










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488On-line version ISSN 2665-4385
Rev. Colomb. Entomol. vol.37 no.2 Bogotá July/Dec. 2011
Sección Básica
Wasmannia auropunctata (Hymenoptera: Formicidae): ¿unicolonial o multicolonial? en el valle geográfico del río Cauca
Wasmannia auropunctata (Hymenoptera: Formicidae): ¿Unicolonial o multicolonial? in the valley Cauca river
BEATRIZ SALGUERO R.1, INGE ARMBRECHT2, HERNANDO HURTADO T.3 y ÁNGELA MARÍA ARCILA C.4 1 m.Sc. Departamento de matemáticas Universidad Autónoma de occidente, Cali, Colombia. bsalguero@uao.edu.co autora para correspondencia. 2 Ph.D. Departamento Biología Universidad del Valle. Cali - Colombia. inge.armbrecht@correounivalle.edu.co. 3 m.Sc. Departamento Estadística Universidad del Quindío. Armenia - Quindío lhhurtado@uniquindio.edu.co. 4 Ph.D. Corpoica-Caribia, zona Bananera-magdalena. aarcila@corpoica.org.co recibido: 3-mar-2011 - Aceptado: 30-oct-2011
Resumen: Con el objetivo de identificar la estructura social de la pequeña hormiga de fuego Wasmannia auropunctata, en ambientes drásticamente perturbados, se modeló la distribución espacial de esta especie en dos parches de bosque seco con grados de perturbación contrastantes, ubicados al sur y norte del valle geográfico del río Cauca, regiones para las cuales W. auropunctata es considerada nativa con estructura social multicolonial. Para los análisis se tuvieron en cuenta las variables densidad poblacional y nidos/m2 en los biotopos: borde, interior de bosque y potrero. la modelación estuvo enmarcada en el campo de la geoestadística, donde se estimaron los modelos de semivariogramas y parámetros asociados, que definieron la continuidad espacial de las variables de interés y que fueron utilizados para la interpolación y construcción de mapas en regiones del espacio no muestreadas, a través del krigeado. Se encontró que la distribución espacial estuvo bien descrita por modelos de semivariogramas esféricos y exponenciales, que explicaron el patrón de autocorrelación espacial. los mapas de interpolación de obreras por cebo y de nidos/m2, permitieron mostrar como la pequeña hormiga de fuego presenta una estructura social unicolonial en el parche de bosque de San Julián, al sur del departamento del Valle asociado a un mayor grado de perturbación, lo que puede llegar a justificar su baja agresividad intraespecífica en su rango nativo. En contraste, en el parche de bosque de Alejandría al norte del departamento y con un menor grado de perturbación, presentó una estructura social multicolonial con alta agresión intraespecífica.
Palabras clave: Estructura social. Fragmentación. Colombia. modelación matemática. Hormiga del fuego.
Abstract: With the aim of identifying little fre ant Wasmannia auropunctatas social structure in drastically disturbed environments, we modeled the spatial distribution of this species in two patches of dry forest with contrasting levels of disturbance, at southand and north of the Cauca river valley, a region for which W. auropunctata is considered native with a multicolonial social structure. For the analysis, we took into account both, population density and nests/m2, in biotopes: edge, interior forest and pasture. The modeling was framed in the feld of geostatistics, were, by the estimation of semivariograms models and parameters, the spatial continuity of variables we were interested in was defined. They were used to interpolate and construct the maps for those regions not sampled all of this through kriging. it was found that the spatial distribution model was well described by spherical and exponential semivariograms, which explained the pattern of spatial autocorrelation. interpolation maps of workers per bait and nests/m2 allowed usto show that the little fre ant has an unicolonial social structure in the patch of san Julians forest, at south of the valley. That was associated with a greater degree of disturbances, which may justify their low intraspecific aggression in its native range. in contrast, in the Alejandrias forest patch situated at north of the department and which exhibited a lower disturbance, it introduced a social structure multicolonial and high intraspecific aggression.
Key words: Social structure. Fragmentation. Colombia. mathematical modeling. Fire ant
Introducción En Colombia, el valle geográfico del río Cauca, alguna vez estuvo dominado por una espesa vegetación arbórea propia de bosque seco tropical (bs-T), pero debido a la actividad antrópica sufrió una drástica fragmentación (Velasco 1982), quedando tan solo con 2% de cobertura boscosa (Arcila 2007). Este escenario fue parte de un fenómeno más amplio, y hoy se considera al bs-T como uno de los ecosistemas más amenazados en el Neotrópico (Didier et al. 2001). El reemplazo de casi la totalidad del bosque por cultivos, convierte el paisaje en parches de bosque rodeados por una matriz altamente transformada, hostil para la conservación de especies que requieren grandes extensiones de bosque continuo para su desarrollo y que facilita la invasión de especies generalistas (Kattán 2002). Aunque se conoce poco de los efectos de la fragmentación del hábitat sobre las comunidades de invertebrados (Didham et al. 1998), se ha documentado que algunas especies son insensibles a los cambios estructurales y utilizan hábilmente los hábitats del borde (Kotze y Samways 2001), mientras que otras, incluyendo varias especies de insectos, responden rápidamente a factores relacionados con los bordes (laurance y Vasconcelos 2001). En la evaluación de ambientes perturbados, los insectos son de gran utilidad pues cumplen con una serie de criterios ecológicos, biogeográficos, de conservación, monitoreo y calidad del ambiente (Brown 1991; Kremen et al. 1993; iAvH 1998). las hormigas (Hymenoptera: Formicidae) cumplen con las condiciones requeridas (Brown 1991) y pueden refejar la naturaleza de la vegetación, las condiciones físicas, perturbación y posiblemente la variedad de otros invertebrados que están presentes en un área dada (Burbidge et al. 1992). En Colombia se han realizado estudios sobre la