting behavior of dung beetle (Scarabaeinae) Instituto de Ecología, México. 220 p. [ Links ]
HALFFTER, G.; FAVILA, M. E. 1993. The Scarabaeinae (Insecta: Coleoptera) an animal group for analysing, inventorying and monitoring biodiversity in tropical rainforest and modified landscapes. Biology International 27: 15- 21. [ Links ]
HANSKI, I.; CAMBEFORT, Y. 1991. Dung beetle ecology. Princeton University Press, New Jersey. 520p. [ Links ]
HOLDRIDGE, L. R.; GRENKE, W.C.; HATHEWAY, W.H.; LIANG. T.; TOSI. J.A. 1971. Forest environments in tropical life zones, a pilot study. Pergamon Press, Oxford. P. 747 En: INSTITUTO ALEXANDER VON HUMBOLDT, IAVH. 1998. El bosque seco tropical (Bs-T) en Colombia. Grupo de Exploraciones y Monitoreo Ambiental GEMA, IAVH, Villa de Leyva. P. 1-24. [ Links ]
HOWDEN, H.; YOUNG, O. 1981. Panamanian Scarabaeinae: taxonomy, distribution and habits (Coleoptera: Scarabaeidae). Contributions of the American Entomological Institute 18: 1-204. [ Links ]
HUSTON, M. 1994. Biological Diversity. The coexistence of species on changing landscapes. Cambridge. University Press N. Y. 671 p. [ Links ]
INSTITUTO ALEXANDER VON HUMBOLDT, IAvH. 1998. Programa de inventarios de la biodiversidad Grupo de exploración y monitoreo GEMA. El bosque seco tropical (Bs-T) en Colombia. http://araneus.humboldt.org.co/download/inventarios/bst/ doc3.pdf. [ Links ]
JANZEN, D.H. 1983. Seasonal changes in abundance of large nocturnal dung beetles (Scarabaeidae) in a Costa Rica deciduous forest and adjacent horse pastures. Oikos 33: 274-283. [ Links ]
JIMÉNEZ, L.; MEDIETA, W.; GARCIA, H.; AMAT-GARCÍA, G. 2008. Notas sobre los escarabajos coprófagos (Coleoptera: Scarabaeinae) en ambientes secos de la región de Santa Marta, Colombia. Acta Biológica Colombiana, 13 (2): 203-208. [ Links ]
LAUER, W. 1992. Climate and weather. p. 7-53. En Lieth, H; Werger, M.J.A. Tropical rain forest ecosystems. Biogeographical and ecological studies. Elsevier, New York. [ Links ]
MARTÍNEZ, N. J.; GARCÍA, H.; PULIDO, A.; OSPINO, D.; NARVÁEZ, J.C. 2009. Fauna de escarabajos coprófagos (Coleoptera: Scarabaeinae) en un gradiente altitudinal en la vertiente Noroccidental de la Sierra Nevada de Santa Marta, Colombia. Neotropical Entomology 38 (6): 708- 715. [ Links ]
MEDINA, C.; LOPERA, A. 2000. Clave Ilustrada para la Identificación de Géneros de Escarabajos Coprófagos (Coleoptera: Scarabaeinae) de Colombia. Caldasia 22 (2): 299-315. [ Links ]
MEDINA, C.; LOPERA, A.; VITOLO, A.; GILL, B. 2001. Escarabajos Coprófagos (Coleoptera: Scarabaeidae: Scarabaeinae) de Colombia. Biota Colombiana 2(2): 131-144. [ Links ]
MORENO, C. E. 2001. Métodos para medir la biodiversidad. M&T- Manuales y Tesis SEA, vol.1. Zaragoza, 84 p. [ Links ]
MORENO, C. E.; HALFFTER, G. 2000. Assessing the completeness of bat biodiversity inventories using species accumulation curves. Journal of Applied Ecology 37: 149-158. [ Links ]
NEALIS, V. G. 1977. Habitat associations and community analysis of south Texas dung beetles (Coleoptera: Scarabaeinae). Canadian Journal of Zoology 5: 138-147. [ Links ]
NORIEGA, J.; REALPE, E.; FAGUA, G. 2007. Diversidad de escarabajos coprófagos (Coleoptera: Scarabaeidae) en un bosque de galería con tres estadios de alteración. Universitas Scientiarum, edición especial (12): 51-63. [ Links ]
NOSS, R.F. 1983. A regional landscape approach to maintain diversity. Bio-Science 33: 700-706. [ Links ]
OLIVEIRA, V. H. F. 2009. Variaçao na fauna de besouros rola - bosta (Coleoptera: Scarabaeinae) entre habitats de cerrado, mata seca e mata ciliar em uma regiao de transiçao cerrado - caatinga no norte de Minas Gerais. Anais do III Congresso Latino Americano de Ecología, 10 a 13 de Setembro de 2009, Sao Lourenço [ Links ]
M.G. PINHEIRO, F.; DINIZ, I. R.; COELHO, D.; BANDEIRA, M. P. S. 2002. Seasonal pattern of insect abundance in the Brazilian Cerrado. Australian Ecology 27:132-136. [ Links ]
TAPIA, A. 2005. Preferencia por fecas de tapir amazónico (Tapirus terrestris) de escarabajos estercoleros (Coleoptera: Scarabaeidae: Scarabaeinae) en bosque secundario amazónico. Tapir Conservation 14 (17): 24- 28. http://www.tapirs.org/Downloads/ newsletters/TapirCons17web.pdf [ Links ]
VAZ-DE-MELLO, F. Z.; EDMONDS, W.D. 2007. Géneros y subgéneros de la subfamilia Scarabaeinae (Coleoptera: Scarabaeidae) de las Américas. Versión 2.0 Español. 30 p. http://www.scarabnet.org/ScarabNet/Taxonomy/Entries/2007/1/26_Neotropical_ Genera_Key_V1.4_files/claveespaniol2.0.pdf [ Links ]
VIDAURRE, T.; LEDEZMA, J.; AMAYA, M.; FUENTES, K. 2009 Variación temporal de los escarabajos coprófagos (Coleoptera: Scarabaeidae: Scarabaeinae) del Jardín Botánico Municipal de Santa Cruz de La Sierra, Bolivia. Anais do III Congresso Latino Americano de Ecología, 10 a 13 de Setembro de 2009, Sao Lourenço-MG. [ Links ]

