










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488On-line version ISSN 2665-4385
Rev. Colomb. Entomol. vol.36 no.2 Bogotá July/Dec. 2010
Estadios y variación temporal de Eurygerris fuscinervis (Heteroptera: Gerridae) en los Andes de Colombia
Stages and the temporal variation of Eurygerris fuscinervis (Heteroptera: Gerridae) in the Andes of Colombia
DORA N. PADILLA-GIL1 y OSVALDO ARCOS P.2
1 Ph. D., Profesora Asociada, Universidad de Nariño, Ciudad Universitaria Torobajo, Facultad de Ciencias Exactas y Naturales. Departamento de Biología. Bloque 3, piso 4, Nariño, Colombia. dnpadilla@udenar.edu.co Autora para correspondencia.
2 Zootecnista. Laboratorista, Universidad de Nariño, Ciudad Universitaria Torobajo, Nariño.
Recibido: 3-mar-2010 • Aceptado: 26-oct-2010
Resumen: Se describen e ilustran los estadios del desarrollo post-embrionario de Eurygerris fuscinervis. El número de individuos por estadio y la variación temporal, en época seca y lluviosa, se estudiaron en seis localidades, ubicadas en cuatro municipios del Departamento de Nariño, Colombia: Pasto, Buesaco, Funes y Tangua. Se colectaron 1.342 individuos, 831 adultos y 511 ninfas, de cinco estadios. Las poblaciones presentaron estabilidad temporal. No hubo diferencias significativas en cuanto a las épocas seca y lluviosa, como tampoco respecto a la altitud (2.260−2.700 m).
Palabras clave: Insectos acuáticos. Chinches patinadores. América del Sur.
Abstract: The stages of post-embryonic development of Eurygerris fuscinervis are described and illustrated. The number of individuals per stage and the temporal variation, during the dry and rainy seasons, were studied in six localities located in four municipalities of the Department of Nariño, Colombia: Pasto, Buesaco, Funes y Tangua. A total of 1,342 individuals were collected, 831 adults and 511 nymphs, corresponding to five stages. The populations exhibited temporal stability; there were no significant differences with respect to the dry and rainy seasons, nor with respect to altitude (2,260−2,700 m).
Key words: Aquatic insects. Water-striders. South America.
Introducción
Los gérridos están ampliamente distribuidos en los ecosistemas acuáticos lóticos y lénticos; en general están asociados a la película superficial del agua. En Colombia se han publicado varios estudios taxonómicos sobre los gérridos, entre estos los de Aristizabal (2002), Padilla y Nieser (2003), Polhemus y Polhemus (1995) y trabajos como los de Morales-Castaño y Molano-Rendón (2009) en relación con el género Eurygerris; sin embargo, los estudios sobre la fenología y ecología del grupo son escasos tanto en Colombia como en el neotrópico. Molano-Rendón et al. (2008) presentaron una descripción sobre el hábitat de las diferentes especies de gérridos encontrados en Colombia.
La especie objeto de estudio es Eurygerris fuscinervis (Berg, 1898) en Colombia se presenta en la Región Andina y el Valle de Sibundoy. Morales-Castaño y Molano-Rendón (2009) incluyen en su rango de distribución a Chile, Argentina, Bolivia, Perú, Ecuador, Venezuela, Guatemala y México. En los ecosistemas acuáticos altoandinos se observa la emergencia continua de la especie E. fuscinervis, de ciclo multivoltino, con cinco estadios ninfales, y adultos, tanto ápteros como macrópteros (observación personal). Por ser una especie ampliamente distribuida en los ecosistemas acuáticos de los municipios de Pasto, Buesaco, Funes y Tangua, se eligió para evaluar su variabilidad en cuanto a abundancia en los diferentes estadios, así como de adultos ápteros y alados, en las épocas seca y lluviosa y la posible influencia de la altitud, en tal variabilidad (2260-2700 m).
El objetivo de este trabajo es determinar la variación temporal y altitudinal de E. fuscinervis en ecosistemas acuáticos de los Andes del suroeste de Colombia. Estos resultados constituyen un aporte al conocimiento de la biología y ecología de esta especie en el neotrópico y pueden servir de base para futuros estudios sobre fenología, ecología y genética de poblaciones y comunidades.