fauna de hormigas en pequeños parches de bosque seco a lo largo del valle geográfico del río Cauca y se han encontrado más de 200 especies de hormigas (Armbrecht y Chacón de Ulloa 1999; Chacón de Ulloa y Armbrecht 2006). Dentro de este gran grupo se destaca la pequeña hormiga de fuego Wasmannia auropunctata (roger, 1863), por su abundancia, facilidad para identificar y colectar. Se distribuye en un rango geográfico bastante amplio, es nativa de los trópicos americanos y ha invadido zonas tropicales y subtropicales de otros continentes del mundo (Silberglied 1972; Wetterer et al. 1999; Ulloa - Chacón y Cherix 1990). Esta hormiga es una especie generalista en su dieta, anida en hojarasca y posee características propias de las especies vagabundas (tramp), descritas por Holldobler y Wilson (1990), como poliginas, unicoloniales, reproduciéndose por fisión de las colonias y dispersándose por las actividades humanas (Ulloa-Chacón y Cherix 1990; Passera 1994). En parches de bosque seco tropical del Valle del río Cauca se ha encontrado una correlación negativa entre la incidencia de captura de Wasmannia auropunctata y la riqueza de otras hormigas, por esta razón ha sido propuesta por Armbrecht y Ulloa - Chacón (2003), como un indicador de baja biodiversidad de la comunidad de hormigas. Todo parece indicar que la fragmentación causa pérdidas de especies por aislamiento liberando recursos y nichos que son aprovechados por W. auropunctata favoreciendo así la colonización de estos hábitats perturbados (Arcila 2007; Philpott et al. 2010). En su rango introducido, la pequeña hormiga de fuego no exhibe agresividad hacia sus congéneres (Clark et al. 1982; Ulloa - Chacón 1990). En su rango nativo, se ha reportado estructura social multicolonial con agresividad intraespecífica. Sin embargo, lópez (2005) encontró estructura unicolonial con baja agresión intraespecífica en zonas agrícolas altamente perturbadas, donde se comporta como especie invasora, mas no dentro de los bosques relativamente bien conservados en el valle geográfico del río Cauca. A nivel interespecífico, la pequeña hormiga de fuego sí presenta un comportamiento agresivo (Wetterer y Porter 2003) especialmente donde ha sido introducida afectando negativamente otras especies silvestres (Silberglied 1972; Clark et al. 1982). Los diferentes comportamientos de agresividad de W. auropunctata se expresan claramente en la distribución espacial que exhibe la hormiga, tanto en hábitats perturbados como naturales. Cuando su distribución es muy agregada, su tendencia es a comportarse como en su rango nativo y cuando su distribución tiende a uniforme se comporta como invasora, exhibiendo unicolonialidad, es decir, múltiples nidos interco-nectados y poca agresividad intraespecífica (Ulloa-Chacón y Cherix 1990; lópez 2005). Dada la anterior problemática con la pequeña hormiga de fuego, es necesario explorar el comportamiento de las poblaciones en su rango nativo utilizando herramientas de la estadística espacial. Usualmente, se utilizan métodos de la estadística clásica para los análisis de distribución espacial, basados en índices de dispersión, que no tienen en cuenta la localización exacta de la muestra. Estos métodos suponen independencia en los datos e igual distribución, por lo que pueden errar en la diferenciación de patrones espaciales disímiles (ramírez-Dávila y Porcayo-Camargo 2008; robertson 1987). Al no tener en consideración toda la información espacial, los investigadores podrían llegar involuntariamente a resultados y predicciones erróneas (Carrol y Pearson 2000; rossi et al. 1992; moral 2004), debido a que se puede presentar autocorrelación. El uso de la geoestadística ha ido en incremento, para fortalecer estudios donde se presenta autocorrelación espacial, puesto que comprende un conjunto de herramientas y procedimientos estadísticos que permiten analizar, modelar y predecir los valores de una variable que se muestra distribuida en el espacio o en el tiempo en forma continua y autocorrelacionada (isaaks y Srivastava 1989; Díaz - Francés 1993). A diferencia de los índices de dispersión, que ignoran la localización de las muestras (Taylor 1984), los métodos geoestadísticos requieren información georreferenciada obteniéndose la localización espacial bidimensional exacta de la variable de estudio, adicionalmente el tratamiento de la información con estas técnicas permite elaborar mapas de interpolación muy útiles, para establecer centros de concentración de las poblaciones. En las dos últimas décadas se han utilizado en ecología técnicas geostadísticas con notable éxito (rossi et al. 1992), para obtener mapas de interpolación de variables ecológicas en regiones del espacio no muestreadas (isaaks y Srivastava 1989; rossi et al. 1992; Díaz - Francés 1993) donde al conocer la distribución espacial de un insecto plaga en alguna región geográfica, se pueden aplicar controles para disminuir sus poblaciones (Díaz-Francés 1993; liebhold et al. 1993; ramírez-Dávila et al. 2005; ramírez-Dávila y Por-cayo-Camargo 2008; Nordmeyer 2009). Dada la necesidad de conocer la distribución espacial de W. auropunctata en los parches de bosque seco tropical existentes en el valle geográfico del río Cauca, especialmente en el bosque de San Julián al sur, un sitio altamente perturbado y en el bosque Alejandría al norte, un sitio poco perturbado, se planteó el presente estudio que propone presentar con herramientas de la geoestadística, la modelación matemática de la distribución de la hormiga W. auropunctata. El estudio modela dos variables, obreras por cebo y nidos/m2, (en adelante nidos y agregados significarán lo mismo) en los biotopos, interior de bosque, borde de bosque y matriz circundante. Se espera que la modelación actúe como un ente informador desde el cual se pueda discutir la estructura social de la pequeña hormiga de fuego en dos parches de bosque altamente contrastantes de acuerdo a su grado de perturbación. los resultados permitirán establecer estrategias de manejo de estos ecosistemas encaminados a la coexistencia de otras especies de hormigas con W. auropunctata.