Materiales y Métodos
Área de estudio. Se muestrearon seis localidades, ubicadas en cuatro municipios: Pasto, Buesaco, Funes y Tangua (Fig.1). Todas las localidades presentan un régimen de precipitación bimodal, la época más seca comprende los meses de junio a septiembre, la más lluviosa son octubre y noviembre (IGAC 1985). A continuación se citan las localidades de muestreo, y las fechas en las cuales se realizaron los muestreos, durante el 2009.
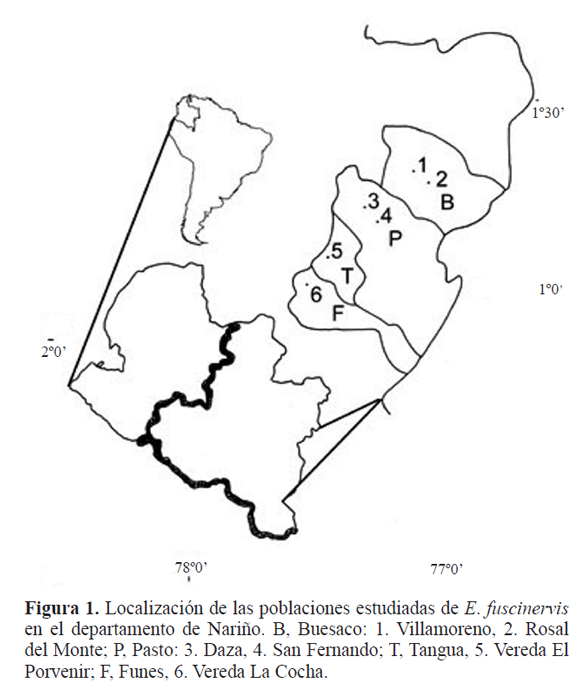
En el Municipio de Pasto, 1º10’12”N -77º12’36”W, las localidades: Daza y San Fernando, con una altitud de 2700 m y 2640 m, respectivamente. Daza: Junio 30, Octubre 12; San Fernando: Junio 17, Noviembre 11. En el Municipio de Buesaco, 1º23’38”N -77º08’54”W, las localidades: Villamoreno y Rosal el Monte, con una altitud de 2330 m y 2420 m, respectivamente. Junio 19, Octubre 12.
En el Municipio de Funes, 0º48’-1º03’N-77º53-33’W, Vereda La Cocha a 2340 m de altitud. Junio 3, Noviembre 2. En el Municipio de Tangua, 3º19’N-77º24’W, Vereda El Porvenir a 2260 m de altitud. Junio 24, Noviembre16.
Muestreo. Cada localidad se visitó dos veces, una en época seca y otra en época lluviosa. Los muestreos se realizaron en las horas de la mañana. Se colectó el material con una red. Cuatro colectores trabajaron simultáneamente en cada sitio de muestreo. Los especímenes fueron transportados al laboratorio en frascos con etanol al 70%, donde posteriormente se procesaron y determinaron cada uno de los estadios. Los adultos fueron sexados y separados en ápteros y macrópteros (con ala completa); los estadios fueron asignados con base en las medidas del largo total y el ancho del pronoto.
Todos los especímenes examinados para el estudio de su desarrollo postembrionario proceden de la localidad de Funes, único sitio donde se observaron y colectaron huevos. En la descripción de los estadios ninfales se utilizan las siguientes abreviaturas: largo y ancho: L/A; largo total: Lt; ancho de la cabeza, a través de los ojos: Ac; ancho del pronoto, en su parte más ancha: Ap; ancho máximo del mesonoto: Am; abdomen largo total y ancho máximo a través del conexivo: abdomen L/A; ninfa hembra: Nh; ninfa macho: Nm. Todas las medidas en milímetros.
Para la comparación entre poblaciones se realizó un conteo total de los especímenes por estadio ninfal sin tener en cuenta el sexo de los estadios IV y V. Todos los adultos ápteros y macrópteros se contaron. Las abreviaturas adoptadas para cada estadio son: Ma, macho áptero; Ha, hembra áptera; Mm, macho macróptero; Hm, hembra macróptera; estados ninfales n1-n5. Todos los especímenes fueron depositados en la colección de entomología de la Universidad de Nariño (PSO-CZ).
Se evaluó el efecto de altitud y la estacionalidad frente al número de individuos según su estadio de desarrollo, adultos ápteros macho y hembra, adultos macrópteros macho y hembra, y cinco estadios ninfales. Se aplicó el análisis de varianza ANOVA/MANOVA, mediante el software Statgraphics (2000).
Resultados
Desarrollo post-embrionario. Se caracteriza el desarrollo post-embrionario de la forma áptera de E. fuscinervis y se ilustra con base en la hembra, (Fig. 2). Para todos los estadios se presentan las medidas morfométricas luego la descripción.
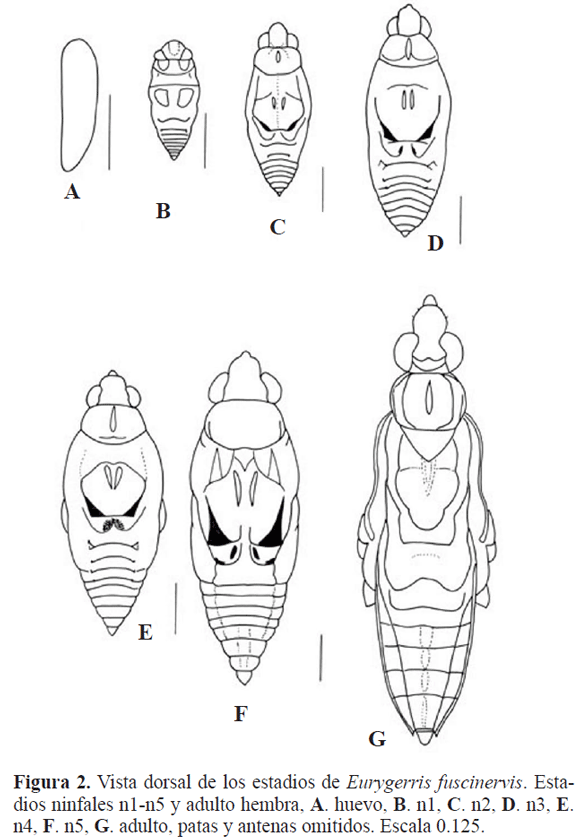
Los tres primeros estadios son indistintos para machos y hembras, en los estadios IV y V se observa dimorfismo sexual, el cual se describe. El mesonoto en todos los estadios ninfales es más largo que el pronoto conservando una proporción 2:1; el mesonoto es más largo que el metanoto manteniendo una proporción 4:1.
Huevo (Fig. 2A) L 1,92; A 0.56. Color blanco, textura lisa, embebido por una sustancia transparente que lo protege, antes de eclosionar toma un color amarillo. Los huevos son depositados en la superficie y a ambos lados de una ramita flotante, estas ramas flotantes están cerca de la orilla del lago.
Estadio I (Fig. 2B) Lt 2,72; Ac 0,76; Ap 0,96; Am 1,28. Color castaño claro; centro dorsal de la cabeza, lados del pronoto, meso y metanotos con áreas castaño oscuro; ojos negros. Es visible la sutura ecdisial de la cabeza y una sutura que divide el mesonoto, éstas ausentes en el resto de estadios ninfales.
Estadio II (Fig. 2C) Lt 4,93; Ac 1,12; Ap 1,24; Am 1,60. Color castaño claro; ojos negros; disco del pronoto con una mancha clara a lo largo de la línea media dorsal, mesonoto con dos manchas cortas paralelas al eje medio del cuerpo, lados del margen mesopostnotal ligeramente esclerotizados y castaño oscuro.
Estadio III (Fig. 2D) Lt 5,86; Ac 1,28; Ap 1,44; Am 2,0. Similar a la descripción del estadio II, difiere en el mayor grado de esclerotización y oscurecimiento de los lados del márgen mesopostnotal y metanoto con una mancha castaño oscuro a ambos lados de la línea media.