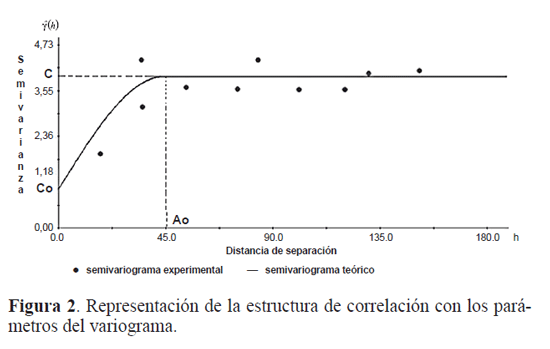
Materiales y Métodos El trabajo se realizó en dos etapas: el muestreo de W. auropunctata, siguiendo los protocolos de muestreo optados por Arcila (2007) y la modelación matemática de la distribución espacial de la pequeña hormiga de fuego. Arcila (2007) seleccionó 10 parches de bosque a lo largo de la cuenca alta del río Cauca, de los cuales dos fueron seleccionados para el presente estudio: San Julián (3°638,8N 76°3141,2W) en el sur limitando con el departamento del Cauca y Alejandría (4°4958,6N 75°532.4W) en el norte limitando con el departamento de risaralda. Para estos sitios se registró una altura sobre el nivel del mar entre 900 y 1100m respectivamente, con temperaturas promedio de 24°C y precipitaciones anuales fuctuando entre 1000 y 2000mm (CVC 1990), características propias del bosque seco tropical (bs-T) (Fig. 1). los parches de bosque presentan una estructura interna muy similar (Armbrecht y Ulloa - Chacón 2003), pero fueron diferentes en tamaño, forma, grado de aislamiento y tipo de matriz que los rodea, la cual por lo general se encontró constituida por cultivos de caña de azúcar y potreros. El área total de los parches fue de 10,6 hectáreas para San Julián y de 15,3 hectáreas para Alejandría, con una extensión interna de bosque sucesional tardío del orden de 3,5 Ha, caracterizándose como bosques muy pequeños (Arcila 2007). En San Julián y Alejandría se obtuvieron datos del número de obreras y nidos/m2, en el borde de bosque, interior de bosque y matriz circundante (potrero o caña de azúcar). Para el análisis se tomaron sólo los registros que permitieran información en términos de un arreglo en el plano así fuese irregular, situación recomendable para los análisis de estructura espacial (giraldo 2002; isaaks y Srivastava 1989; Díaz - Francés 1993; zas 2006). Arcila (2007) y lópez (2005) seleccionaron para la matriz que fue el elemento más extenso del paisaje, un borde consistente en una franja de 50m de ancho a partir del límite entre el parche y la matriz. De acuerdo con Didham et al. (1998), ésta debe ser superior a los 100 metros, pero dada el área tan pequeña de la zona boscosa, este ancho dejaría las zonas de estudio sin interior de bosque. En cada uno de los biotopos se trazaron aleatoriamente transectos lineales, con estaciones de muestreo separadas 20m, procurando que su distribución fuera uniforme en todo el área, se alcanzaron cerca de 30 estaciones para cada biotopo, para un total de 90 para cada localidad. los transectos de la matriz se dispusieron de forma paralela a 50m de los bordes del bosque (lópez 2005). En cada estación de muestreo se colocaron cebos de atún, en cuadros de papel blanco de 4cm x 4cm, expuestos por 3 horas, después de las cuales fueron revisados para registrar la presencia de W. auropunctata, posteriormente se colectaron y se llevaron al laboratorio para realizar el conteo de obreras capturadas. En aquellas estaciones de muestreo donde se registró la presencia de W. auropunctata se colocaron cuadrantes de 1m2 separados por una cuadrícula de 20cm x 20cm. En cada uno de los cuadrantes se realizó la búsqueda directa en hojarasca y tronquitos huecos, para establecer la densidad de nidos/m2 de W. auropunctata. Cada estación de muestreo fue georreferenciada usando un geoposicionador portátil garmín 12 Xl. Análisis Geoestadístico. A través del análisis geoestadístico se determinó la estructura de autocorrelación entre los datos y su uso en la predicción o interpolación a través de las técnicas conocidas como kriging (Hurtado et al. 2007). inicialmente, la información se organizó en bases de datos Excel georreferenciadas en un sistema de coordenadas UTm, con origen en centro - occidente (4°3556,57N 77°451,3W) (Com. Per. 2007, Jorge mejía PhD, oSSo observatorio Sismológico del Sur occidente). Se realizó con el software statgraphics Plus 5.1, el análisis exploratorio de datos originales para obreras por cebo y nidos/m2, para reconocer valores atípicos, analizar las medidas de tendencia central en los datos. Con el software gS+ (versión 5.1) se obtuvieron los mapas de ubicación de los sitios de muestreo con su respectivo registro. Cuando el supuesto de normalidad no se satisfzo, se realizó una transformación Box - Cox tipo potencia, la cual suaviza la varianza Cabe anotar, que esta transformación es reversada para la interpolación e interpretación de los resultados. En cada sitio de estudio se estimó un semivariograma experimental a partir de los registros de obreras por cebo y nidos /m2, a través de la siguiente ecuación (isaaks y Srivastava 1989) Donde y(h) corresponde al semivariograma experimental para el intervalo de distancia h; N(h) corresponde al número de pares de puntos muestrales separados por el intervalo de distancia h; Z(x¡) es el valor de la variable regionalizada de estudio en el punto x. y Z(x¡+h) es el valor de la variable regionalizada de estudio en el punto x¡ +h. En geoestadística, las variables regionalizadas, son aquellas variables aleatorias medidas en el tiempo o en el espacio y que presentan estructura de correlación, son funciones Z(x) que toman un valor en cada punto x del espacio y que