Estadio IV (Fig. 2E) Nh: Lt 6,53; Ac 1,32; Ap 1,52; Am 2,16; abdomen L/A 1,92/1,84; esterno abdominal VII L/A 0,32/0,84 Nm: Lt 6,26; Ac 1,28; Ap 1,36; Am 1,84; abdomen L/A 1,76/1,28; esterno abdominal VII L/A 0,32/0,52
En los estadios IV y V se acentúa la coloración castaño oscuro mesopostnotal, en el metanoto, y el delineamiento castaño oscuro de los segmentos del abdomen dorsal.
Dimorfismo sexual: la Nh es un poco más robusta que la Nm y el abdomen de la Nm es más angosto. El conexivo de la Nm es ligeramente levantado en los segmentos I-IV y en la Nh es horizontal; el esterno abdominal VII tanto de la Nm como de la Nh es igual de largo pero en la Nh es más ancho; el esterno abdominal VIII en la Nh presenta una línea media difusa y la Nm carece de ésta.
Estadio V (Fig. 2F) Nh: Lt 8,10; Ac 1,60; Ap 1,72; Am 2,32; abdomen L/A 2,6/1,88; esterno abdominal VII L/A 0,56/1,04. Nm: Lt 7,60; Ac 1,40; Ap 1,68; Am 2,04; abdomen L/A 2,48/1,52; esterno abdominal VII L/A 0,36/0,60. Las ninfas en este estadio mantienen sus diferencias respecto al conexivo más engrosado en la Nm; el esterno abdominal VII, en la Nm es casi la mitad del tamaño de la Nh; en el esterno abdominal VIII de la Nh se observa una línea media, la Nm carece de ésta; urito IX es más largo que ancho en la Nm y en la Nh es más ancho que largo.
Adultos ápteros. Ma: Lt 9,46; Ac 1,32; Ap 1,60; Am 2,0; abdomen L, segmentos I-VII/A 2,44/1,40; esterno abdominal VII L/A 0,40/0,84. Ha (Fig. 2G): Lt 11,6; Ac 1,84; Ap 1,80; Am 2,84; abdomen L, segmentos I-VII/A 3,28/2,20; esterno abdominal VII L/A 0,8/1,44
Caracterización del hábitat. Daza, Pasto: se caracteriza por tener pozos pequeños de aguas lentas, rodeados por escasa vegetación circundante, sin vegetación flotante, sustrato compuesto por arena y rocas pequeñas, agua cristalina y profundidad menor a 50 cm. E. fuscinervis se encontró como única especie.
San Fernando, Pasto: presenta pozos pequeños de aguas estancadas, rodeados por escasa vegetación de cultivos o potreros, sin vegetación flotante, sustrato arcillo-limoso, agua ligeramente turbia y profundidad menor a 50 cm. E. fuscinervis se encontró como única especie.
Villamoreno, Buesaco: muestra un manantial de agua, de baja corriente, rodeado por árboles, una parte expuesta y la otra umbrosa, sin vegetación flotante, sustrato limoso, agua cristalina y profundidad menor a 50 cm. E. fuscinervis coexiste con Rhagovelia villamoreno n. sp.
Rosal el Monte, Buesaco: presenta pozos de aguas lentas, rodeados por escasa vegetación circundante, sin vegetación flotante, sustrato compuesto por rocas pequeñas, agua cristalina y profundidad menor a 70 cm. E. fuscinervis se encontró como única especie.
Vereda La Cocha, Funes: es un gran lago, rodeado por gramíneas, con vegetación flotante cerca de la orilla, agua cristalina y profundidad mayor a un metro. E. fuscinervis coexiste con Buenoa funensis Padilla-Gil, 2010.
Vereda El Porvenir, Tangua: es un lago, rodeado por gramíneas, cubierto por helecho acuático flotante Azolla filiculoides y profundidad mayor a un metro. E. fuscinervis se encontró como única especie.
En varias localidades de Buesaco, no consideradas para este estudio, de Rosal del Monte y Vereda Pajajoy, E. fuscinervis se encontró en el Río Buesaquito y quebradas de aguas rápidas, expuestas y cristalinas, coexistiendo con Rhagovelia buesaquensis Padilla-Gil, 2009.
En las localidades de Daza, Villamoreno, Rosal del Monte y Tangua también se observaron especímenes de E. fuscinervis en pozos pequeños formados por aguas lluvia cercanos a los caminos. Generalmente los adultos se encuentran en grupos separados de aquellos donde están las ninfas; los adultos macrópteros también se encuentran separados de los ápteros.
Análisis comparativo de las poblaciones. Se colectaron y caracterizaron 1342 individuos, 831 adultos y 511 ninfas de los diferentes estadios ninfales de las seis localidades. El número total de individuos por época en cada una de las localidades se indica en la Tabla 1. La localidad donde se colectó el mayor número de individuos fue Rosal del Monte (334) y aquella con el menor número de individuos fue Daza (106). No hubo diferencias en la abundancia de los estadios de E. fuscinervis entre épocas seca y lluviosa en ninguna de las localidades (F = 1,15, P > 0.05).
Los estadios por localidad presentaron diferencias significativas (P < 0,05), para cuatro localidades: San Fernando (F = 39.75), Villamoreno (F = 7,04), Rosal del Monte (F = 10,29), y Funes (F = 10.85). Las dos localidades que no presentaron diferencias significativas fueron Daza y Tangua ( P > 0,05). La altitud no marcó diferencias significativas (F = 1,91, P > 0,05).
En San Fernando, los estadios con diferencias significativas fueron los adultos ápteros machos y hembras (Fig. 3A); en Villamoreno las diferencias las presentaron los estadios adultos ápteros machos, las hembras ápteras y el estadio ninfal 1 (Fig. 3B); en Rosal del Monte se presentaron cuatro grupos, los adultos ápteros machos, las hembras ápteras, los adultos macrópteros machos y hembras, y el resto de estadios ninfales (Fig. 3C). En Funes, se presentan tres grupos, machos ápteros, hembras ápteras y el resto de los estadios (Fig. 3D).
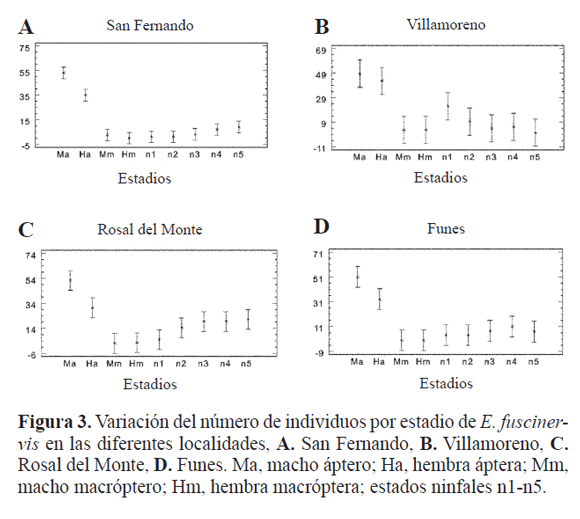
Es de señalar que en Daza y Funes no se registraron adultos macrópteros y en San Fernando y Rosal del Monte, se colectaron sólo en época lluviosa; en las otras dos localidades predominaron en época lluviosa.
En la época seca, en Tangua los estadios ninfales uno y dos y en Funes, el estadio ninfal uno, no se registraron. Predominan los machos ápteros con más del 55% (55-79%) para casi todas las localidades excepto para Villamoreno, en época lluviosa, donde dominaron las hembras ápteras con un 66% y Tangua, también en época lluviosa, con el 52% de hembras ápteras. En época seca, se colectaron cuatro individuos macrópteros en Villamoreno (dos) y en Tangua (dos). En época lluviosa los machos de la forma macróptera fueron levemente más frecuentes con el 51% que las hembras 49%.
Discusión
En Argentina Mazzucconi y Bachmann (1993a, 1993b) registraron formas micrópteras para E. fuscinervis; además señalan la asociación de las formas áptera y macróptera con ambientes estables e inestables respectivamente. En el material colectado en las seis localidades, no se detectó la forma micróptera; predominó la forma áptera (95%), y los machos ápteros; del 5% de los adultos correspondientes a la forma macróptera, el 90% fue colectado en época lluviosa, y la relación entre machos y hembras se aproximó a uno.
La presencia de mayor número de individuos alados en la época lluviosa, probablemente está relacionada con la dispersión de la especie, a través de la colonización de nuevas charcas temporales, formadas en época de mayor precipitación. Por otra parte como señala Andersen (1982) el polimorfismo alar puede ser favorecido, si los hábitats preferidos son una mezcla de sitios permanentes y temporales, tal parece ser el caso de E. fuscinervis. Los mecanismos ambientales o genéticos que determinan el polimorfismo alar en esta especie constituyen un campo inexplorado.