definen un proceso estocástico (Hurtado et al. 1996). Se consiguieron semivariogramas experimentales, a los cuales se les ajustó un modelo teórico que corresponde a una función definida positiva (isaaks y Srivastava1989), la gráfica resultante de y(h) en función de h, describe el grado de autocorrelación presente. la mayoría de los modelos teóricos de ajuste, contienen tres parámetros comunes: el efecto pepita o nugget effect, que corresponde a la variación causada por errores experimentales o de muestreo, generalmente ocasionados por una microvariabilidad en la variable de estudio, la meseta o sill que es la cota superior del semivariograma y corresponde a la máxima variabilidad de la variable regionalizada y el rango o alcance, que es aquel valor de h para el cual el semivariograma alcanza la meseta, en términos prácticos es la distancia a partir de la cual dos observaciones son independientes, (Fig. 2). En los anteriores ajustes, es importante determinar el nivel de dependencia espacial. Este valor se obtiene del cociente entre los valores del efecto pepita y la meseta, expresando su resultado como un porcentaje. Si este valor es menor del 25% se considera como un nivel de dependencia alto; un valor entre 26% y 75% corresponde a un nivel de dependencia moderado; finalmente un nivel por encima del 76% corresponde a un nivel de dependencia bajo (Cambardella et al. 1994). También, como establece Díaz - Francés (1993) para que el modelo de correlación describa la realidad es recomendable que el valor del efecto pepita no supere el 50% del valor de la meseta ó cota superior. Posterior al establecimiento de la dependencia espacial, se usa el modelo del semivariograma ajustado, para realizar la interpolación en aquellos puntos que no fueron muestreados, usando algoritmos del kriging, los cuales permiten estimar el valor de la variable de interés en un punto muestral específico a partir de los restantes valores muestrales. El procedimiento anterior permitió elaborar un mapa representando el comportamiento de las dos variables de densidad de W. auropunctata en la zona estudiada. También se realizaron en Arc map versión 9, los mapas de distribución de obreras por cebo con las isolineas de la variable nidos/m2 superpuestas. 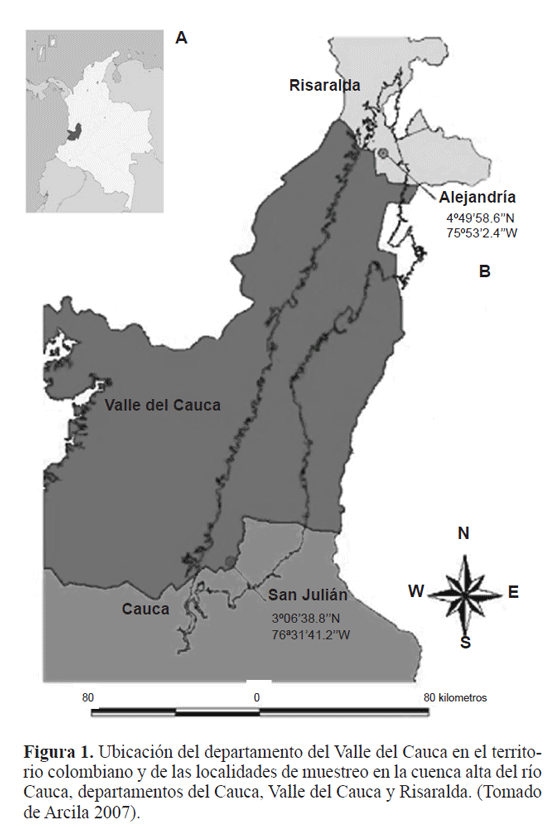
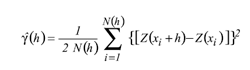
Resultados
De acuerdo con el análisis exploratorio de los datos, San Julián presentó 657 obreras por cebo y 4,2 nidos /m2 en promedio, mientras que en Alejandría los registros fueron de 20 y 1,31 respectivamente. En ambos parches de bosque, el coeficiente de variación para obreras por cebo y para nidos/m2, fue mayor a 100%.
Para todas las variables se estimaron variogramas acotados o con meseta finita, en ningún caso exhibieron semivarianza constante en función de la distancia, (pepita puro) lo que muestra que los modelos encontrados, si refejaron una estructura de dependencia espacial para las variables. las variables en las dos localidades se llevaron a los datos originales por la transformación inversa y muestran que el valor del efecto pepita fue menos del 48,1% del valor de la meseta en la mayoría de los modelos, esto conduce a establecer que al menos el 51,9% de la variabilidad total es debida a la dependencia alta en estas escalas de muestreo, resultando en la mayoría de los modelos con una dependencia espacial moderada (Tabla 1). El valor de la pepita o nugget no superó el 50% de la meseta ó cota superior del variograma, por lo que los modelos conseguidos para este estudio describen correctamente la variabilidad total, con una dependencia espacial moderada.
El coeficiente r2 para obreras por cebo, igual a 0,924 en San Julián y a 0,789 en Alejandría, indicaron la calidad del ajuste del semivariograma teórico al semivariograma experimental. En la figura 3 se presentan los modelos de semivariogramas para las variables descritas en las dos localidades.
los mapas de distribución bi y tridimensional de obreras por cebo para San Julián muestran un gradiente positivo, con una agregación hacia la región suroriente de la localidad (Fig. 4). El mapa geográfico correspondiente a la zona de estudio, permitió evidenciar la tendencia de la densidad obreras por cebo hacia la zona del borde en la esquina suroriente, en límites con la matriz de caña (Fig. 5). los mapas de distribución bi y tridimensional de agregados o nidos por metro cuadrado para Alejandría muestran tres agrupamientos (Fig. 6): uno en potrero, al noroeste, y los otros dos en el borde, hacia el costado este en el norte y en cercanías al sur. Similar a San Julián el mapa geográfico correspondiente a la zona de estudio, permitió evidenciar la tendencia de la densidad obreras por cebo hacia las zonas del borde en cada una de las concentraciones (Fig. 7).
los mapas de distribución de obreras por cebo junto con las isolineas de nidos por metro cuadrado para las dos localidades exhibieron una interpolación muy similar al de obreras por cebo, se observa como densidades altas de obreras por cebo, coinciden con los mayores registros de nidos por metro cuadrado, que en San Julián alcanzó 15 nidos/m2 y en Alejandría llegó hasta 9 nidos/m2 (Fig. 8).
Discusión
mediante herramientas de la geoestadística, este estudio permitió conseguir modelos teóricos de semivariogramas ajustados a los semivariogramas experimentales, con el valor de r2 y el cociente valor de la pepita sobre el valor de la meseta, lo cual verificó la calidad del ajuste. los semivariogramas experimentales encontrados, presentaron estructura de dependencia espacial, puesto que en ningún caso la semivarianza fue constante en función de la distancia, es decir, ninguno de los variogramas estimados exhibió una línea recta horizontal (giraldo 2002; Díaz - Francés 1993; isaaks y Srivastava 1989). Se ajustaron semivariogramas teóricos exponenciales y esféricos, acotados, exhibiendo una meseta u horizonte de correlación finito (Samper y Carrera 1990). Estos modelos son apropiados, para el análisis de una variable como la densidad de insectos en una zona de estudio, por ser los que mejor describen la distribución errática de la población en distancias pequeñas o reducidas (isaaks y Srivastava 1989).
En ninguno de los ajustes el valor de la pepita o nugget, superó el 50% de la meseta ó cota superior del variograma, una característica que según Díaz - Francés (1993), es recomendable para que el modelo de correlación describa bien la realidad. los mapas de distribución o de predicción permitieron identificar tendencias y agrupaciones de las variables. El número de obreras por cebo y el número de nidos/ m2 mostraron para los dos parches de bosque, un gradiente positivo desde el interior del bosque hacia el borde y un comportamiento agregado de W. auropunctata en todos los parches de bosques y sus matrices aledañas. En resumen, los resultados del modelamiento matemático en los fragmentos bosques fueron consistentes, es decir, el nivel de dependencia espacial indica una distribución agregada de la hormiga, y el comportamiento de la densidad y los nidos de obreras fue coherente respecto a un gradiente hacia los bordes, como lo exhiben los mapas de distribución de abundancia junto con las isolineas de nidos/m2, donde se muestra una relación directa entre las dos variables (Salguero 2008).
los mapas de distribución para obreras por cebo y nidos/m2, presentaron agregaciones con diferentes densidades poblacionales, considerablemente altas para la localidad de San Julián con un poco más de 1600 obreras por cebo, comparada con Alejandría, la cual fue aproximadamente de 141 obreras/cebo. Para ambos sitios los valores máximos de la variable coincidieron con los de agregaciones de nidos/m2. la distribución y densidad de la pequeña hormiga de fuego fue sorprendentemente contrastante en los dos bosques, pues estando en su rango nativo se esperaba que el comportamiento de la especie fuese multicolonial (le Breton et al. 2004), donde la agresividad intraespecífica fuese muy común, con tendencia a exhibir muchas colonias que presentasen límites territoriales definidos.
los resultados de este estudio pueden analizarse a la luz de los estudios de agresividad realizados por lópez (2005) en tres bosques secos de la cuenca media del río Cauca: San Julián, El medio y Alejandría. Esta investigadora encontró que cuando está en altas densidades poblacionales, la hormiga presenta muy bajos índices de agresividad y viceversa. Así, se justifica que la habilidad de W. auropunctata para alcanzar altas densidades poblacionales en San Julián se debe al hecho que presenta una baja agresividad intraespecífica en su rango nativo, fenómeno que también ocurre en los sitios donde la especie es introducida (Ulloa-Chacón y Cherix 1990).
No sucedió lo mismo con Alejandría, bosque donde presentó una baja densidad poblacional y que de acuerdo con lópez (2005), presentaría una alta agresividad intraespecífica en su rango nativo. Por lo anterior, es posible afrmar que, desde la interpolación exhibida en los mapas de distribución de obreras por cebo para San Julián, se concibe esta localidad como una gran unicolonia, para la cual la agresión intraespecífica no está (le Breton et al. 2004), lo que permite un libre intercambio entre los aproximadamente 15 nidos comprendidos en ella.
En Alejandría, el mapa de distribución de obreras por cebo exhibió tres agregaciones, con agrupaciones hasta de 7 nidos/m2, donde es posible que se presenten eventos de agresión intraespecífica y permite ratificar así su estructura social multicolonial, como se predice para su rango nativo. la riqueza de hormigas para este bosque alcanza las 69 especies (Armbrecht 1995; Armbrecht 1996; Armbrecht y Chacón de Ulloa 1997; lozano-zambrano 2002), por lo que se podría hipotetizar que la competencia contribuye a regular las poblaciones de la pequeña hormiga de fuego en esta localidad. Por otro lado, para San Julián, los parámetros del variograma para obreras por cebo, se estimó un rango Ao de 162,4 m (Tabla 1), el cual parece relativamente alto, teniendo en cuenta que la distancia máxima entre dos puntos extremos de la localidad uno ubicado en el potrero al norte y otro en el borde al sur no superaron los 310m, este resultado según giraldo 2002, es un indicador de fuerte dependencia espacial para el caso considerado. Dada la estructura social unicolonial para el bosque de San Julián discutida anteriormente, se puede establecer que para la variable obreras por cebo, las distancias hasta de 162,4m desde la esquina sureste, corresponderá a hormigas de la misma supercolonia.
Para Alejandría el valor del rango para obreras por cebo fue de 137,3m (Tabla 1), y la distancia entre las tres agregaciones fueron de 319,4m, 481,1m y 373,24m, mayores que el rango, lo que permitiría establecer que son colonias independientes, con lo que se estaría corroborando la naturaleza multicolonial discutida anteriormente para esta localidad.
Es posible que la perturbación y aislamiento de los bosques secos del valle geográfico del río Cauca, provoquen cambios demográficos en la pequeña hormiga de fuego y en el resto del ensamblaje de hormigas. Tres factores podrían explicar la dominancia de la pequeña hormiga de fuego: una de carácter genético, otra relacionada con su organización social y una tercera de carácter ecológico, relacionada con la perturbación humana mediante análisis con microsatélites en poblaciones nativas de W. auropunctata sugirieron que la dominancia de esta hormiga está relacionada con la perturbación humana (Foucaud et al. 2009). Foucaud y colegas (2009) mediante análisis con microsatélites en poblaciones nativas de W. auropunctata sugirieron que la dominancia de esta hormiga está relacionada con la perturbación humana. En el presente estudio, el parche de bosque San Julián ubicado al sur del área de estudio, ha sido uno de los más perturbados con una fragmentación severa, causada por la enorme intervención antrópica, y labores productivas más intensas (Arcila 2007). Además, este bosque es uno de los que mayor índice de aislamiento presentan (Arcila 2007).