El rango altitudinal en el cual se colectaron los especímenes (2.260-2.700 m) no marco una diferencia significativa en las poblaciones. Se amplió el rango altitudinal, indicado para esta especie por Molano-Rendón et al. (2008) desde los 1800 m hasta los 2.700 m.
No hubo diferencias significativas entre las épocas seca y lluviosa; también se observó el predominio de adultos ápteros y la presencia de todos los estadios ninfales. Esto lejos de ser resultados concluyentes debido al muestreo, nos permiten verificar la existencia de estas poblaciones y conocer algunos aspectos sobre su estado actual espacio-temporal e igualmente sirven como punto de partida para el planteamiento de otros interrogantes sobre la biología y ecología de esta especie como son: cuáles son las características y duración del ciclo de vida?, cuántos huevos son ovipositados por hembra y en qué lapso?, se mantienen las frecuencias de emergencia de los adultos formas ápteras y macrópteras por población/año?, qué factores ambientales influyen para el mantenimiento de las poblaciones a través del año?, las poblaciones presentan suficiente cantidad de adultos y estadios ninfales para asegurar su continuidad temporal y espacial?, qué factores inciden para mantener la abundancia de los distintos estadios?
Agradecimientos
Agradecemos a los estudiantes G. Montenegro y N. Nicola, por su ayuda en las salidas de campo. A la Vicerrectoría de Investigaciones (VIPRI) de la Universidad de Nariño por financiar esta investigación. A Carlos E. Sarmiento, editor de la revista y a dos revisores anónimos por la revisión del manuscrito.
Literatura citada
ANDERSEN, N. M. 1982. The semiaquatic bugs (Hemiptera, Gerromorpha). Entomonograph 3: 1-455. [ Links ]
ARISTIZÁBAL, H. 2002. Los hemípteros de la película superficial del agua en Colombia. Parte 1. Familia Gerridae. Academia Colombiana de Ciencias Exactas Físicas y Naturales. Editora Guadalupe, Bogotá. Colombia. 239 p. [ Links ]
IGAC (NSTITUTO GEOGRÁFICO AGUSTÍN CODAZZI). 1985. Nariño Aspectos Geográficos. Bogotá D. C. 118 p. [ Links ]
MAZZUCCONI, S. A.; BACHMANN, A. O. 1993a. Micropterismo en Eurygerris fuscinervis (Berg, 1898) (Heteroptera: Gerridae). Revista de la Sociedad Entomológica Argententina 52(1-4): 53- 56. [ Links ]
MAZZUCCONI, S. A.; BACHMANN, A. O. 1993b. Familia Gerridae (Heteroptera): estudio comparativo de las alas de las especies argentinas, chilenas y uruguayas. Revista de la Sociedad Entomológica Argentina 52(1-4): 87-99. [ Links ]
MOLANO-RENDÓN F.; MORALES-CASTAÑO I. T.; SERRATO- HURTADO C. 2008. Clasificación y Hábitats de Gerridae (Heteroptera-Gerromorpha) en Colombia. Acta Biológica Colombiana 13(2):41-60. [ Links ]
MORALES-CASTAÑO, I.; MOLANO-RENDÓN, F. 2009. Revisión de los géneros Eurygerris y Tachigerris (Hemiptera: Tachygerrini) para la región neotropical. Revista Mexicana de Biodiversidad 80: 395-410. [ Links ]
PADILLA, D. N.; NIESER, N. 2003. Nueva especie de Tachygerris y nuevos registros de colecta de las Gerridae (Hemiptera: Heteroptera) de Colombia. Actualidades Biológicas 25(78): 39-49. [ Links ]
POLHEMUS, J. T.; POLHEMUS, D. A. 1995. A phylogenetic review of the Potamobates fauna of Colombia (Heteroptera: Geridae) with descriptions of three new species. Proceedings of Entomological Society of Washington 97 (2): 350-372. [ Links ]
STATGRAPHICS. 2000. Statgraphics version 5.0. www.statgraphics.com [ Links ]