El deterioro ambiental de los bosques se manifesta al comparar dos estudios separados por diez años. Armbrecht (1996), reportó una extensión de 6 hectáreas para San Julián, mientras que Arcila (2007) reportó 3,5 hectáreas para este mismo bosque. Además, la zona exhibe paisajes convertidos en guaduales y monocultivos de caña de azúcar, y hasta con penetración de ganado. Al tratarse de un bosque muy perturbado, hay un aumento de ramas y troncos caídos los cuales son aprovechados eficientemente por W. auropunctata para anidar (Armbrecht y Ulloa-Chacón 2003), lo que posiblemente contribuye a elevar su densidad de población. De lo anterior se puede concluir, que la connotación de W. auropunctata como especie vagabunda, que es atraída hacia hábitats perturbados (Fowler et al. 1990), es soportada por Armbrecht y Ulloa-Chacón (2003) en un estudio sobre el desempeño de la pequeña hormiga de fuego como bioindicadora de la diversidad de otras hormigas. las investigadoras encontraron que en catorce parches de bosque seco tropical en el valle geográfico del río Cauca (Colombia), los bosques más al sur (San Julián y otro llamado Colindres), fueron menos diversos y exhibieron las más altas abundancias de W. auropunctata, no tuvieron barreras protectoras en el borde del bosque y mostraron más signos de penetración humana y de ganado en su interior afectando la vegetación del sotobosque y calidad del suelo. los anteriores resultados fueron corroborados posteriormente para los mismos bosques por Achury et al. (2012).
Es digno de atención que precisamente aquellos bosques en los cuales W. auropunctata, y no especialmente otras especies de hormigas fue dominante, eran los menos diversos. Se sospecha que la tendencia de las poblaciones de esta hormiga a crecer en la dirección norte - sur en la zona de estudio, se debió precisamente a su preferencia por ambientes perturbados, con matrices manejadas intensamente en las cuales la transformación hacia monocultivos de azúcar está tomando lugar.
Conclusiones
Este estudio corrobora una vez más el valor de usar herramientas de la geoestadística para el modelamiento de fenómenos biológicos (rossi et al. 1992; Díaz y Casar 2004) tales como la densidad y distribución espacial de W. auropunctata. El modelo permitió revelar características en su rango nativo que explican por qué la especie es invasora en otros continentes o en islas. la obtención de una estructura de correlación espacial permite establecer si la escala y los métodos de muestreo utilizados fueron los apropiados en éste. Se logró evidenciar como en el parche de bosque de San Julián, que presenta la mayor intervención humana, la hormiga presenta una estructura social unicolonial, con altísima densidad (33 veces más que Alejandría) y tendencia a alcanzar los bordes. En contraste, la estructura social de W. auropunctata es multicolonial en el bosque de Alejandría, que es uno de los bosques más conservados del valle geográfico, y en este caso la hormiga se comporta espacialmente conforme con lo predicho para su rango nativo.
Agradecimientos A maría Dolly garcía y gladys Elena Salcedo por su invaluable asesoría académica. A los grupos de investigación gEA-DES de la Universidad Autónoma de occidente; Biología, Ecología y manejo de Hormigas de Universidad del Valle, gEAHNA de Universidad del Valle por sugerencias durante el estudio. Este estudio fue parcialmente financiado por la Universidad Autónoma de occidente y el instituto Alexander von Humboldt (obtención datos de campo).
Literatura citada ACHURY, R.; CHACÓN DE ULLO A, P.; ARCIL A, A. 2012. Effects of the heterogeneity of the landscape and the abundance of Wasmannia auropunctata on ground assemblages in a Tropical Dry Forest. Psyche. Doi:10.1155/2012/960475. [ Links ] ARCILA, A.M. 2007. Determinantes paisajísticos de la dinámica poblacional de la pequeña hormiga de fuego Wasmannia auropunctata (Roger) en fragmentos de bosque seco tropical. Tesis de Doctorado en Biología. Universidad del Valle, Cali, Colombia. 125 p. [ Links ] ARMBRECHT, I. 1995. Comparación de la mirmecofauna en fragmentos boscosos del Valle geográfico del río Cauca, Colombia. Boletín Museo de Entomología Universidad del Valle 3(2): 1-14. [ Links ] ARMBRECHT, I. 1996. Análisis de la diversidad del ensamblaje de hormigas en fragmentos de bosque seco en el Valle del Cauca, Colombia. Tesis de Maestría en Biología. Universidad del Valle. Cali. 130 p. [ Links ] ARMBRECHT, I.; ULLO A - CHACÓN, P. 2003. The Little Fire ant Wasmannia auropunctata (Roger) (Hymenoptera: Formicidae) as a diversity Indicator of Ants in Tropical Dry Forest Fragments of Colombia. Environmental Entomology 32 (3): 542-547. [ Links ] ARMBRECHT, I.; CHACÓN DE ULLO A, P. 1999. Rareza y diversidad de hormigas en fragmentos de bosque seco colombianos y sus matrices. Biotrópica 31: 646-653. [ Links ] ARMBRECHT, I.; CHACÓN DE ULLO A, P. 1997. Composición y diversidad de hormigas en bosques secos relictuales y sus alrededores, en el Valle del Cauca, Colombia. Revista Colombiana de Entomología 23: 45-50. [ Links ] BROWN, K. S. Jr. 1991. Conservation of neotropical environment, insects as indicators, pp. 305-404. En Conservation of insects and their habitats. Collins, N. M.; Thomas, J. A. (eds.). Academic Press. San Diego. [ Links ] BURBIDGE, A. H.; LEICESTER, K.; MCDAVITT, S.; MAJER, J. D. 1992. Ants as indicators of disturbance at Yanchep National Park, Western Australia. Journal of the Royal Society of Western Australia 75: 89-95. [ Links ] CAMBARDELL A, C.; MOORM AN, T.; NOVAK, J.; PARKIN, T.; KARL EN, D.; TURCO, R.; KONOPKA, A. 1994. Field scale variability of soil properties in central Iowa soils. Soil Science Society of American Journal 58: 1501-1511. [ Links ] CARROL , S. S.; PEARSON, L. 2000. Detecting and modeling spatial and temporal dependence in conservation biology. Conservation Biology 14 (6): 1893-1897. [ Links ] CHACÓN DE ULLO A, P.; ARM BRECHT, I. 2006. Las hormigas del ecosistema bosque seco tropical. Tomo II , pp. 345-351. En Chávez, M. E.; Santamaría, M. (eds.). Informe Nacional sobre el Avance en el Conocimiento y la Información de la Biodiversidad 1998-2004. Instituto de Investigaciones de Recursos Biológicos Alexander von Humboldt. Bogotá D.C., Colombia. 2 Tomos. [ Links ] CLARK, D. B.; GUAYASAMÍ N, C.; PAZMI NO, O.; PAEZ DE VILL ACÍS, Y. 1982: The tramp ant Wasmannia auropunctata: autoecology and effects on ant diversity and distribution on Santa Cruz Island, Galápagos. Biotrópica 14: 196-207. [ Links ] CVC. Corporación Autónoma Regional del Valle del Cauca. 1990. Comparación de la cobertura de bosques y humedales entre 1957 y 1986 con delimitación de las comunidades críticas del valle geográfico del río Cauca. Informe. Cali. 49 pp. [ Links ] DÍAZ-FRANCÉS, E. 1993. Introducción a Conceptos Básicos de geoestadística. Memorias seminario Estadística y Medio Ambiente. Centro de Investigación en Matemáticas, CIM AT, Guanajuato. México. [ Links ] DIAZ-VIERA, M. A.; CASAR-GONZÁLEZ, R. 2004. Geoestadística aplicada. Tema: Geoestadística fractal. [online] [consultado 2 julio, 2011] Available from World Wide Web: (URL : http://132.248.182.189/cursos/geoest/Presentaciones/CG8a. pdf). [ Links ] DIDHAM, R. K.; HAMMO ND, P. M.; LAWTON, J. H.; EGGLETON, P.; STOR K, N. E. 1998. Beetle species responses to tropical forest fragmentation. Ecological Monographs 68: 295- 303. [ Links ] DIDIER, G.; LOZANO, F.; OSPINA, J. L; QUICENO, M. P. 2001. Plan de acción regional en biodiversidad para norte de Santander. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. Cúcuta, 200 p. [ Links ] ESRI ® ARCGI S TM 9.0 GI S and mapping software. [ Links ] FOUCAUD, J.; ORIVEL, J.; FOURNIER, D.; DELABIE, J. H. C.; LOISEAU, A.; LEBRETON, J.; CERDAN, P.; ESTOUP, A. 2009. Reproductive system, social organization, human disturbance and ecological dominance in native populations of the little fire ant, Wasmannia auropunctata. Molecular Ecology 18 (24): 5059-5073. [ Links ] FOWLER H. G.; BERNARDI, J.; DELABIE, J.; FOR TI, L.; PEREIR A DA SIL VA, V. 1990. Major ant problems in South America. pp. 3-14. In Applied myrmecology: A world perspective. Vander Meer, R.K.; Jaffe, J.; Cedeño, A., (eds). Westview press. Inc Boulder, Colorado. [ Links ] GIRALDO, H. R. 2002. Introducción a la Geoestadística. Teoría y aplicación. Universidad Nacional de Colombia. Simposio de Estadística. 250 p. [ Links ] GS+ Geostatistics for the Environmental Sciences Version 5.1. Gamma Design Software 1982- 2002 Stanford Center for reservoir Forecasting. [ Links ] HÖLL DOBLER, B.; WIL SON, E. O. 1990. The ants. Springer - Verlag, Berlin. 450 p. [ Links ] HURTADO, H.; SALCEDO, G. E.; SERR ANO, A. J. 1996. Modelos Estadísticos para información autocorrelacionada. Cuadernos de Biomatemáticas 2. Universidad de Quindío. 60 p. [ Links ] HURTADO, L. H.; SALCEDO, G. E.; GARCÍA, M. D. 2007. Design and validation of an optimal network for monitoring the Water quality in a hydrological system: A case study. Proceedings. of the 2006 International of Mathematical and Computational Biology. Biomat 2006 322-336. E - paper serviços editorial is Ltda. Rio de Janeiro - Brasil. [ Links ] IAvH, 1998. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. Informe Nacional sobre el estado de la biodiversidad 1997 - Colombia. Bosque seco tropical, pp. 56-71. En Chávez, M.E.; Arango, N. (eds.). Tomo I: Diversidad Biológica. Instituto Humboldt, PNUMA, Ministerio del Medio Ambiente, vol. 3. Santafé de Bogotá, Colombia. [ Links ] ISAAKS, E. H.; SRI VASTAVA, R. M. 1989. An introduction to applied geostatistics. Oxford University Press, New York. [ Links ] KATTAN, G. H. 2002. Fragmentación: patrones y mecanismos de extinción de especies, pp. 561-590. En Guariguata, M.R.; Kattan, G.H. (eds.). Ecología y conservación de bosques. Libro Universitario Regional. Editorial Tecnológica de Costa Rica. [ Links ] KOTZE, D.; SAMWAYS, M. J. 2001. No general edges effects for invertebrates at Afromontane forest / grassland ecotones. Biodiversity and Conservation 10: 443-446. [ Links ] KREMEN, C.; COLWELL , R. K.; ERWIN, T. L.; MURPHY, D. D.; NOSS, R. F.; SANJAYAN, M. A. 1993. Terrestrial arthropod assemblages: their use in conservation planning. Conservation Biology 7: 796-808. [ Links ] LAURANCE, W. F.; VASCONCELO S, H. L. 2001. Ecological Effects of Habitat Fragmentation in the Tropics, pp. 33 -49. En Schroth G.; Da Fonseca, G. A. B.; Harvey, C. A.; Gascon, C.; Vasconcelos, H. L.; Izac, A. M. N. (eds.). Agroforestry and Biodiversity Conservation in Tropical. Island Press. [ Links ] LE BRETON, J.; DELABIE, J. H. C.; CHAZEAU, J.; DE JEAN, A.; JOURDAN, H. 2004. Experimental evidence of Large - scale unicoloniality in the tramp ant Wasmannia auropunctata (Roger). Journal of Insect Behavior. 17 (2): 263-271. [ Links ] LIEBHOL D, A. M.; RO SSI, R. E.; KEMP, W. P. 1993. Geostatistics and geographic information systems in applied insect ecology. Annual Review of Entomology 38: 303-327. [ Links ] LÓPEZ, M. P. 2005. Colonias de Wasmannia auropunctata (Roger) (Hymenoptera: Formicidae) Composición y agresividad en biotopos con diferente grado de perturbación en el Valle del Cauca. Trabajo de grado. Programa de Biología. Facultad de Ciencias. Universidad del Valle. Cali, Colombia. 55 pp. [ Links ] LOZANO-ZAMBRANO, F. H. 2002. Hormigas, diversidad y relaciones especies/área en fragmentos de bosque seco Tropical- Tesis de Maestría. Programa de postgrado en Ciencias - Biología. Facultad de Ciencias. Universidad del Valle, Cali - Colombia. [ Links ] MORAL, F. J. 2004. Aplicaciones de la Geoestadística en las Ciencias Ambientales. [online] [consultado 2 julio, 2006] Ecosistemas 2004/1 Available from World Wide Web: (URL : http//www.aet.org/ecosistemas/041/revision3.htm). [ Links ] NORDMEYER, H. 2009. Spatial and temporal dynamics of Apera spica-venti seedling populations. Crop Protection. 28 (10): 831- 837. [ Links ] PASSERA, L. 1994. Characteristics of tramp species, pp. 23-41. En Williams, D.F. (eds.). Exotic Ants Biology, impact and control of Introduced species. Westview Studies in Insect Biology. Boulder. Colorado. [ Links ] Philpott , S. M., I. Perfect o, I.; Armbrecht , I.; C. L. 2010. Effects of disturbance and habitat transformation on ant diversity and function. Chapter 8. Pag. 137-156 En: L. Lach, C. Parr & K, Editors. Abbott Ant Ecology. Oxford University Press. Oxford, UK. [ Links ] RAMÍREZ - DÁVIL A, J. F.; POR CAYO-CAMARGO , E. 2008. Distribución espacial de las ninfas de Jacobiasca lybica (Hemíptera: Cicadellidae) en un viñedo de Andalucía, España. Revista Colombiana de Entomología 34 (2): 169-175. [ Links ] RAMÍREZ - DÁVIL A, J. F.; GONZÁLEZ - ANDUJAR, J. L.; LÓPEZ - MARTÍNEZ, M. A.; OCETE, R. 2005. Modelización y mapeo de la distribución espacial de las ninfas del mosquito verde Jacobiasca lybica (Bergenin & Zanon) (Hemíptera: Cicadellidae) en viñedo modelización y mapeo. Boletín Sanidad Vegetal Plagas 31: 119-132. [ Links ] ROBERTSON, G. P. 1987. Geostatistics in Ecology: Interpolating with known variance. Ecology 68 (3): 744-748. [ Links ] ROSSI, R. E.; MULLA, D. J.; JOURNEL, A. G.; FRANZ, E. H. 1992. Geostatistical tools for modelling and interpreting ecological spatial dependence. Ecological Monographs, 62 (2) pp. 277-314. [ Links ] SALGUERO , B. 2008. "Modelación matemática de la distribución espacial de Wasmannia auropunctata (Roger) y su relación con el hábitat en parches de bosque seco tropical en el valle geográfico del río Cauca" Tesis de Maestría. Universidad del Valle, Facultad de Ciencias, Departamento de Biología. 235 p. [ Links ] SAMPER, F. J.; CARRERA, J. 1990.Geoestadística. Aplicaciones a la Hidrogeología Subterránea. Centro Internacional de Métodos Numéricos en Ingeniería. Universidad Politécnica de Catalunya. Barcelona. 250 p. [ Links ] SILBERGLI ED, R. 1972. The little fire ant, Wasmannia auropunctata serious pest in the Galapagos Island. Noticias de Galápagos 19/20: 13-15. [ Links ] STATGR APHICS 5 PLUS 2000. A manugistic product. Statistical Graphics Corporation. [ Links ] TAYLOR , L. R. 1984. Assessing and interpreting the spatial distributions of insect populations - Annual Review of Entomology 29: 321: 357. [ Links ] ULLOA - CHACÓN, P. 1990. Biologie de la reproduction chez la petite fourmi de feu Wasmannia auropunctata (Roger) (Hymneoptera: Formicidae). Tesis de doctorado. University de Lausanne. 161 p. [ Links ] ULLOA-CHACÓN, P.; CHERI X, D. 1990. The little Fire Ant Wasmannia auropunctata (Roger) (Hymenoptera: Formicidae), pp. 281-289 In: Vander Meer, R. K.; Jaffe, K.; Cedeño, A. (eds.). Applied Myrmecology: a world perspective. Westview Press, Boulder, Colorado. [ Links ] VELASCO, L. M. 1982. Historia del hábitat vallecaucano 1536- 1982. Corporación Autónoma Regional del Valle del Cauca, CVC, Cali Colombia. 105 pp. [ Links ] WETTERER, J. K.; POR TER, S. D. 2003. The little fire ant, Wasmannia auropunctata (Roger): distribution, impact and control. Sociobiology 42: 1-41. [ Links ] WETTERER, J. K.; WALSH, P. D.; WHITE, L. J. T. 1999. Wasmannia auropunctata (Roger) (Hymenoptera: Formicidae), a highly destructive tramp ant, in wildlife refuges of Gabon. West Africa. African Entomology 7: 292-294. [ Links ] ZAS, R. 2006. Consecuencias de la estructura especial en el diseño y análisis de experimentos en campo. [online] [citado 2 julio, 2006] Ecosistemas 15 (3): 107-114. Available from World Wide Web: (URL : http: //www.revistaecosistemas.net/articulo.asp?Id=434). [ Links ]


{kind=link}